4. Регуляция метаболической активности за счёт компартментализации ферментов и их взаимодействия с клеточными мембранами
Эукариотные и прокариотные клетки имеют пространственные «отсеки», в которых локализована часть их ферментативного аппарата. Так, у грамотрицательных бактерий некоторые гидролазы локализованы в периплазматическом пространстве (между внешней и цитоплазматической мембранами). Эти обстоятельства создают возможность регуляции ферментативной активности путем управления скоростью проникновения в «отсек» субстрата или выхода из него фермента (компартментация).
Многие белки и в том числе ферменты способны обратимо взаимодействовать с клеточной мембраной, что приводит к изменению физико-химических свойств белков и их ферментативной активности (аллотопия). Так, гидрофобные взаимодействия липидов и белков могут переводить последние в неактивное (латентное) состояние, напротив, электростатические взаимодействия вызывают активирование некоторых ферментов.
В свою очередь степень и характер взаимодействия ферментов с другими мембранными белками и липидами в определенной степени зависят от внутриклеточной концентрации электролитов, а следовательно, могут регулироваться при изменении физиологических условий.
Ферменты, катализирующие серию последовательных реакций, иногда образуют ансамбли, локализованные в цитоплазме (дегидрогеназы -кетокислот) или в клеточной мембране (ферменты дыхательной и фотосинтетической цепей). Регуляция в таких ансамблях имеет свои особенности, так как продукт, образуемый на предыдущей стадии, «подхватывается» следующим ферментом без выхода в окружающую среду.
Лекция №6
РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ БИОMEMБPAH. ПACСИВНАЯ ПРОНИЦАЕМОСТЬ И ТРАНСПОРТНЫЕ ФУНКЦИИ ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ БАКТЕРИЙ. ОРГАНИЗАЦИЯ И РЕГУЛЯЦИЯ ТРАНСПОРТНЫХ ПРОЦЕССОВ НА УРОВНЕ БИОСИНТЕЗА. СБОРКА И ФУНКЦИОНИРОВАНИЕ КОМПОНЕНТОВ ТРАНСПОРТНЫХ СИСТЕМ. ЭНЕРГЕТИКА ТРАНСПОРТНЫХ ПРОЦЕССОВ У МИКРООРГАНИЗМОВ
1. Пассивная проницаемость и транспортные функции цитоплазматической мембраны бактерий
Основную роль в транспорте веществ из окружающей среды в клетки большинства микроорганизмов выполняют компоненты аппарата, локализованного в цитоплазматической мембране.
