

Синтез гормонов в тканях стеблей, корней, плодов нт. Д.
Синтез гормонов в хлоропластах
Гормоны, ингибиторы
Эпигенез
Транспорт ассимилятов
Фотосинтез
Согласно этой схеме, фитогормоны образуются в разных частях растений, в том числе в хлоропластах, и действуют на процессы фотосинтеза как дистанционно, так и непосредственно на уровне хлоропластов. Дистанционное действие осуществляется благодаря регулирующему влиянию фитогормонов на процессы роста и развития (эпигенез), на отложение веществ в запас, на транспорт ассимилятов, т. е. на формирование и активность аттрагирующих центров. С другой стороны фитогормоны оказывают прямое действие на функциональную активность хлоропластов через изменение состояния мембран, активность ферментов, генерацию трансмембранного потенциала. Доказана также роль фитогормонов, в частности цитокинина, в биогенезе хлоропластов, синтезе хлорофилла и ферментов цикла Кальвина.
Ряд фитогормонов (индолилуксусная кислота, гибберелли- ны, абсцизовая кислота), а также некоторые физиологически активные фенолы образуются в тканях листа. Цитокинин же, от которого во многом зависит формирование листа и его структур, поступает из других частей растения, прежде всего из корня. Такая система создает взаимозависимость всех органов, обеспечивая регуляцию функциональной активности в 6 целом растении.
яявя^яй!й!Я!1™,в! Под экологией фотосинтеза понимают зависимость продук- Экология тивности фотосинтеза от факторов внешней среды: интен-
фотосинтеза сивности и качества света, концентрации С02, температуры,
водного режима тканей листа, минерального питания и др.
Влияние этих факторов и адаптация к ним растений очень
существенны для растениеводства.
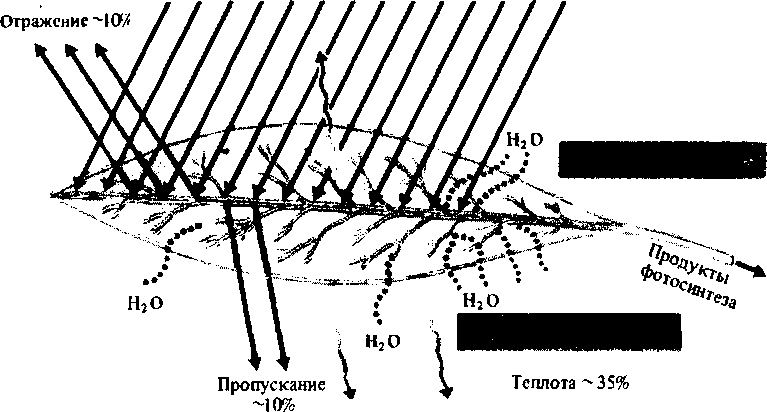
Интенсивность и спектральный состав света. В среднем листья поглощают 80 — 85% энергии фотосинтетически активных лучей солнечного спектра (400—700 нм) и 25% энергии инфракрасных лучей, что составляет около 55% от энергии общей радиации. На фотосинтез расходуется 1,5 — 2% поглощенной энергии (фотосинтетически активная радиация — ФАР) (рис. 3.21).
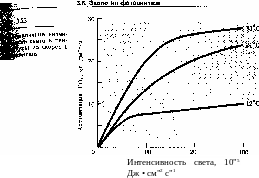
Зависимость скорости фотосинтеза от интенсивности света имеет форму логарифмической кривой (рис. 3.22). Прямая зависимость скорости процесса от притока энергии наблюдается только при низких интенсивностях света. Фотосинтез начинается при очень слабом освещении. Впервые это было показано А. С. Фаминцыным в 1880 г. на установке с искусственным освещением. Света керосиновой лампы оказалось достаточно для начала фотосинтеза и образования крахмала в растительных клетках. У многих светолюбивых растений максимальная (100%) интенсивность фотосинтеза наблюдается при освещенности, достигающей половины от полной солнечной, которая, таким образом, является насыщающей. Дальнейшее возрастание освещенности не увеличивает фотосинтез и затем снижает его.
Анализ световых кривых фотосинтеза позволяет получить информацию о характере работы фотохимических систем и ферментативного аппарата. Угол наклона кривой характеризует скорость фотохимических реакций и содержание хлорофилла: чем он больше, тем активнее используется световая энергия. Обычно больше он у теневыносливых растений, обитающих под пологом леса, и у глубоководных водорослей. У этих растений, приспособленных к условиям слабого освещения, хорошо развитый пигментный аппарат позволяет активнее использовать низкие интенсивности света.
Активность фотосинтеза в области насыщающей интенсивности света характеризует мощность систем поглощения и восстановления С02 и определяется концентрацией С02. Чем выше кривая в области насыщения интенсивности света, тем мощнее аппарат поглощения и восстановления С02. У светолюбивых растений насыщение достигается при значительно
Рис. 3.22
25
 Зависимость
скорости фотосинтеза от интенсивности
света у кукурузы
Зависимость
скорости фотосинтеза от интенсивности
света у кукурузы
большей освещенности, чем у теневыносливых. Например, у теневыносливого печеночного мха маршанции световое насыщение фотосинтеза достигается при 1000 лк, у светолюбивых древесных растений — при 10 — 40 тыс. лк, а у некоторых высокогорных растений Памира (где освещенность достигает максимальных на Земле значений порядка 180 тыс. лк)— при 60 тыс. лк и выше. Светолюбивы большинство сельскохозяйственных и древесных растений, а также водоросли мелководий.
