

27. Азотистые основания, входящие в структуру нуклеиновых кислот – пуриновые и пиримидиновые. Нуклеотиды, содержащие рибозу и дезоксирибозу. Структура. Номенклатура.
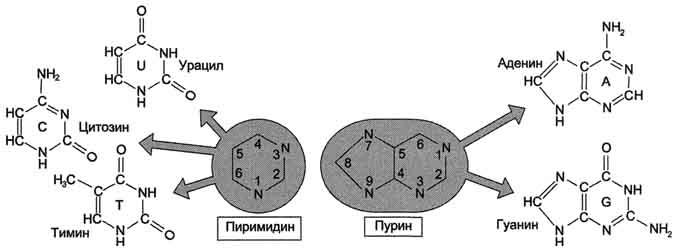
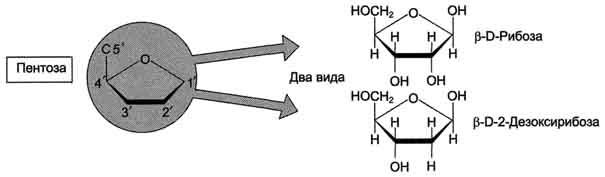
27. Первичная структура нуклеиновых кислот. ДНК и РНК–черты сходства и различия состава, локализации в клетке, функции. Вторичная структура ДНК (модель Уотсона и Крика). Связи, стабилизирующие вторичную структуру ДНК. Комплементарность. Правило Чаргаффа. Полярность. Антипараллельность.
Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинукпеотидной цепи.
Каждая фосфатная группа в полинукпеотидной цепи, за исключением фосфорного остатка на 5'-конце молекулы, участвует в образовании двух эфирных связей с участием 3'- и 5'-углеродных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3', 5'-фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращённо записывают с помощью однобуквенного кода, например -A-G-C-T-T-A-C-A- от 5'- к 3'-концу.
В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При рН 7 фосфатная группа полностью ионизирована, поэтому in vivo нуклеиновые кислоты существуют в виде полианионов (имеют множественный отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной
Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце - ОН-группа 3'-углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК. Гидроксильная группа у 2'-углеродного атома рибозы делает молекулу РНК нестабильной. Так, в слабощелочной среде молекулы РНК гидролизуются даже при нормальной температуре, тогда как структура цепи ДНК не изменяется.
|
Признаки |
ДНК |
РНК |
|
Местонахождение в клетке |
Ядро, митохондрии, хлоропласты |
Ядро, рибосомы, цитоплазмы, митохондрии, хролопласты |
|
Местонахождение в ядре |
Хромосомы |
Ядрышко |
|
Строение макромолекулы |
Двойной неразветвленный линейный полимер, свернутый правозакрученной спиралью |
Одинарная полинуклеотидная цепочка |
|
Мономеры |
Дезоксирибонуклеотиды |
Рибонуклеотиды |
|
Состав нуклеотид а |
Азонистое основание (пуриновое-аденин, гуанин, пиримидиновое – тимин, цитозин); дезоксирибоза (углевод); остаток фосфорной кислоты |
Азонистое основание (пуриновое-аденин, гуанин, пиримидиновое-урацил, цитозин);рибоза (углевод); остаток фосфорной кислоты |
|
Типы нуклеидов |
Адениловый (А), гуаниловый(Г), тимидиловый (Т), цитидиловый (Ц) |
Адениловый (А), гуаниловый (Г), уридиловый (Т),цитидиловый (Ц) |
|
Свойства |
Способная к самоудвоению по принципу комплементарности А=Т, Т=А, Г=Ц, Ц=Г Стабильна. |
Не способна к самоудвоению. Лабильна. |
|
Функции |
Химическая основа хромосомного генетического материала (гена); синтез ДНК, синтез РНК, информация о структуре белков. |
Информационная (иРНК) – передает код наследственной информации о первичной структуре белковой молекулы, рибосомальная (рРНК) – входит в состав рибосом; транспортная (тРНК) – переносит аминокислоты к рибосомам; митохондриальная и платидная РНК – входят в состав рибосом этих органелл |
Вторичная структура ДНК. В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи). При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль.
Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки - большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.