Наружная мембрана большинства микроорганизмов является барьером для проникновения большинства гидрофильных (и некоторых гидрофобных) веществ. Избирательная проницаемость наружной мембраны обеспечивается двумя основными способами: 1) образованием гидрофильных «каналов», или «пор», 2) с помощью специальных структурных белков (поринов), обеспечивающих проникновение гидрофильных веществ с молекулярной массой до нескольких сотен дальтон, 3) путем локализации в наружной мембране ряда специфических транспортных систем (для ионов желе мальтозы, витамина В12, нуклеозидов).
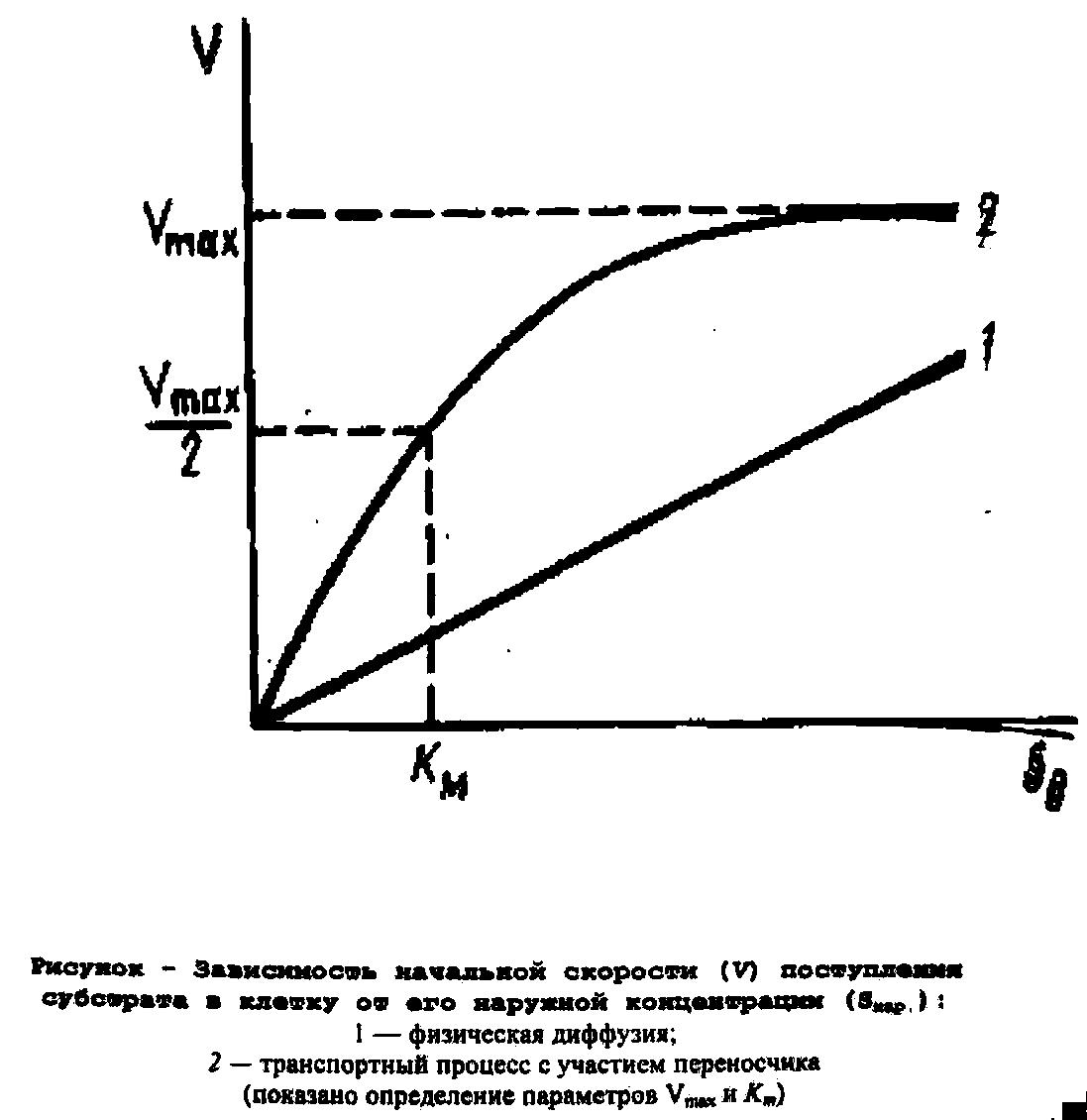
Пассивная проницаемость
 Пассивная
проницаемость
- проникновение
через мембрану веществ без прямого
участия мембранных посредников за счет
теплового движения молекул (физическая
диффузия).
Конечным итогом физической диффузии
обычно является уравнивание наружной
(SHАР)
и внутриклеточной (SBHУTР)
концентраций вещества. Начальная
скорость физической диффузии линейно
зависит от внешней концентрации вещества,
точнее от градиента концентраций (S
= SBHУTР
- SHАР),
а изменение температуры (в пределах
физиологического диапазона) мало влияет
на скорость процесса (рис.).
Пассивная
проницаемость
- проникновение
через мембрану веществ без прямого
участия мембранных посредников за счет
теплового движения молекул (физическая
диффузия).
Конечным итогом физической диффузии
обычно является уравнивание наружной
(SHАР)
и внутриклеточной (SBHУTР)
концентраций вещества. Начальная
скорость физической диффузии линейно
зависит от внешней концентрации вещества,
точнее от градиента концентраций (S
= SBHУTР
- SHАР),
а изменение температуры (в пределах
физиологического диапазона) мало влияет
на скорость процесса (рис.).
Таким путем поступают в клетку вода, неполярные и малополярные молекулы газов (О2, Н2, N2) и углеводороды.
Транспортные функции цитоплазматической мембраны
Для большинства гидрофильных природных субстратов (сахаров, аминокислот, органических кислот и т.д.) коэффициент диффузии через двойной липидный слой мембраны имеет очень низкую величину. Скорость диффузии этих веществ недостаточна для обеспечения нормальной скорости метаболических процессов. В связи с этим к функциям мембраны относится проведение таких транспортных процессов, как облегченная диффузия и активный транспорт.
