

128 Глава 5
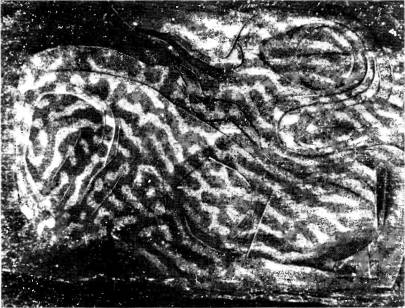
Рис. 86. Тому же животному, что и в предыдущем случае, в глаз неделей раньше была введена радиоактивная аминокислота (пролин). После промывания среза в воде с целью растворить 2-дезоксиглюкозу был получен радиоавтограф. Радиоактивная метка выявляет здесь колонки глазодоминантности. Эти колонки не имеют никакой видимой связи с ориентационными колонками.
промежуточные варианты снова дойти до вертикальной ориентации). В некоторых местах мы находили структуру в виде правильных полос на площади в несколько квадратных миллиметров. Мы задавались вопросом: нет ли какой-то связи между конфигурациями ориентационных полос и полос глазо доминантности — может быть, например, они идут параллельно или же пересекаются под прямым углом? В том же самом эксперименте нам удалось выявить глазодоминантные колонки, после того как в глаз животного была введена радиоактивная аминокислота, а затем полученные срезы ткани анализировались с помощью двух методик (результат показан на рис. 86). Мы не смогли обнаружить никакой явной корреляции. Сравнивая сложную картину ориентационных колонок с гораздо более простым распределением колонок глазодо-минантности, очень трудно было решить, какая могла бы быть связь между ними. .
У метода с применением дезоксиглюкозы есть один серьезный недостаток: никогда нет полной уверенности в том, что полученная картина распределения действительно связана с интересующим нас свойством стимула. Например, когда в качестве стимула используется рисунок из черных и белых вертикальных полос, как можно убедиться, что получившееся распределение связано именно с ориентацией этих полос, т. е. что в темных областях находятся клетки, отвечающие на вертикальные линии, а в светлых — клетки, которые на них не реагируют? Ведь вместо этого решающей особенностью данного стимула могло бы быть то, что он черно-белый, а не цветной, или же то, что он состоит из широких, а не узких полос, или, наконец, то, что экран расположен именно на таком, а не ином расстоянии от животного. Одно из косвенных подтвержде-
АРХИТЕКТУРА ЗРИТЕЛЬНОЙ КОРЫ 129
нии того, что действие дезоксиглюкозы затрагивает механизмы настройки на ориентацию, — это отсутствие каких-либо пятен или полос в слое 4С, где клетки не обладают избирательностью к ориентации. Другим подтверждением служит результат, полученный в исследованиях М. Страйкера из Калифорнийского университета в Сан-Франциско. Страйкер делал протяженные проходки микроэлектродом в стриарнои коре кошек параллельно ее поверхности и каждый раз, когда встречались клетки с определенной оптимальной ориентацией, производил локальное разрушение ткани; затем после инъекции радиоактивной дезоксиглюкозы включался стимул с полосками одинаковой ориентации. В этих экспериментах была обнаружена явная корреляция между получаемым распределением метки на гистологическом срезе и ориентацией стимула. Наиболее четко удалось недавно выявить ориентационные колонки с помощью красителей, чувствительных к электрическому напряжению, которые в течение многих лет разрабатывал Л. Коэн в Йеле. Г. Блэсдел из Питт-сбургского университета применил их при исследовании коры головного мозга. При использовании этого метода краситель, чувствительный к напряжению и
окрашивающий клеточные мембраны, наносят на поверхность коры наркотизированного животного, и нервные клетки поглощают его. Когда в эксперименте животному предъявляют стимул, все реагирующие на него клетки изменяют свой цвет. Если в области, близкой к поверхности коры, таких клеток окажется достаточно много, эти изменения цвета можно уловить с помощью
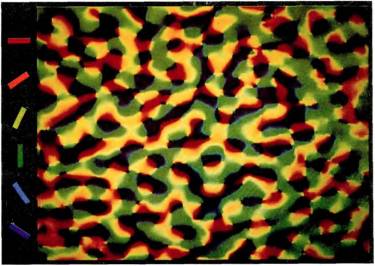
Рис. 87. В этом эксперименте Г. Блэсдел использовал для обработки стриарнои коры обезьяны окрашивание, чувствительное к электрическому напряжению. Обезьяне предъявлялись полосы последовательно изменяющейся ориентации. Изображение коры было получено с помощью телевизионной техники. С помощью компьютерной обработки изображений было получено окончательное изображение, на котором участкам, реагирующим на ту или иную ориентацию стимула, был приписан определенный цвет. В любой небольшой области коры ориентационные зоны проявляются в виде параллельных полос, так что в целом полный набор элементов, настроенных на разные ориентации, дает картину, окрашенную во все цвета радуги.
130 ГЛАВА 5
современных методов обработки телевизионных изображений. Хотя изменения цвета едва заметны, с помощью чувствительных телекамер и машинных методов фильтрации шумов их можно обнаружить. Блэсдел использовал в своих опытах стимуляцию полосами определенной ориентации, после чего фотографировал полученное распределение активности клеток на участке площадью в несколько квадратных сантиметров. Затем та же процедура повторялась при многих других ориентациях стимула. После этого каждой ориентации был поставлен в соответствие свой цвет — красный для вертикальной, оранжевый для положения часовой стрелки в 1 час и т. д.; потом все изображения были наложены друг на друга. Поскольку изоориентационная линия должна постепенно смещаться по мере изменения ориентации, в любой небольшой зоне изображения должен возникнуть радужный узор. Именно это и обнаружил Блэсдел в действительности. Примеров успешного применения этого метода пока еще мало, и нет возможности интерпретировать получаемые изображения в таких понятиях, как разрывы и реверсии, однако метод представляется многообещающим.
