

110 Глава 5
клеткой стимул, мы почти всегда использовали внеклеточное отведение, помещая кончик микроэлектрода рядом с клеткой; в этом случае регистрировались не изменения мембранного потенциала, а токи, связанные с импульсами. Часто при этом оказывалось, что мы регистрируем реакции не одной, а нескольких клеток одновременно — скажем, в том случае, когда кончик микроэлектрода останавливался на полпути между двумя телами нейронов. Импульсные разряды одиночных клеток при таком отведении почти идентичны, однако величина и форма импульсов зависят от расстояния и от взаимного расположения клеток, так что разряды, отводимые одновременно от двух клеток, обычно оказываются разными, и поэтомуих можно легко различить. Выполняя такого рода отведения от двух клеток, мы смогли отчетливо увидеть, чем различаются соседние клетки и в чем они одинаковы.
В одном из первых таких отведений были обнаружены две корковые клетки, отвечавшие на противоположные движения руки, которой махали перед животным. В этом случае две лежавшие бок о бок клетки давали разные (по существу, противоположные) реакции на движение стимула. Однако в других отношениях эти клетки почти наверное проявляли сходные свойства. Если бы я в 1956 году был уже подготовлен к изучению ориентационной избирательности, то я, весьма вероятно, обнаружил бы, что обе предпочитаемые ориентации близки к вертикальной, так как данные клетки хорошо отвечали на горизонтальные движения стимула. Тот факт, что обе они реагировали на возвратно-поступательное движение руки, означает, что положения рецептивных полей этих клеток приблизительно совпадали. Если бы я исследовал эти нейроны на глазодоминантность, то скорее всего этот параметр тоже оказался бы одинаковым.
Уже в первых записях активности корковых нейронов нас поразило то, как часто две клетки, реакции которых можно регистрировать одновременно, одинаковы по глазодоминантности, сложности и, что самое удивительное, по оптимальной ориентации стимулов. Такие совпадения, вряд ли случайные, позволяют предположить, что клетки с одними и теми же свойствами объединены в группы. Возможность такой группировки весьма заинтересовала нас, и как только это предположение подтвердилось, мы стали выяснять, каковы размеры и форма этих групп.
Исследование коры
С помощью микроэлектродов можно исследовать только отдельные точки коры. Для того чтобы получить представление о трехмерной организации мозга, приходится медленно погружать электрод в глубину, время от времени останавливать его для записи активности какой-нибудь клетки (а возможно, — двух или трех клеток), отмечать по специальной шкале показания глубины, а затем повторять все сначала. Рано или поздно кончик микроэлектрода пройдет через весь корковый слой, и тогда электрод можно вынуть и снова ввести его в каком-нибудь другом месте. После окончания эксперимента делают срез, окрашивают его и исследуют под микроскопом с целью определить положение каждой из нервных клеток, активность которых регистрировалась. В одном эксперименте длительностью около 24 часов обычно удается сделать две-три проходки примерно по 4—5 миллиметров каждая. За одну проходку можно наблюдать ответы примерно 200 клеток.
Микроэлектрод настолько тонок, что едва удается найти след от его прохождения под микроскопом, поэтому нет оснований думать, что при проходке микроэлектродом будет повреждено много клеток и это может повлиять на
АРХИТЕКТУРА ЗРИТЕЛЬНОЙ КОРЫ 111
реакции близлежащих нейронов. Вначале нам было трудно находить на гистологическом срезе след от микроэлектрода, не говоря уже о том, чтобы определить положение кончика электрода, и это затрудняло оценку положения тех клеток, активность которых была записана. Эту трудность удалось преодолеть, когда было обнаружено, что пропускание через микроэлектрод слабого тока приводит к разрушению клеток в ближайшей окрестности кончика микроэлектрода, и на гистологических срезах эта зона разрушения хорошо видна. К счастью, при пропускании тока сам микроэлектрод не повреждается. Поэтому за одну проходку ток пропускают 3—4 раза, отмечая при этом глубину погружения микроэлектрода, а так как глубину отмечают и при регистрации активности клеток, то можно оценить и положение каждой из них. Разумеется, при воздействии тока погибает несколько клеток около кончика микроэлектрода, однако их не так много, чтобы могла быть нарушена работа более удаленных нейронов. Для того чтобы при этом не исказить ответы клеток, лежащих впереди на пути микроэлектрода, его кончик немного продвигают вперед, регистрируют активность клеток, а затем кончик отводят назад и тогда уже пропускают ток.
Вариации в сложности
Как и следовало ожидать, клетки во входном корковом слое — слое 4 — проявляют более простое поведение, чем клетки на выходе. Мы уже отмечали в этой главе, что у обезьяны клетки в слое 4С(3, куда приходят волокна из четырех верхних (мелкоклеточных) слоев НКТ, по-видимому, не обладают избирательностью к ориентации стимула и ведут себя подобно клеткам, имеющим круглые рецептивные поля с центром и периферией. В слое 4Са, имеющем входы от двух вентральных (крупноклеточных) слоев НКТ, некоторые клетки обладают круглыми полями с центром и периферией, а всем остальным, видимо,
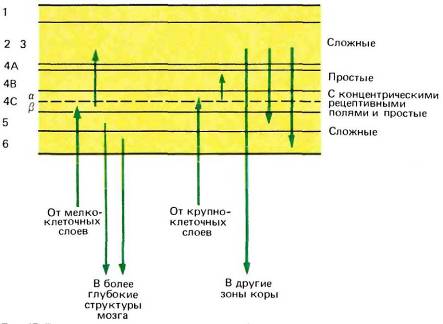
Рис. 67. Примерная схема размещения клеток физиологически различного типа в разных слоях стриарной коры.