

Влияние факторов 1нешней среды на рост растений
менее глубоко — подпрограмма образования флоэмы. Клетки, занимающие промежуточное положение, становятся камбиальными, т. е. делятся по замкнутому циклу, формируя элементы ксилемы и флоэмы.
Несмотря на некоторую механистичность представленной схемы, гипотеза «морфогенетических тестов» Боннера находит поддержку в экспериментах последних лет. У растений найдены рецепторы фитогормонов, позволяющие клеткам оценивать их состав и количество в окружающей среде. При культивировании растительных клеток в искусственной среде установлен «эффект массы». Единичная изолированная клетка редко переходит к делению. Чем гуще высеяны клетки (например, на поверхность питательного агара), тем большее их число начинает делиться. На рис. 11.14, А показано, что если яйцеклетки фукуса помещены близко друг от друга, то ризоиды образуются в сторону центра группы («групповой эффект»). Это явление можно объяснить тем, что каждая яйцеклетка синтезирует и выделяет в окружающую среду ИУК, и концентрация этого фитогормона в центре группы оказывается более высокой, чем снаружи. Как уже говорилось, ауксин индуцирует у яйцеклеток фукуса образование ризоидов. Таким образом, тест на величину Труппы клеток может быть опосредован концентрацией фитогормонов или других физиологически активных веществ, выделяемых клетками.
На рост растений оказывают влияние многие факторы внешней среды. Прежде всего это физические факторы: свет (его интенсивность, качество, продолжительность и периодичность), температура (величина и периодичность), сила тяжести, газовый состав, магнитное поле, влажность, питательные вещества (минеральные и органические) и механические воздействия
Влияние
света или постоянной темноты в течение
8 суток на рост и морфогенез проростков
фасоли

Темнота Свет
(например, ветер). Кроме того, находясь в составе растительных сообществ, растение испытывает влияние продуктов жизнедеятельности других растений (аллелопатия), а также физиологически активных веществ микроорганизмов (антибиотиков, ростовых веществ).
Свет. Рост растений может происходить на свету и в темноте. Зеленые органы высших растений в темноте приобретают ряд морфологических особенностей, отличающих их от растений, выращиваемых на свету (рис. 11.17). Такие растения называются этиолированными. У двудольных при росте в темноте сильно вытягиваются междоузлия, а листовые пластинки недоразвиты; у проростков кукурузы вытягиваются мезокотили. У этиолированных растений, как правило, недоразвиты механические ткани и устьица. Растения лишены хлорофилла и имеют бледно-желтый цвет из-за присутствия каротиноидов.
Явление этиоляции не связано с отсутствием хлорофилла или недостатком питания: этиоляция отмечена у многих грибов и у проростков хвойных, способных зеленеть в темноте.
И МСГШШ-
при.. f^cyubvx
темп*
МСГШШ-
при.. f^cyubvx
темп*
Для устранения этиоляции достаточно кратковременного (5—10 мин) освещения растений один раз в сутки, что не влияет на образование хлорофилла. Предотвращение этиоляции регулируется фитохромом (красный — дальний красный свет) или системой фитохром — флавопротеин (реакция на дальний красный — синий свет). Наиболее сильный морфогене-тический эффект оказывает сине-фиолетовая часть спектра.
Этиоляция — важная приспособительная реакция проростков растений, находящихся в почве: за счет быстрого вытягивания мезокотиля, гипокотиля или эпикотиля побег выносится на поверхность, к свету. При освещении этиолированных растений торможение роста растяжением наступает через несколько минут, остальные эффекты света (развертывание первого листа, закладка следующих листьев, рост волосков и т. д.) проявляются лишь через несколько часов.
Температура. Как и другие процессы, рост растений зависит от температуры. В пределах от 0 до 35 °С влияние температуры на рост подчиняется правилу Вант-Гоффа, но свыше 35 — 40 °С скорость роста снижается. В зависимости от приспособленности к действию температур различают растения теплолюбивые с минимальными точками для роста выше 10 °С и оптимальными при 30 — 40 °С и холодостойкие с минимальными точками от 0 до 5°С и оптимальными при 25 — 31 °С. Максимальные температуры для роста большинства растений находятся в пределах 35 — 45 °С. Оптимальной называют температуру, при которой рост осуществляется наиболее быстро. Но высокая скорость роста не всегда благоприятна для формирования растения. При «выгонке» растений умеренного пояса при высоких температурах стебель становится слабым, а листья тонкими. В поле такие растения полегают.
