

15.Глікопротеїни — гібридні молекули, що є складними білками, з поліпептидною
основою яких ковалентно зв’язані олігосахаридні (гліканові) ланцюги.
Глікопротеїни та близькі до них за будовою гліколіпіди створюють поверхне-
воклітинний глікокалікс, компоненти якого беруть участь у міжклітинній взаємодії,
регуляції клітинного поділу та її порушеннях при злоякісному рості. Глікопротеїни
клітинної поверхні реалізують функцію взаємного розпізнавання клітин, що має
особливе значення в процесах імунітету: полісахариди клітинної мембрани є
антигенами, відносно яких розвиваються реакції клітинного імунітету, зокрема
при трансплантації органів та тканин.
Вміст вуглеводного компоненту в більшості глікопротеїнів невеликий, але іноді
складає до 50-80 % маси молекули. Кількість вуглеводних ланцюгів у складі
окремих молекул глікопротеїнів змінюється від одного до кількох десятків. У свою
чергу, кожен олігосахаридний ланцюг глікопротеїнів містить від 1 до 15 монозних
залишків. Ці ланцюги можуть бути лінійними або розгалуженими.
Найчастіше у вуглеводному фрагменті глікопротеїнів організму людини зустрі-
чаються такі цукри, як галактоза (Gal), глюкоза (Glc), маноза (Man), фукоза (Fuc),
N-ацетилгалактозамін (GalNAc), N-ацетилглюкозамін (GlcNAc), N-ацетилнейра-
мінова (сіалова) кислота (NeuAc)
Подібно до протеогліканів, у молекулах глікопротеїнів олігосахаридні ланцюги
зв’язані з поліпептидною частиною молекули за рахунок:
1) O-глікозидного зв’язку між цукром та OH-групою серину або треоніну;
2) N-глікозидного зв’язку між цукром та амідною групою аспарагіну.
Представниками О-зв’язаних глікопротеїнів є муцини (слизові білки) і деякі
білки крові та біомембран. У складі цих глікопротеїнів цукром, що безпосередньо
зв’язаний з Ser або Thr пептидного ланцюга, є GalNAc.
N-зв’язані глікопротеїни мають як олігосахаридний фрагмент, що безпосередньо
зв’язаний з NH-групою аспарагіну, пентасахарид (Man)3(GlcNAc)2, з яким сполучені
різні за будовою зовнішні олігосахаридні фрагменти. Ці зовнішні розгалужені
олігосахаридні ланцюги нагадують молекулярні “антени”, що відіграють головну
роль у взаємодії клітин між собою та з іншими білковими і полісахаридними струк-
турами.
16 Нуклеотиди — трикомпонентні сполуки, які побудовані з азотистої основи
пуринового чи піримідинового ряду, залишків пентоз (рибози або дезоксирибози) та
фосфату.
В основі структури азотистих основ нук-
леотидів лежать ароматичні гетероциклічні
сполуки пурин та піримідин.
Пуринові основи нуклеїнових кислот
У гідролізатах нуклеїнових кислот постійно містяться дві пуринові основи — аденін
(
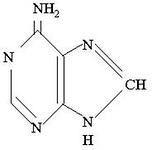
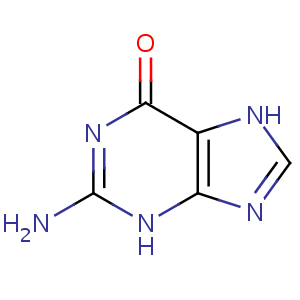 А)
та гуанін
(Г)
А)
та гуанін
(Г)
Піримідинові основи нуклеїнових кислот
До складу нуклеотидів нуклеїнових кислот входять три головні піримідинові основи:
урацил (У), тимін (Т), цитозин (Ц).
Оксипохідні пурину та піримідину можуть перебувати у двох таутомерних фор-
мах — лактамних і лактимних, — залежно від рН середовища. У складі нуклео-
тидів нуклеїнових кислот оксипохідні пурину та піримідину знаходяться в лактамній
формі, що сприяє утворенню міжмолекулярних водневих зв’язків між пуринами
та піримідинами окремих ланцюгів у дволанцюговій структурі молекул ДНК та в
одноланцюгових РНК.


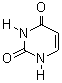
цитозин тимін урацил
17.Мінорні нуклеотиди
Крім зазначених вище основних п’яти азотистих основ (двох пуринових та трьох
піримідинових), до складу деяких нуклеїнових кислот входять у відносно незначних
кількостях додаткові (мінорні) азотисті основи та відповідні їм мінорні нуклеотиди.
