

Глава 5
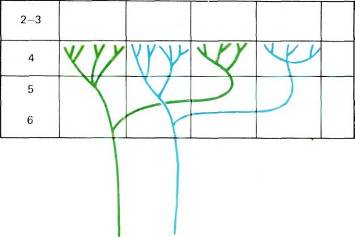
Рис. 69. Каждый аксон из HKT сначала проходит в стриарной коре через нижние слои, последовательно ветвится и затем оканчивается в слое 4С. Здесь он образует скопления синаптических окончаний шириной 0,5 мм, отделенные друг от друга промежутками такой же ширины. Все волокна от одного глаза группируются в одних и тех же участках. В промежутках между этими участками группируются волокна от другого глаза. Область разветвлений одного волокна из крупноклеточного слоя HKT может составлять от 2 до 3 мм (окончания находятся в слое 4Са). Волокно из мелкоклеточного слоя HKT ветвится в более ограниченной области (в слое 4Ср), обычно занимающей лишь одну или две колонки.
настоящее время чаще всего используют фермент пероксидазу из хрена. Этот фермент распространяется по аксону в обоих направлениях и катализирует химическую реакцию, продукт которой очень эффективно окрашивает клетку. Поскольку фермент служит катализатором, достаточно ничтожных его количеств, чтобы получить интенсивную окраску. Важно и то, что в самой нервной ткани фермента с подобными свойствами нет: это исключает возможность нежелательной окраски фона.
Тот факт, что кора разбита на колонки глазодоминантности, проходящие от поверхности мозга вплоть до белого вещества, подтвердил морфологические данные о том, что группы клеток в слое 4С служат главными местами передачи зрительной информации в клеточные слои, лежащие выше и ниже этого слоя. Наличие некоторого числа горизонтальных и диагональных связей длиной около миллиметра, идущих по всем направлениям, должно приводить к некоторому размыванию четких зон доминирования правого или левого глаза в слоях, расположенных выше и ниже слоя 4С (рис. 70). Поэтому можно ожидать, что клетка, расположенная прямо над центром зоны доминирования левого глаза в слое 4, будет явно лучше отвечать на стимуляцию этого глаза (а возможно, и полностью контролироваться им), тогда как клетка, расположенная ближе к границе между участками разной глазодоминантности, может оказаться бинокулярной без какого-либо доминирования одного из глаз. Действительно, при горизонтальном передвижении электрода в верхнем слое коры или же в слое 5 или 6 отмечаются постепенные изменения глазодоминантности: сначала будут встречаться клетки, намного сильнее активируемые одним глазом, затем клетки, у которых эта асимметрия выражена слабее, потом клетки с равной зависимостью от обоих глаз, после чего начнет усиливаться влияние другого «глаза. Такие плавные переходы резко контрастируют с той внезапной сменой глазодоминантности, которую мы наблюдаем при движении электрода вдоль слоя 4С (рис. 71).
АРХИТЕКТУРА ЗРИТЕЛЬНОЙ КОРЫ 115
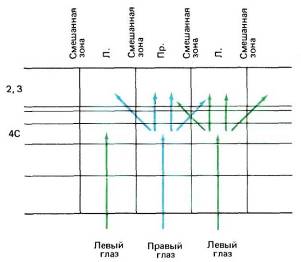
Рис. 70. Из-за наличия горизонтальных и диагональных связей выше слоя 4 происходит перекрывание колонок глазодоминантности и границы их размываются.
Если смотреть «сбоку», две совокупности клеток в слое 4С выглядят как чередующиеся участки. Однако нам хотелось узнать, как будет выглядеть распределение этих участков при взгляде на поверхность коры сверху. Допустим, что они отмечены на поверхности коры черным или белым цветом. С точки зрения топологии можно распределить эти участки несколькими разными способами: в шахматном порядке, в виде чередующихся черных и белых полос, в виде черных пятен на белом фоне или использовать любые комбинации этих паттернов. На рис. 72 показаны три возможных варианта такого распределения. Решать поставленную задачу с помощью одних только микроэлектродов означает использовать одномерные средства для расшифровки неизвестной трехмерной структуры. Это задача безнадежная — все равно что пытаться
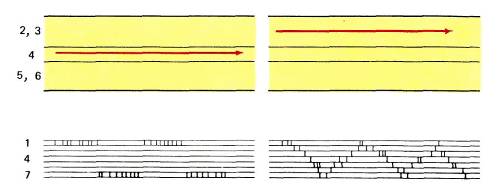
Рис. 71. В отличие от четких границ между колонками глазодоминантности в слое 4 коры, границы колонок в верхних (2 и 3) и нижних (5 и 6) слоях коры размыты. Стрелками показано направление продвижения микроэлектрода в слое 4 (вверху слева) и в слое 2 или 3 (вверху справа). Внизу представлена глазодоминантность клеток, ответы которых регистрировались в одной проходке микроэлектрода. В слое 4 видны резкие переходы от группы 1 (влияет только контралатеральный глаз) к группе 7 (влияет только ипсилатеральный глаз). В других слоях встречается много бинокулярных клеток, а глазодоминантность периодически изменяется, проходя через промежуточные уровни (цифры 1, 4 и 7 означают категории клеток по глазодоминантности).