Газовый состав. Для роста растений необходимо присутствие кислорода. Однако кратковременное снижение наполовину его содержания значительно не сказывается на росте растений. Даже в условиях длительного недостатка кислорода в зоне корней при затоплении рост продолжается, хотя и с более низкой скоростью. Это связано, как уже отмечалось, с включением адаптационных механизмов, позволяющих использовать 02 нитратов, воздухоносных тканей и т. д.
Избыток С02 в атмосферном воздухе приводит к увеличению растяжимости клеточных стенок и кратковременному (в течение нескольких часов) усилению роста тканей (эффект «кислого роста»). Выяснено, что влияние С02 на рост основано на способности снижать рН клеточных стенок и таким образом индуцировать рост клеток. Эффект С02 не зависит от присутствия кислорода. Именно с эффектом «кислого роста» наряду с затенением может быть связано чрезмерное удлинение нижних междоузлий злаков в загущенных посевах и вследствие этого полегание растений.
Водный режим. Процесс роста клеток растений растяжением осуществляется путем вакуолизации при поступлении в клетки воды. Поэтому недостаточное снабжение клеток водой задерживает рост. Корни способны расти только в достаточно влаж-
11.6
Использование синтетических
регуляторов
роста в растениеводстве
ной почве, почти насыщенной водяным паром, с осмотическим давлением почвенного раствора не более 1 — 1,5 МПа. Надземные части растений всегда находятся в более сухом воздухе с влажностью 50—70%. От потери воды ткани наземных органов защищены кутикулярно-эпидермальным слоем. Поэтому в мезофилле листьев упругость водяного пара обычно не бывает ниже 98 — 99% относительной влажности. При длительном недостатке воды в тканях фаза растяжения заканчивается быстрее, что приводит к укорочению стебля и корня, к уменьшению размеров листьев, к их мелкоклеточности и т. д. Недостаток воды до начала и в период стеблевания злаков (осуществляемого за счет роста растяжением) особенно резко снижает урожай.
Минеральное питание. На росте растений благоприятно сказывается высокое содержание в почве минеральных элементов, особенно азота. Однако способствующие быстрому росту высокие концентрации азота задерживают процессы дифферен-цировки, в частности закладку цветков. Высокий минеральный фон приводит к разрастанию вегетативных органов и необходим при наращивании зеленой массы кормовых растений. Но чрезмерное удобрение снижает урожай плодов и зерна. Поэтому по методу В. А. Чеснокова в гидропонной культуре лучший урожай огурцов, моркови, клубней картофеля получается при одноразовом или периодическом голодании растений, особенно по азоту.
Синтетические регуляторы роста для практических целей начали использоваться с 40-х годов. Их роль в сельскохозяйственном производстве год от года возрастает. Особенно важное значение регуляторы роста приобрели в плодоводстве и при возделывании пшеницы. Экономическая выгода от их использования многократно превысила те затраты, которые были сделаны при изучении этих физиологически активных веществ.