Найбільша кількість мінорних нуклеотидів зустрічається в молекулах транспортних
РНК (тРНК) — до 5 % загального нуклеотидного складу. До мінорних нуклеотидів
належать метильовані похідні звичайних азотистих основ, зокрема, 1-метиладенін,
2-метиладенін, 6-диметиладенін, 1-метилгуанін, 7-метилгуанін, 1-метилурацил,
5-оксиметилурацил, 3-метилцитозин тощо. ДНК людини містять значну кількість
5-метилцитозину, інформаційні РНК — N-метильовані похідні аденіну та гуаніну.
Нуклеотидом незвичайної структури, що входить до складу тРНК, є псевдоуридин
() — нуклеотид, в якому рибоза приєднана до урацилу в 5-му положенні, тобто не
азот-вуглецевим, а вуглець-вуглецевим зв’язком.
Біологічні функції мінорних нуклеотидів до кінця не з’ясовані.
Біохімічні функції вільних нуклеотидів:
1. Участь в енергетичному обміні (реакціях окисного фосфорилювання) — функ-
цію виконують нуклеотиди аденілової системи (АТФ, АДФ). Ці ж нуклеотиди та
АМФ можуть бути алостеричними модуляторами певних регуляторних ферментів,
зокрема ферментів гліколізу, біосинтезу пуринових нуклеотидів.
2. Участь у метаболічних реакціях у ролі коферментів, зокрема:
– НАД, НАДФ, ФАД, ФМН — у реакціях біологічного окислення;
– УТФ, УДФ — у реакціях біосинтезу глікогену;
– ЦТФ, ЦДФ — у біосинтезі гліцерофосфоліпідів.
18. Первинна структура нуклеїнових кислот
Як уже зазначалося, всі класи нуклеїнових кислот (ДНК та РНК) є високо-
молекулярними сполуками, основою первинної структури яких є полінуклео-
тидний ланцюг, побудований із мономерів — нуклеотидів.
Окремі нуклеотиди сполучаються між собою в полінуклеотидний ланцюг за рахунок
фосфодіефірних зв’язків, що утворюються між 3'- та 5'- гідроксильними групами пентоз
(рибоз або дезоксирибоз) сусідніх нуклеотидів.
Біологічні функції ДНК:
1. Збереження спадкової інформації.
Кількість ДНК у соматичних та статевих клітинах організму людини є сталою
величиною, яку ці клітини отримують у процесах запліднення батьківських гамет та
подальшого поділу зиготи.
2. Передавання генетичної інформації нащадкам.
Подвоєння молекул ДНК у процесі реплікації та передавання нащадкам копій
материнських молекул є основою консерватизму спадковості, збереження протягом
багатьох поколінь основних біологічних ознак виду.
3. Реалізація генетичної інформації.
Ця біологічна функція здійснюється за рахунок передачі закодованої в ДНК інформації
молекулам інформаційних (матричних) РНК (транскрипції) та подальшої розшиф-
ровки цієї інформації при синтезі білків (трансляції).
Сукупність зазначених біологічних функцій ДНК та механізмів їх реалізації отримала
назву — центральна догма молекулярної біології (Ф.Крік) (рис. 3.1):
Експериментальне доведення генетичної ролі ДНК (феномен трансформації)
У 1928 р. англійським мікробіологом Ф. Гріффітом (F. Griffith) при вивченні двох
штамів пневмококів Streptococcus pneumoniae — патогенного, що викликає
пневмонію в людини та мишей (капсульної S-форми), та непатогенного мутанта
(безкапсульної R-форми) — було відкрито явище трансформації. Вона полягала
в можливості перетворення непатогенної R-форми в патогенну S-форму:
трансформація
R S
Гріффіт встановив, що трансформація пневмококів відбувається за умов
взаємодії в організмі піддослідних тварин (мишей) вбитої нагріванням S-форми
(патогенної) з живою непатогенною R-формою. Був зроблений висновок, що у
вбитих нагріванням вірулентних клітинах пневмококів (штам S) присутній певний
трансформуючий фактор, який, проникаючи в живі невірулентні клітини (штам
R), змінює біологічні властивості останніх, надаючи їм властивість патогенності,
до того ж ця властивість є спадковою:
трансформуючий фактор
Пізніше, в 1944 р., групою дослідників із Рокфелерівського інституту (США) —
О.Евері, К.Мак-Леодом та М.Мак-Карті (O.Avery, C.McLeod, M.McCarthy) при
дослідженні хімічної природи екстрактів патогенних пневмококів, які спричиняють
трансформацію, було доведено, що трансформуючим фактором пневмококів є
клітинна ДНК:
ДНК
Таким чином, у результаті досліджень О.Евері та співавторів генетичне
поняття “фактор спадковості”, або “ген” уперше набуло конкретного моле-
кулярного змісту — ним виявилась дезоксирибонуклеїнова кислота.