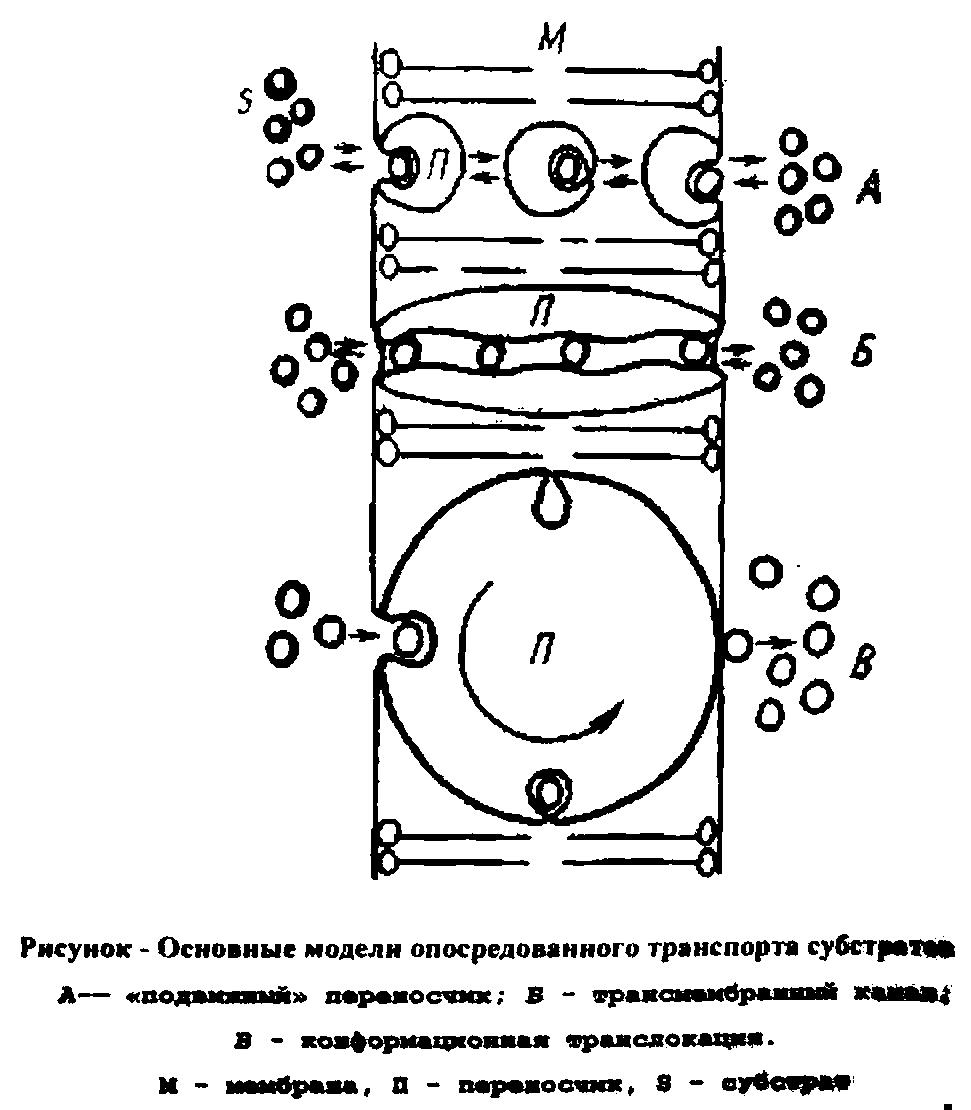
Облегченная диффузия
При облегченной диффузии поступление веществ в микробную клетку осуществляется с помощью специфических мембранных переносчиков. Это мембранные белки, имеющие общее название пермеазы, которые в ряде случаев индуцируются своими субстратами. Переносимое вещество связывается с пермеазой снаружи и освобождается внутри клетки. При облегченной диффузии, так же как и при пассивной диффузии, переносимый субстрат движется по градиенту концентрации (т.е. от более высокой к более низкой концентрации), причем ни один из этих процессов не требует метаболической энергии.
В результате этого транспортного процесса происходит уравнивание внешней (SHАР) и внутренней (SBHУТР) концентраций переносимого вещества, либо незначительное превосходство последней (SBHУТР> SHАР).
Активный (концентрирующий) транспорт
Системы активного транспорта могут создавать внутри микробной клетки концентрации растворенных веществ, которые в тысячи раз превышают их концентрации во внешней среде. Это обеспечивает возможность развития микроорганизмов в условиях низкого содержания питательных веществ. Активный транспорт характеризуется специфичностью по отношению к субстрату, которая обеспечивается мембранным переносчиком. Когда переносчик обращен к внешней поверхности мембраны, он имеет высокое сродство к субстрату, а когда обращен к ее внутренней поверхности - низкое. Благодаря этому субстрат «накачивается» в клетку. Этот процесс сопряжен с затратой метаболической энергии, обеспечивающей диссоциацию субстрата и переносчика на внутренней поверхности мембраны. Например, с помощью механизма активного транспорта при участии -галактозидпермеазы в клетку поступает лактоза.
Этот транспортный процесс требует затраты энергии, в результате чего происходит аккумуляция вещества в клетке, и внутренняя концентрация (SBHУТР) переносимого вещества значительно превосходит наружную концентрацию (SHАР), т.е. SBHУТР >> SHАР.
Стереоспецифичность - это отличительная особенность транспортных процессов от физической диффузии. Она означает, что близкие по химической структуре вещества конкурируют при транспорте за общий переносчик. Ограниченное количество молекул переносчика в мембране приводит к тому, что зависимость начальной скорости транспортного процесса от концентрации субстрата описывается уравнением ферментативной кинетики Михаэлиса-Ментен с аналогичными параметрами (Кт и vmax), а графически выражается гиперболой.