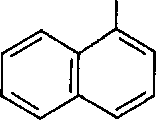
Регуляторы роста ауксинового типа. Некоторые синтетические соединения влияют на растения подобно ИУК, однако они действуют, как правило, в меньших концентрациях и более продолжительно, так как не разрушаются и не связываются в тканях так быстро, как природная ИУК. Эти вещества относятся к индольным, фенольным соединениям и к нафтилалкил-карбоновым кислотам:
(CHjlj-CHjCOOH
О—СН2СООН
С1
С1![]()
![]()
СН2СООН
2,4-дихлорфенокси-уксусная кислота
1-нафтил уксусная кислота (1-НУК)
(2,4-D)
Эти синтетические регуляторы роста находят самое разнообразное применение:
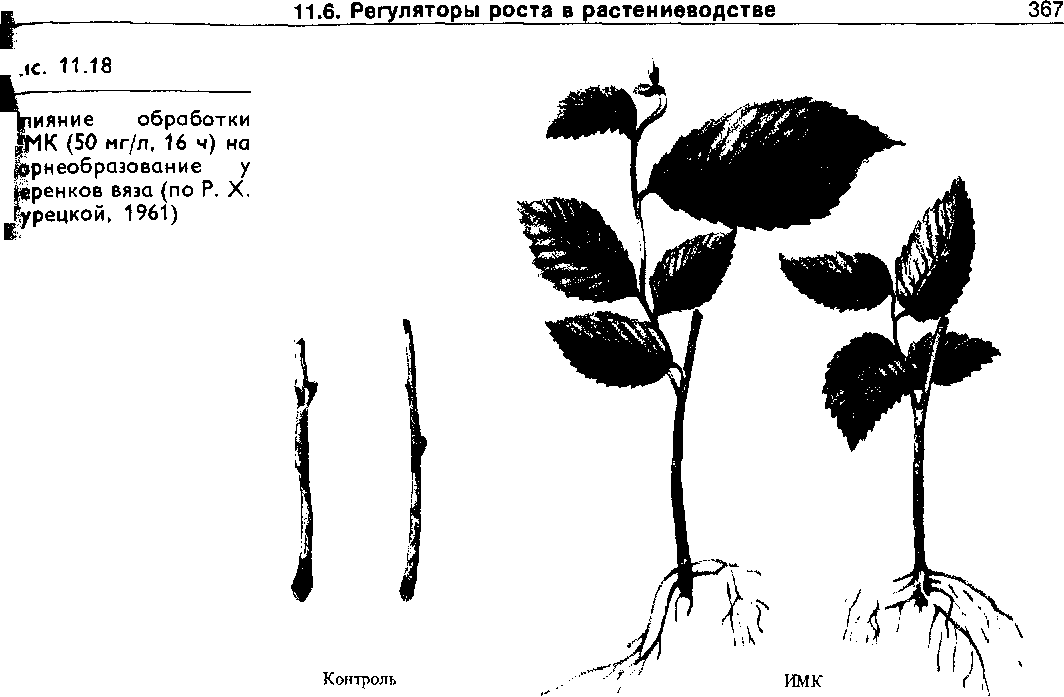
1. Стимуляция укоренения черенков. Для уко- ренения трудно приживляемых плодовых и лесных культур ос- нования их черенков обрабатывают раствором ИМК или 1-НУК (рис. 11.18).
Получение п а р т е н о к а р п и ч е с к и х (бессемянных) плодов и стимуляция плодообразо-в а н и я. Опрыскивание цветков томатов, огурцов и некоторых других культур растворами синтетических ауксинов индуцирует завязывание плодов без опыления. Этот прием широко используют в теплицах.
Уменьшение предуборочного опадения плодов. Обработка кроны яблонь, груш и др. 1-НУК или 2,4-Д задерживает образование отделительного слоя в плодоножках и существенно снижает потери урожая. Предуборочное опрыскивание замедляет также созревание плодов, что благоприятствует их дальнейшему хранению.
Прореживание цветков и завязей у плодовых. Для борьбы с периодичностью плодоношения бывает необходимо удалить излишнее количество образовавшихся цветков. Для этого раствором 1-НУК в повышенных концентрациях (15 — 50 мг/л) обрабатывают кроны деревьев во второй половине цветения. Опадение цветков связано с образованием этилена.
Уничтожение сорняков. 2,4-Д и другие хлорфе-ноксикислоты в дозах 0,6—1,5 кг/га широко используются для уничтожения широколиственных сорняков в посевах пшеницы, риса, кукурузы и других культур.
Рис.
