

2. Роль энергетического заряда в регуляции клеточного метаболизма
Аденилаты выполняют в клетке важные регуляторные функции, представляя собой общие для катаболизма и анаболизма сигналы, обеспечивающие нужное соотношение между получением энергии и процессами биосинтеза. Внутриклеточное содержание AТФ, AДФ и AMФ (или, точнее, соотношение между этими тремя аденилатами) определяет скорость отдельных реакций, а тем самым и сложных процессов распада и синтеза.
В катаболических (энергетических) путях метаболизма (в промежуточном обмене) отрицательным эффектором для ферментов часто служит соединение, являющееся аккумулятором энергии (АТФ, пирофосфат и др.), тогда как другие компоненты аденилатной системы (АМФ, АДФ) могут выступать в качестве положительных эффекторов. Следовательно, активность рассмотренных ферментов определяется общим «энергетическим зарядом» клетки. Энергетический заряд клетки (потенциалом Аткинсона) вычисляется по формуле:
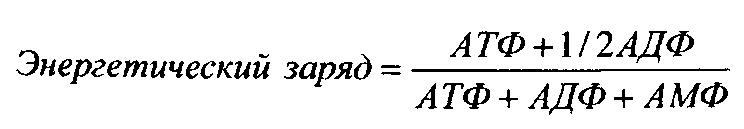
Его величина относительна, варьирует в пределах от 0 до 1 и характеризует молярную долю АТФ в общем балансе адениновых нуклеотидов, то есть служит мерой ее обеспеченности высокоэнергетическими соединениями в расчете на общее число «аденозиновых единиц» (аденилатов). Аллостерические ферменты активируются или ингибируются тем или иным аденилатом и именно это обеспечивает согласованную регуляцию всего метаболизма клетки. Если, например, энергетический заряд клетки возрастает, то активность катаболических ферментов снижается, а активность ферментов, участвующих в процессах синтеза, увеличивается. При уменьшении энергетического заряда наблюдается обратная картина. Энергетический заряд большинства живых клеток находится в пределах 0,75-0,90 (у растущих клеток равняется примерно 0,8). Клетки Е. coli погибают, когда значение этой величины меньше 0,5.
Адениннуклеотиды регулируют активность РНК-полимеразы, изменяя ее сродство к различным промоторам. Повышение содержания АДФ и АМФ и снижение эффективности процессов регенерации АТФ приводит к подавлению синтеза рРНК и других компонентов белоксинтезирующего аппарата, что представляется целесообразным в неблагоприятных для роста условиях. При высокой внутриклеточной концентрации АТФ и ГТФ, отражающей благоприятное энергетическое состояние клетки, синтез рРНК и других компонентов аппарата трансляции стимулируется. В результате регуляции процессов синтеза и распада АТФ в клетках живых организмов поддерживается стационарное энергетическое состояние.
3. Регуляторные эффекты Пастера и Крэбтри
 Регуляторный
«эффект Пастера»
- торможение гликолиза дыханием -
объясняется энергозависимой конкуренцией
за АДФ и фосфат между системами
фосфорилирования в дыхательной цепи
(окислительное
фосфорилирование) и
на уровне субстрата (субстратное
фосфорилирование).
Хорошо известно, что факультативно
анаэробные клетки, растущие в анаэробных
условиях, потребляют сравнительно очень
большие количества глюкозы (и образуют
большие количества продуктов брожения).
Если же ввести в среду кислород, то
клетки начнут немедленно его поглощать
и уровень потребления глюкозы резко
снизится. Это снижение скорости
гликолитического расщепления глюкозы
с началом дыхания и носит название
эффекта Пастера.
Регуляторный
«эффект Пастера»
- торможение гликолиза дыханием -
объясняется энергозависимой конкуренцией
за АДФ и фосфат между системами
фосфорилирования в дыхательной цепи
(окислительное
фосфорилирование) и
на уровне субстрата (субстратное
фосфорилирование).
Хорошо известно, что факультативно
анаэробные клетки, растущие в анаэробных
условиях, потребляют сравнительно очень
большие количества глюкозы (и образуют
большие количества продуктов брожения).
Если же ввести в среду кислород, то
клетки начнут немедленно его поглощать
и уровень потребления глюкозы резко
снизится. Это снижение скорости
гликолитического расщепления глюкозы
с началом дыхания и носит название
эффекта Пастера.
Эффект Пастера обусловлен подавлением активности фермента фосфофруктокиназы, который катализирует реакцию, лимитирующую общую скорость гликолиза, а именно фосфорилирование фруктозо-6-фосфата с образованием фруктозо-1,6-дифосфата. Фосфофруктокиназа представляет собой поливалентно регулируемый аллостерический фермент, который активируется АДФ и подавляется АТФ и цитратом. Знание аллостерических свойств фосфофруктокиназы позволяет сделать следующий вывод: если аэробно растущие дрожжевые клетки лишить кислорода и приостановить таким образом окислительное фосфорилирование, то в клетке снизится соотношение концентраций АТФ/АМФ, что приведет к повышению активности фермента и соответственно к ускорению реакции. Таким образом, снижение активности фосфофруктокиназы связано с ускорением образования АТФ и началом функционирования ЦТК при переходе к аэробному окислению глюкозы.
Регуляторный «эффект Кребтри» - торможение дыхания гликолизом - явление, обратное «эффекту Пастера», также объясняется энергозависимыми конкурентными взаимоотношениями систем окислительного и субстратного фосфорилирования за АДФ и фосфат. Одна из точек зрения состоит в том, что эффект Кребтри обусловлен активацией гликолиза высокими концентрациями глюкозы, что приводит к избытку продуктов гликолиза, которые, главным образом в виде ацетата, экскретируются в среду. Другая точка зрения заключается в том, что эффект Кребтри можно моделировать, применяя довольно низкие (миллимолярные) концентрации глюкозы, регистрируя при этом явление «дыхательного контроля» у бактерий. Обе точки зрения сходятся в одном - в основе механизма эффекта Кребтри лежит регуляторная способность факультативных анаэробов модулировать в широких пределах поток через гликолиз, чтобы компенсировать при необходимости нефункционирующий ЦТК усилением субстратного фосфорилирования и по той же причине иметь механизм сброса избытка продуктов гликолиза при анаэробиозе.
Таким образом, скорость катаболизма определяется не концентрацией тех или иных субстратов в среде, а потребностью микробной клетки в энергии.