

30. Зависимость энергетического профиля ионного канала от заполнения канала, от биоэлектрических процессов. Селективность каналов.
Канал с одним участком связывания может находится в двух состояниях – в свободном и связанном. Канал с двумя участками связывания – в 4-х состояниях (рис.5.7).
Если один участок в канале заполнен, то поступление в свободный участок второго иона будет затруднено в связи с необходимостью затрат на преодоление энергии кулоновского взаимодействия с уже находящимся в канале ионом.
Попадание иона в левую потенциальную яму приведет к повышению энергетического барьера, соответствующего переходу иона из раствора в правую потенциальную яму (рис.5.7).
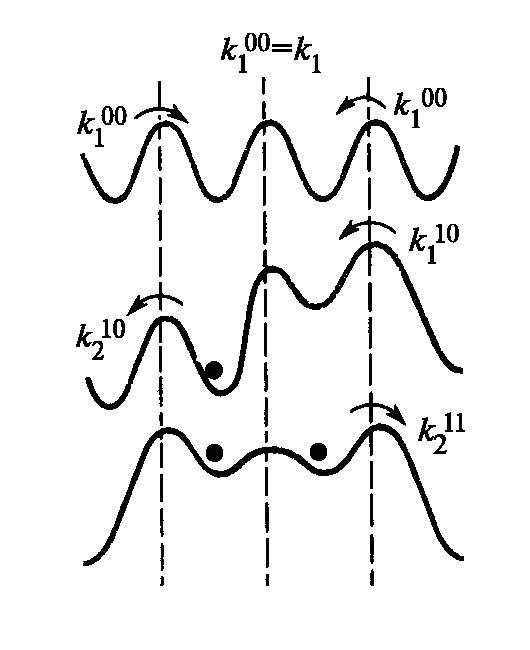
Рис. 5.7. Зависимость энергетического профиля от заполнения канала при учете кулоновских взаимодействий (Рубин, 2004)
k1 —константы поступления иона в раствор, k2 — константы выхода иона из раствора. Верхние индексы соответствуют разным состояниям канала с разным заполнением участков связывания
Вход второго иона в канал по сравнению с первым затрудняется, а выход его идет напротив легко. Т.е. кулоновское ион-ионное взаимодействие ускоряет общий ионный транспорт через канал.
Сильное электрическое поле иона вызывает поляризацию и переориентацию близлежащих полярных групп белка-переносчика. При этом смещается уровень потенциальной энергии в участке связывания и изменяется высота ближайших энергетических барьеров.
Особое важное значение могут иметь циклические изменения конформации мембранных белков, обусловленные биоэнергетическими процессами. Такие конформационные перестройки протекают в АТФазе энергосберегающих мембран, в цитохромах - при изменении окислительно-восстановительного потенциала, в бактериородопсине - при возбуждении хромофорной группы. По-видимому системы активного транспорта ионов, использующие энергию света, представляют трансмембранный ионный канал с регулируемой высотой энергетических барьеров.
Селективность каналов биомембран.
При проникновении иона в канал возрастание свободной энергии, обусловленное потерей гидратной оболочки (500-700 Дж/моль), компенсируется понижением энергии при взаимодействии с дипольными группами канала. Проникновение иона облегчает кулоновское взаимодействие катиона с фиксированным анионным центром в просвете канала.
Селективность зависит от радиуса анионного центра. Для больших анионных радиусов энергия кулоновского взаимодействия мала, определяющим фактором становится взаимодействие катиона с водой. В этом случае энергия возрастает с увеличением ионного радиуса: Cs>Rb>K>Na>Li.
Для малых анионных радиусов преобладает притяжение катиона, поэтому катионы меньшего радиуса связываются прочнее: Li>Na>K>Rb>Cs. Когда размер иона достигает критического значения, его проницаемость падает до нуля.
Для органических катионов (гидроксиламин, гидразин) также важна способность образовывать водородные связи.
На селективность канала влияет также химическое строение макромолекулы, а также движение сопряженных с ионов молекул воды.