У растений, осуществляющих С3путь фотосинтеза, насыщение происходит при более низкой интенсивности света, чем у растений с С4-путем превращения углерода, высокая фото-синтезирующая активность которых проявляется только при высоком уровне освещенности.
В области светового насыщения интенсивность фотосинтеза значительно выше интенсивности дыхания. При снижении освещенности до определенной величины интенсивности фотосинтеза и дыхания уравниваются. Уровень освещения, при котором поглощение ССь в процессе фотосинтеза уравновешивается выделением С02 в процессе дыхания, называется световым компенсационным пунктом. Его величину определяют при 0,03% С02 и температуре 20 °С. Значение светового компенсационного пункта неодинаково не только у теневыносливых (составляет примерно 1 % от полного света) и светолюбивых растений (около 3 — 5% от полного солнечного света), но и у листьев разных ярусов одного и того же растения, оно зависит также от концентрации ССь в воздухе. Чрезмерно высокое освещение резко нарушает процесс биосинтеза пигментов, фотосинтетические реакции и ростовые процессы. что в итоге снижает общую продуктивность растений.
Существенно, что даже кратковременное изменение условий освещенности влияет на интенсивность фотосинтеза. Это важ-
 ное
адаптационное свойство позволяет
растениям в фито-ценозах полнее
использовать свет. Фотосинтетический
аппарат «настраивается» на периодические
сдвиги освещенности при ветре, на
частоту мелькания бликов в доли секунды.
ное
адаптационное свойство позволяет
растениям в фито-ценозах полнее
использовать свет. Фотосинтетический
аппарат «настраивается» на периодические
сдвиги освещенности при ветре, на
частоту мелькания бликов в доли секунды.
На ход световых кривых фотосинтеза влияют изменения других факторов внешней среды. Например, при низких температурах (12 °С) повышение интенсивности света становится малоэффективным (рис. 3.23). Температурный оптимум у растений с С3-типом фотосинтеза лежит в пределах 25 — 35 °С. Повышение концентрации С02 с увеличением освещенности приводит к возрастанию скорости фотосинтеза (рис. 3.24).
Помимо интенсивности для процесса фотосинтеза важен и качественный состав света. Скорость фотосинтеза в разных участках спектра, выравненных по количеству энергии, не одна и та же. Наиболее высока интенсивность фотосинтеза в красных лучах.
Почему именно красные лучи наиболее эффективны для фотосинтеза? Во-первых, потому, что энергия 1 кванта красного света (176 кДж/моль = 42 ккал/моль) вполне достаточна для перехода молекулы хлорофилла на первый синглетный уровень возбуждения s* (см. рис. 3.4). Затем эта энергия целиком может быть использована на фотохимические реакции. Энергия же 1 кванта синего света выше (293 кДж/моль = = 70 ккал/моль). Поглотив квант синего света, молекула хлорофилла переходит на более высокий уровень синглетного возбуждения s*, и эта излишняя энергия превращается в теплоту при переходе молекулы в состояние s*. Энергия 1 кванта красного света примерно эквивалентна энергии перехода окислительно-восстановительного потенциала системы от е'0 = +0,8 В до е'0= -0,8 В (см. рис. 3.10). Энергия 1 кванта инфракрасных лучей уже недостаточна для фотоокисления воды, но у фотосинтезирующих серных бактерий эта энергия
>ис. 3.24
0,32%
С02
$заимовлияние
интен-:ивности света и концентрации
С02
на ско-эость фотосинтеза у
мха
(Б. А. Рубин, а> i.
Ф.
Гавриленко, 1977) я
0,16%
0,04%
<
0,01%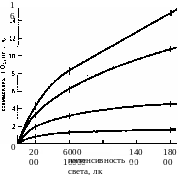
Во-вторых, красный свет всегда присутствует в лучах прямой солнечной радиации. Если солнце находится под углом 90°, то красные лучи составляют примерно '/4 часть полного солнечного света. Если же солнце стоит низко, красные лучи становятся преобладающими. При стоянии солнца под углом 5° красный свет составляет 2/3 от полного (рис. 3.25).
Растения, выращенные на синем и красном свету, существенно различаются по составу продуктов фотосинтеза. По данным Н. П. Воскресенской (1965), при выравнивании синего и красного света по квантам, т. е. при одинаковых для фотохимической стадии фотосинтеза условиях освещения, синий свет уже через несколько секунд экспозиции активирует включение 14С в неуглеводные продукты — амино- и органические кислоты, главным образом в аланин, аспартат, малат, цитрат, и в более поздние сроки (через минуты) — во фракцию белков, а красный свет при коротких экспозициях — во фракцию растворимых углеводов и при минутных экспозициях — в крахмал. Таким образом, на синем свету по сравнению с красным светом в листьях дополнительно образуются неуглеводные продукты. Эти различия в метаболизме углерода при действии света разного качества обнаружены у целых растений с С3- и С4-путями ассимиляции С02, у зеленых и красных водорослей; они сохраняются при различных концентрациях СОо и неодинаковой интенсивности света. Но у изолированных хлоропластов различий в образовании крахмала на синем и красном свету не обнаружено. Полагают, что фоторецептором, с деятельностью которого связаны изменения в метабо-