

42.Действие ультрафиолетового излучения на белки, на биомембраны.
Действие УФ-света на белки.
Второй внутриклеточной мишенью при действии УФ-излучения на биосистемы являются белки. УФ-свет инактивирует белки по одноквантовому механизму, производя фотолиз цистина и триптофана. Аминокислоты переходят в синглет-возбужденное состояние и фотоионизируют с образованием катион-радикала и электрона (рис. ).
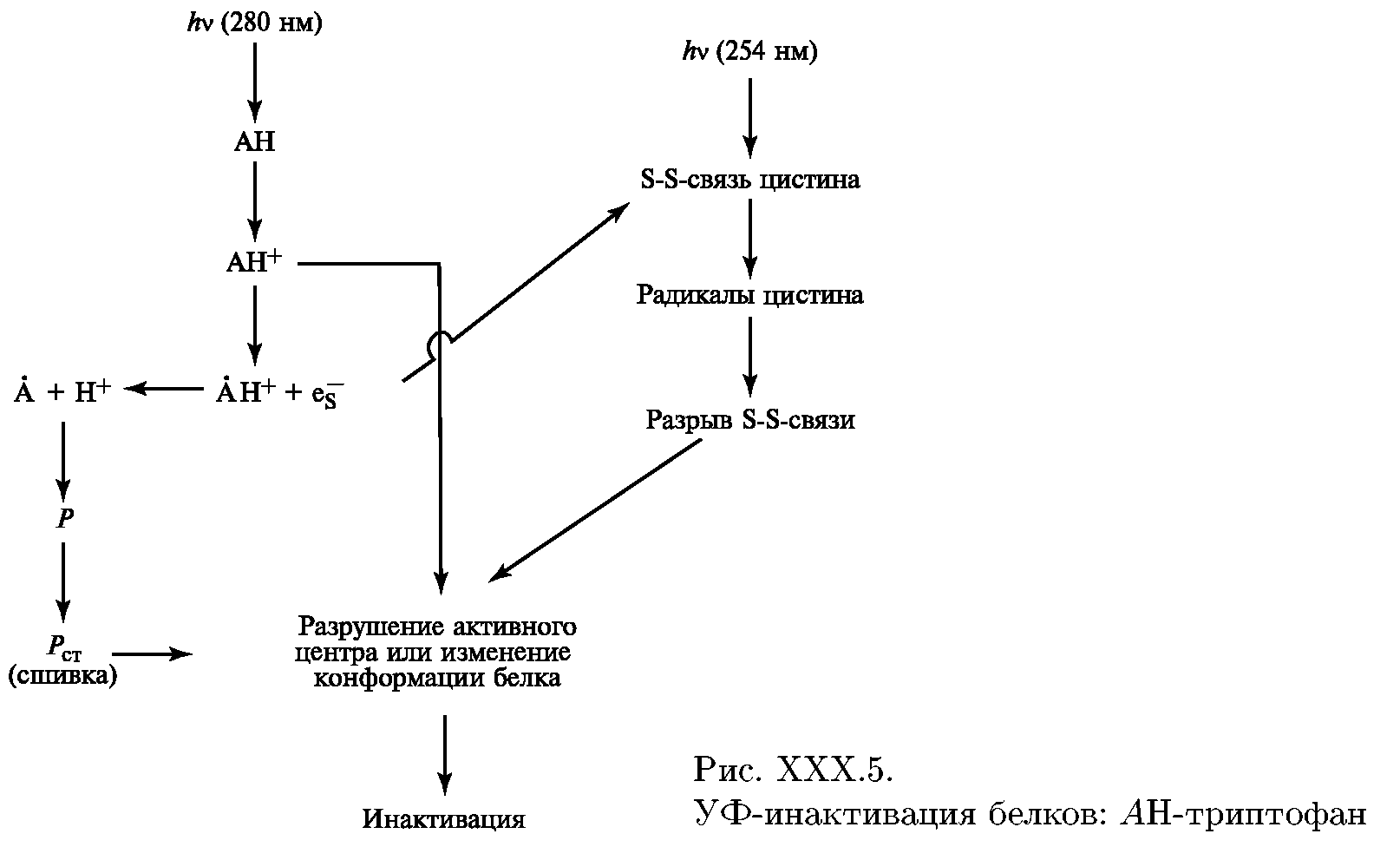
Рис. УФ-инактивация белков: АН-триптофан
Если фотолизу подвергается триптофан, входящий в состав активного центра фермента, то происходит потеря его ферментативной активности. Вне активного центра сшивка может изменить водородные, гидрофобные, и другие слабые связи, поддерживающие конформацию белковой макромолекулы, что ведет также к потере ее активности.
Сольватированный электрон реагирует с другими молекулами, образует активные формы кислорода, гидроперекисный радикал НО2. Кроме того, сольватированный электрон разрушает аминокислоты, особенное сродство он имеет к бисульфидным группам цистина.
Действие излучения на биомембраны. Оптическое излучение может вызвать увеличение проницаемости мембран, особенно в отношении ионов. УФ-излучение изменяет проницаемость мембран животных клеток из-за фотолиза липидов. Сенсибилизаторами, по-видимому, являются гидроперекиси, которые окисляют липиды с образованием гидроперекиси (цепная реакция). Нарушаются свойства липидной части мембраны и разрушаются белковые SH-группы, необходимые для барьерных свойств.
Мембраны растительных клеток имеют более высокую чувствительность к УФ-излучению: деполяризация и увеличение проницаемости. Наиболее чувствительный компонент – мембранная АТФаза. Фотоинактивация происходит при прямом поглощении квантов УФ-излучения, хромофором служит триптофан.