

1 32 Глава 5
клетки с ориентацией 30°, все клетки с ориентацией 42° и т. д.; и наконец, объединить все клетки с доминированием левого глаза и все — с доминированием правого глаза. Если бы мы знали, как устроена кора, то смогли бы предложить много ответов на эти вопросы. Сейчас выскажем одно предположение: возможно, что клетки с разной ориентацией действительно оказывают тормозящее воздействие друг на друга. Если нежелательно, чтобы какая-то клетка отвечала на другие ориентации, отличные от ее собственной, то нетрудно представить себе наличие тормозных связей, которые обостряют настройку данной клетки на определенную ориентацию. Тогда наблюдаемая картина — как раз то, что нам нужно: клетки с одинаковой ориентацией расположены ближе всего друг к другу, но не слишком далеко от клеток с иной, но очень сходной ориентацией; тогда тормозные связи будут достаточно короткими. Возможно другое предположение: если рассмотреть, какие входы от клеток слоя 4 (с круглыми полями) нужны для организации простой клетки с определенной оптимальной ориентацией, легко увидеть, что потребуется лишь небольшое изменение набора входов для того, чтобы получить близко расположенную простую клетку с близкой, но несколько иной ориентацией. Для этого достаточно добавить несколько новых входов, а несколько прежних убрать (рис. 89). Такого рода соображения позволили бы объяснить физическую близость расположения клеток со сходной ориентацией.
Предметом следующей главы будет взаимосвязь между ориентацией, глазо-доминантностью и проекцией поля зрения на кору. Рассмотрение этого вопроса может помочь нам понять, почему нужно так много колонок. Когда к этому добавится еще вопрос о топографии, соответствующая корковая организация будет представляться еще более сложной и интересной.
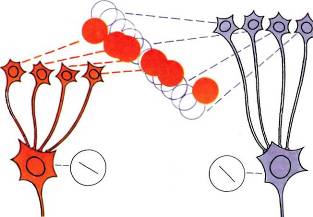
Рис. 89. Показана группа клеток слоя 4 с концентрическими рецептивными полями, необходимая для создания простой клетки, отвечающей на наклонную линию с ориентацией «4.00 — 10.00». Вероятно, эта группа имеет общие клетки с группой, необходимой для реакции на линию с ориентацией «4.30 —10.30», — для превращения одной группы в другую достаточно лишь несколько связей убрать, а несколько добавить.
6. «УВЕЛИЧЕНИЕ» И МОДУЛИ
В предыдущей главе я подчеркивал, что первичная зрительная кора кажется морфологически однородной как невооруженному глазу, так и под микроскопом при использовании большинства обычных методов окрашивания. Однако при более тщательном изучении выяснилось, что однородна и топография колонок глазодоминантности: период чередования зон доминирования левого и правого глаза остается на удивление постоянным от проекции центральной ямки (точки фиксации) до дальней периферии бинокулярной части поля зрения. При помощи метода с инъекцией дезоксиглюкозы мы выявили также однородность топографии ориентационных колонок.
Факт анатомической однородности коры мы вначале восприняли как неожиданность, так как с функциональной точки зрения эта область явно неоднородна по двум важным параметрам. Во-первых, как уже говорилось в главе 3, рецептивные поля ганглиозных клеток сетчатки в зоне центральной ямки и около нее гораздо меньше, чем на периферии. Размеры рецептивного поля типичной сложной клетки в верхнем слое коры в области проекции центральной ямки составляют примерно 0,25x0,5°. Если же рассматривать те части коры, где представлена периферия сетчатки (80—90° от центральной ямки), то здесь чаще всего встречаются рецептивные поля размером 2x4°. Если сравнивать соответствующие площади, то отношение между ними можно оценить как 1:10 или даже 1:30.
Второй вид неоднородности связан с так называемым «увеличением». Это понятие, введенное в 1961 году П. Дэниелом и Д. Уиттериджем, означает расстояние между двумя точками коры, в которые проецируются две точки поля зрения, разделенные расстоянием в 1°. Если идти от центральной ямки к периферии, корковая проекция одного и того же углового расстояния будет становиться все меньше и меньше, т. е. «увеличение» будет уменьшаться. Если сместить стимул на 1° около точки фиксации, то его проекция в коре сдвинется примерно на 6 мм. Если же поместить этот стимул в зону, отстоящую от точки фиксации на 90°, то сдвиг в поле зрения на 1° будет соответствовать сдвигу вдоль поверхности коры всего лишь на 0,15 мм. Таким образом, «увеличение» в центре сетчатки примерно в 36 раз больше, чем на периферии.
Обе рассмотренные неоднородности связаны с одним и тем же обстоятельством — с уменьшением остроты зрения по мере удаления от центра сетчатки к ее периферии. Попробуйте, например, фиксировать взором какую-нибудь букву в самом начале строки и попытайтесь определить, какая буква или слово находится в самом ее конце. Или фиксируйте взглядом букву п в начале слова прогрессивно. Вряд ли вам удастся опознать букву о в конце слова. Вероятно, вам трудно будет опознать также вин перед последней буквой. Для того чтобы