

3. Построение клеточной оболочки и перегородки
Процесс образования новой клеточной оболочки (мембраны и клеточной стенки) включает две основные стадии:
пролиферация поверхностных клеточных структур, т.е. накопление в них нового материала в динамике клеточного цикла;
сегрегация поверхностных структур клетки, т.е. включение нового материала в предшествующие структуры по местам локализации соответствующих центров (сайтов) включения.
Включение белков в мембраны бактерий следует сложной кинетике, свидетельствующей о накоплении предварительно образованных белков в цитоплазме в период подготовки клеточного деления и быстрой их мобилизации в процессе деления. В период деления возрастает активность некоторых литических ферментов (в частности, муреингидролаз), участвующих в образовании брешей в предшествующем каркасе клеточной стенки: необходимых для включения новых ее фрагментов. Таким образом, регуляция активности этих ферментов осуществляется путем перевода их в скрытое, латентное состояние с последующей мобилизацией в определенный момент времени.
Консервативный механизм сегрегации предусматривает включение новых фрагментов на одном из полюсов клетки, при этом поверхностные слои всех дочерних клеток состоят из новообразованного материала. Такой способ сегрегации реализуется в процессе почкования, характерного для некоторых микроорганизмов, но не при обычном клеточном делении.
Полуконсервативный механизм сегрегации предполагает локализацию сайтов включения новых фрагментов в экваториальной зоне клетки. В результате появляются дочерние клетки с полностью новообразованными поверхностными структурами, но лишь при второй генерации после начала наблюдения.
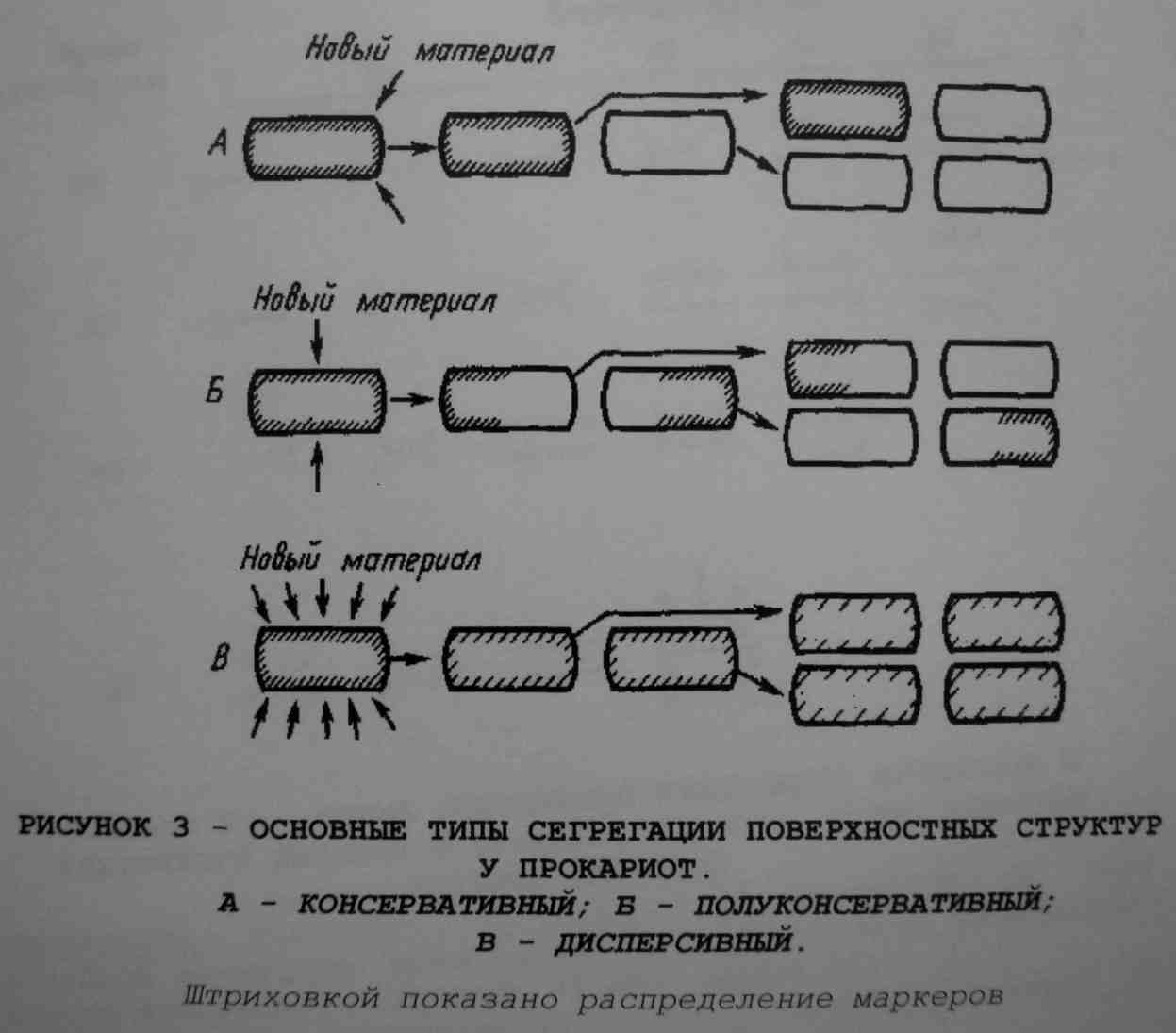 Дисперсивный
механизм сегрегации
предусматривает включение новых
фрагментов на множестве сайтов по всей
поверхности клетки, что приводит к
постепенному «разбавлению» старых
структур новыми и равномерному
распределению маркеров на поверхности
дочерних клеток.
Дисперсивный
механизм сегрегации
предусматривает включение новых
фрагментов на множестве сайтов по всей
поверхности клетки, что приводит к
постепенному «разбавлению» старых
структур новыми и равномерному
распределению маркеров на поверхности
дочерних клеток.
Процесс построения новой клеточной перегородки завершает клеточный цикл. В раскрытии механизмов регуляции этого процесса важный вклад внесло изучение специфических мутантов, из которых наибольшую ценность представляют «условные» мутанты. У мутантов данного типа процесс протекает нормально при обычных физиологических условиях (пермессивных условиях), а при непермессивных условиях (повышенная температура, действие радиации) процесс подавляется. В частности, для изучения регуляции процесса построения клеточной перегородки использовались мутанты Е. coli и Вас subtilis, образующие «миниклетки» (min-мутанты). Существование min-мутаций позволило определить, что сайт, ответственный за образование перегородки и локализующийся в процессе деления в экваториальной зоне клетки, сохраняется на полюсах дочерних клеток при последующем их расхождении. В норме сайты построения перегородки, локализованные на полюсах клетки, выключаются (блокируются) и могут функционировать наряду с экваториальными сайтами лишь у min -мутантов. Это обстоятельство позволяет постулировать существование определенного компонента - активатора сборки клеточной перегородки. На протяжении клеточного цикла образуется ограниченное количество («квант») этого активатора, достаточное для функционирования лишь одного сайта (при этом активатор полностью расходуется). В нормальных клетках количество «квантов» активатора и количество функционирующих сайтов построения перегородки совпадает, а у min-мутантов количество сайтов превышает количество «квантов» активатора.