11.19
Влияние
гиббереллина на урожай кишмиш-ного
сорта винограда (М.Х.Чайлахян,1963):
слева
соцветия
опрыскивались
раствором
ГА3,
справа
—
контрольные
растения
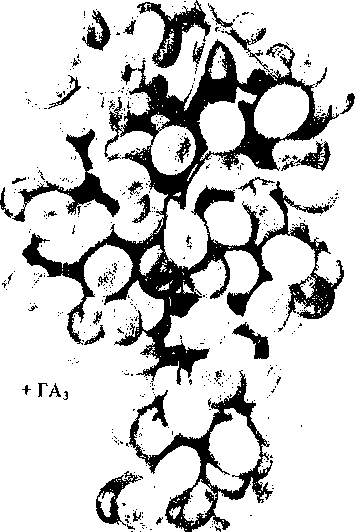
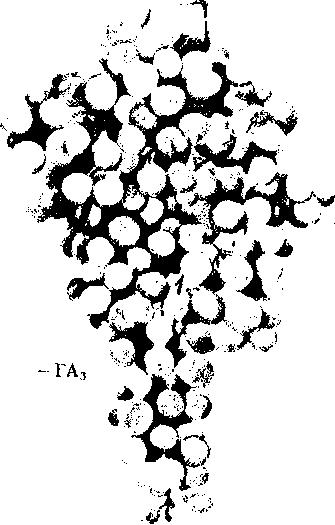
Повышение производства бессемянных сортов винограда. Ценные кишмишные сорта винограда имеют мелкие ягоды. Обработка гиббереллином способствует формированию более крупных кистей с ягодами большего размера (рис. 11.19).
Выведение из состояния покоя. Обработка свежеубранных клубней картофеля раствором гибберелловой кислоты (1—2 мг/л) и тиомочевины (20 мг/л) приводит к их быстрому прорастанию и увеличению количества проросших глазков. Этот прием используется в южных районах, где практикуются вторичные летние посадки картофеля.
Стимуляция образования солода. Гибберел-лин в зерновках ячменя активирует образование амилаз, что улучшает качество солода, используемого в производстве пива.
Ретарданты. Такое название получили синтетические вещества, тормозящие удлинение стебля. Механизм их действия обычно состоит в ингибировании синтеза гиббереллинов в растительных организмах. В качестве ретардантов используют хлорхолинхлорид (XXX, ТУР), алар и др.:
сн2С1-
сн3
\
+/снз сн2—с—nh—n;
сн,
сн2—соон
хлорхолинхлорид
алар
Наиболее широко ретарданты применяют для борьбы с полеганием хлебов, а также для торможения вытягивания рассады овощей, декоративных культур, роста кустарников.
Обработка аларом делает кроны плодовых более компактными и ускоряет переход молодых деревьев к плодоношению. Алар является также эффективным средством, предотвращающим предуборочное опадение плодов.
Этилен. Этилен используют для ускорения созревания зеленых плодов перед их продажей. Для других целей применяют производное этилена — этрел, молекулы которого устойчивы в кислом растворе, но распадаются, проникая в клетки в условиях слабощелочной среды; при этом освобождается этилен:
СЛСН,—СН2—Р—о- + он
II
о
этрел
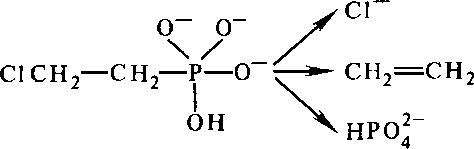
Этрел (или советский препарат гидрел) используется для стимуляции одновременного созревания плодов с последующей их машинной уборкой, а также для прореживания цветков и завязей. Опрыскивание растений огурцов, тыквы и др. раствором этрела приводит к образованию большого количества женских цветков и увеличению урожая плодов.
В ходе онтогенеза растительные организмы проходят ряд этапов: эмбриональный, ювенильный,узрелости и размножения, старости и отмирания. Каждый из этих этапов в свою очередь включает в себя несколько последовательных фаз роста и развития. Закладка органов происходит в апикальных меристемах, формирование тканей начинается с образования инициальных клеток. Особенности роста и морфогенеза обусловлены локальной скоростью и длительностью деления и растяжения клеток, а также векторностью этих процессов, которая определяется поляризацией клеток. Характер деления, растяжения и дифференцировки клеток зависит от взаимодействия клеток и от их местоположения. Высокая способность растений к регенерации определяется их прикрепленным образом жизни. Регенерация осуществляется на основе тех же механизмов, что и процессы морфогенеза при нормальном развитии. Рост растений отличается периодичностью. В неблагоприятные периоды растения переходят в состояние вынужденного или глубокого (физиологического) покоя.
Главными эндогенными факторами, влияющими на рост, морфогенез, регенерацию и развитие растений, являются фитогормоны. В связи с этим они находят все более широкое применение в растениеводстве.
I'. iа в а 12