Карты коры
Теперь, когда мы узнали кое-что о топографическом представлении в коре параметров ориентации и глазодоминантности, мы можем рассмотреть взаимосвязь между этими отображениями и проекциями зрительных полей. Обычно говорят, что сетчатка отображается на кору взаимно однозначно. Однако, учитывая то, что нам известно о рецептивных полях корковых клеток, можно с уверенностью сказать, что это утверждение, строго говоря, неверно, поскольку каждая клетка получает входные сигналы от тысяч палочек и колбочек, а ее рецептивное поле — далеко не точка. Отображение сетчатки на кору гораздо более сложно, чем любое обычное взаимно однозначное отобра-
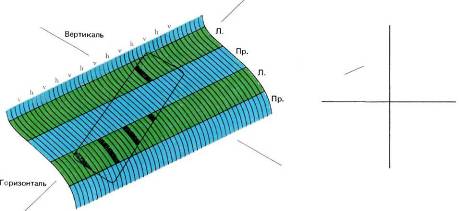
Рис. 88. Наклонный отрезок прямой, предъявленный левому глазу (справа), мог бы вызывать распределение возбуждения в небольшой зоне стриарной коры, показанное слева. Возбужденные участки будут ограничены небольшой зоной коры — длинной и узкой в соответствии с формой стимула. В пределах самой этой зоны возбуждение ограничено колонками глазодоминантности для левого глаза и только теми ориентационными колонками, которые соответствуют ориентации часовой стрелки в 2.00 или 8.00. Как видно, корковое отображение данного стимула не так просто!
АРХИТЕКТУРА ЗРИТЕЛЬНОЙ КОРЫ 131
жение. На рис. 88 я попытался показать распределение в коре тех областей, которые активируются под действием простого стимула (не следует смешивать это с рецептивным полем одиночного нейрона). Стимул здесь — это короткий отрезок линии с наклоном в 60°относительно вертикали, предъявленный только левому глазу. Мы предполагаем, что этот элемент поля зрения проецируется на область коры, выделенную прямоугольником с закругленными углами. В пределах этой области будут возбуждены только те полосы, которые связаны с левым глазом, а в этих полосах — только те участки, которые соответствуют ориентации в 60°. Все они на рисунке зачернены. Таким образом, линия, появившаяся в поле зрения, вызывает причудливую картину распределения возбужденных клеток коры в виде ряда узких полосок.
Теперь становится понятно, насколько наивно представление о зеленом гомункулюсе, который сидит у нас в голове и рассматривает передаваемую в кору картину. На самом деле картина, которая создается в коре, имеет примерно такое же отношение к изображению внешнего мира, как электрические процессы в телекамеры — к передаваемой внешней сцене. Если бы распределение корковой активности точно воспроизводило распределение яркости в поле зрения, то это означало бы, что на пути от сетчатки в кору не произошло ничего существенного. В этом случае действительно потребовался бы маленький зеленый человечек.
Вряд ли можно представить себе, что в процессе эволюции было затрачено столько усилий на столь изящную группировку клеток в двух взаимонезависимых системах колонок, если бы это не обеспечивало какой-то пользы для животного. Пока мы не докопаемся до точной схемы связей, определяющей отображение сетчатки в коре, мы вряд ли сможем до конца понять принципы группировки клеток в коре. Сейчас мы можем только строить логичные догадки. Если мы предположим, что схема, приведенная в главе 4, соответствует реальности, то придется признать: то, что нужно для перехода от простых клеток к сложным или для создания чувствительности к концам линий или к направлению движения, — это определенного рода конвергенция множества клеток на одну клетку, причем у рецептивных полей промежуточных клеток должна быть одна и та же ориентация и примерно одно и то же положение. Пока у нас не было особых оснований предполагать, что клетка с определенной ориентацией рецептивного поля получает входы от клеток с рецептивными полями иной ориентации. (Я здесь несколько преувеличиваю: высказывались предположения, что клетки с разной оптимальной ориентацией стимулов могут быть соединены тормозными связями; в пользу этого имеются лишь косвенные данные, на мой взгляд, не очень убедительные, хотя их нельзя игнорировать.) Если это так, то почему бы не группировать вместе клетки, которые должны быть взаимно связаны? Альтернативный вариант едва ли целесообразен — легко представить себе, как трудно было бы обеспечить надлежащие связи между клетками, если эти клетки разбросаны в коре без учета их сходных свойств! Особенно много связей должно быть между клетками с одинаковой оптимальной ориентацией, и если бы такие клетки распределялись случайным образом, то сеть аксонов, необходимых для их соединения, заполнила бы все пространство между нейронами. Как бы то ни было, фактически они группируются вместе. Изложенные соображения относятся и к клеткам, сходным по глазодоминантности.
Но если клетки со сходными свойствами упаковываются вместе, то почему, продвигая электрод, мы наблюдаем последовательное изменение ориентации небольшими шагами? И чем объясняется цикличность — почему, пройдя все возможные варианты, мы опять возвращаемся к исходной ориентации, а потом весь цикл повторяется снова? Казалось бы, можно было собрать вместе все