

84 Глава 4
простейшей из таких схем, сложная клетка получает входные сигналы от множества простых клеток, рецептивные поля которых имеют одну и ту же ориентацию, но размещены, частично перекрывая друг друга, по всему полю сложной клетки, как показано на рис. 47. Если связи от простых клеток к сложным возбуждающие, то всякий раз, когда в рецептивное поле сложной клетки попадает стимул в виде линии, возбуждаются некоторые простые клетки. В результате будет возбуждаться и сложная клетка.
Обычно в ответ на неподвижную линию сложная клетка дает короткий импульсный разряд (даже если стимул остается включенным). В этом случае мы говорим, что происходит адаптация ответа. Если же перемещать линию в рецептивном поле сложной клетки, наблюдается непрерывный разряд: адаптация преодолевается в результате последовательного срабатывания новых простых клеток.
Читатель, должно быть, заметил, что обе приведенные схемы связей — от клеток с круглыми рецептивными полями к простым клеткам (рис. 44) и от простых клеток к сложным (рис. 47) —предполагают использование возбуждающих связей. Однако в этих двух случаях процессы возбуждения должны быть совершенно различными. В первой из этих схем требуется суммирование одновременных сигналов от клеток с круглыми рецептивными полями, лежащих на одной линии. Во второй же схеме для активации сложной клетки движущимся стимулом необходимо последовательное возбуждение множества простых клеток. Было бы интересно выяснить, с какими морфологическими различиями (если они есть) связано это различие в механизме суммации.
Дирекционалыгая избирательность
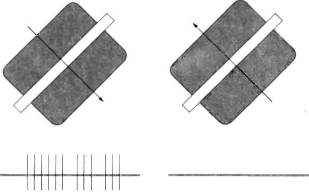
Многие сложные клетки лучше реагируют на движение стимула в одном направлении, чем в противоположном. Различие в реакции часто бывает весьма резким — при одном направлении движения возникает энергичный ответ, а при обратном направлении клетка вообще не отвечает (рис. 48). Как выяснилось, примерно 10—20% клеток в верхних слоях стриарной коры проявляют заметную дирекциональную избирательность. Остальные клетки, по-видимому, не обладают такой избирательностью — мы тщательно исследовали ответы клеток с помощью ЭВМ, пытаясь обнаружить хотя бы небольшое различие в ответах на движение стимула в противоположных направлениях. Таким образом, существуют, видимо, два разных класса клеток — один из них четко проявляет дирекциональную избирательность, другому она не свойственна.
Рис. 48. Реакции данной сложной клетки на движение оптимально ориентированной полосы в противоположных направлениях различны. Длительность каждой записи около 2 секунд. (Для таких клеток не особенно важно, насколько быстро движется полоса; обычно клетка не отвечает только при очень быстром движении, когда полоса кажется размытой, или же при таком медленном, что его вообще трудно заметить.)
ПЕРВИЧНАЯ ЗРИТЕЛЬНАЯ КОРА
85
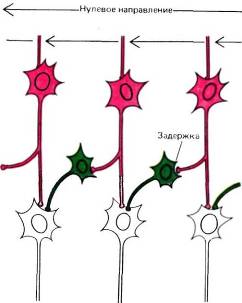
Рис. 49. Эту схему предложили X. Барлоу и У. Левик для объяснения свойства дирекциональной чувствительности. Синапсы, которые красные клетки образуют на зеленых клетках, — возбуждающие, а синапсы, образуемые на белых клетках, — тормозные. Мы предполагаем, что три белые клетки (внизу) конвергируют на одну «главную» клетку.
Если прослушивать импульсную реакцию клетки с сильно выраженной дирекциональной избирательностью, то создается впечатление, что при движении линии в одном направлении стимул как бы резко подталкивает клетку и заставляет ее разряжаться, а при движении в противоположном направлении происходит как бы сбой и стимул становится неэффективным (это напоминает ситуацию, когда вы, заводя часы, поворачиваете головку заводного механизма назад и раздается характерный треск).
Мы не знаем, как устроены входные сети таких клеток с дирекциональной избирательностью. Возможно, что на вход такой клетки подключены простые клетки, реакции которых на движение стимула в противоположных направлениях неравнозначны, асимметричны. Рецептивные поля таких простых клеток асимметричны, как, например, поле, представленное на рис. 42, В. Другую возможную схему предложили в 1965 году Г. Барлоу и У. Левик для объяснения дирекциональной избирательности некоторых клеток в сетчатке кролика — клеток, которых, по-видимому, нет у обезьяны. Если применить их схему к сложным клеткам коры, то нужно предположить наличие промежуточного слоя клеток, расположенных между простыми и сложными клетками (как на рис. 49). Представим себе, что клетка из промежуточного слоя имеет возбуждающий вход от одной простой клетки и тормозный вход — от другой клетки, рецептивное поле которой непосредственно примыкает к предыдущему, и притом всегда с одной стороны. Предположим далее, что тормозный путь включает некоторую задержку, возможно, за счет включения еще одной промежуточной клетки. В таком случае, если стимул движется в одном направлении, скажем справа налево (как на рис. 49, иллюстрирующем модель Барлоу и Леви-ка), то промежуточная клетка возбуждается сигналом с одного из своих входов как раз в тот момент, когда действует торможение, приходящее от другой клетки, рецептивное поле которой только что пересек стимул. Возбуждающий и тормозный эффекты нейтрализуют друг друга, и в результате клетка не дает импульсного разряда. Если же стимул движется в обратном направлении, торможение возникает слишком поздно для того, чтобы предотвратить импульсный разряд. Если множество таких промежуточных клеток конвергирует на