

TOM-3
.pdfции кар тина позитивно изменялась – в значительной степени (до 82%) возрастало число остр овков с интенсивной и умер енно выр аженной степенями гр ануляции. В ацинар ной ткани отмечается появление единичных клеток (ацино-инсуляр ных), обладающих базофильными свойствами. Пр и этом фактически не обнар уживались (0,2%) остр овки без гр ануляции. Вид остр овков из пр епар атов поджелудочной железы интактных, контр ольных и опытных гр упп животных иллюстр ир уется р ис. 2.
Биохимические наблюдения. У животных контр ольной гр уппы на 3-й день после введения аллоксана ур овень глюкозы повышался с 3,6 ± 0,014 ммоль/л до 17,8 ± 0,084 ммоль/л. Пр и этом ур овень инсулина падал с 0,30 ± 0,02 мкг/л до 0,14 ± 0,016 мкг/л. В последующем на 7-й и 15-й дни ур овень глюкозы несколько снижался (соответственно 14,6 ± 1,25 и 12,2 ± 0,71 ммоль/л), а ур овень инсулина медленно повышался (соответственно 0,16 ± 0,014 и 0,18 ± 0,02 мкг/л).
У животных опытной гр уппы исходные ур овни глюкозы и инсулина, не отличались от контр оля (соответственно 3,7 ± 0,09 ммоль/л и 0,36 ± 0,03 мкг/л). Как и пр едполагалось, не было обнар ужено существенных отличий по глюкозе и инсулину от контр оля и на 3-й день наблюдения. Основные статистически достовер ные отличия (p < 0,05) от контр оля отмечены на 7-й и 15-й дни наблюдения. Ур овень глюкозы на 7-й день падал до 7,2 ± 0,39 ммоль/л, а на 15-й день составлял 4,0 ± 0,16 ммоль/л, что почти соответствует исходному ур овню. В то же вр емя ур овень инсулина отчетливо повышался, достигая на
7-й день 0,25 ± 0,03 мкг/л, а на 15-й – 0,27 ± 0,02 мкг/л. Оба значения статистически достовер но отличаются от контр оля (p < 0,05), хотя ур о- вень инсулина на 15-день и не достиг исходного значения. Это вполне соответствует нашим мор фологическим наблюдениям, пр едставленным на таблице 2, где показано, что число остр овков с интенсивно гр анулир ованными инсулинсулинпр одуцир ующими клетками меньше исходного.
Результаты 3-й сер ии опытов пр едставлены на р ис. 3, где сопоставлена динамика ур овней глюкозы и инсулина в кр ови у животных опытной и контр ольной гр упп. Следует отметить, что значения глюкозы и инсулина высоко достовер но отр ицательно коррр елир уют и в контр оль-
ной (r = 0,987), и опытной (r = 0,912) гр уппах.
Обсуждение результатов
Пр едставленные данные показывают, что антигипер гликемическое действие тр анскр аниальной электр остимуляции пр и аллоксановом повр еждении β-клеток может осуществляться двумя путями.
Во-пер вых, электр овоздействие существенно ускор яет восстановление инсулинпр одуцир ующей функции повр ежденных β-клеток, ко-
141
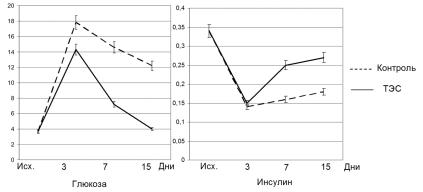
Рис. 3. Динамика изменений ур овней глюкозы и инсулина в кр ови у кр ыс конт- р ольной и опытной гр упп.
Обозначения: Ур овни глюкозы и инсулина в контр оле – пр ер ывистые линии, в опыте – сплошные линии. По оси ор динат – концентр ации глюкозы (в ммоль/л) и инсулина (в мкг/л), по оси абсцисс – дни наблюдений.
тор ые после однокр атного введения аллоксана могут сами постепенно восстанавливаться, но в гор аздо более поздние ср оки [3]. Эффект тр анскр аниальной электр остимуляции подтвер ждается р езультатами мор фологических наблюдений – восстановлением стр уктур ы остр овков, появлением гр анул инсулина в дегр анулир ованных под действием аллоксана β-клетках и увеличением ур овня инсулина в кр ови по ср авнению с контр олем в ср оки, не сопоставимые с контр олем.
Во-втор ых, весьма вер оятно, что электр овоздействие стимулир у- ет также появление новых β-клеток. Известно, что β-клетки обладают в опр еделённой мер е способностью р егенер ир овать как за счет митотического деления, так и путем тр ансфор мации ацинар ных или дуктальных панкр еатических клеток в инсулиноциты [9, 10, 27]. Пр и этом могут фор мир оваться остр овки малой площади, как это и показано в настоящей р аботе. Однако, какой из этих путей возникновения β-кле- ток имеет место или все они одновр еменно активир уются тр анскр аниальной электр остимуляцией, в настоящее вр емя не изучено и может явиться пр едметом дальнейших исследований.
Важно подчер кнуть, что указанные выше выр аженные эффекты тр анскр аниальной электр остимуляции пр оявляются после всего тр ех ежедневных сеансов, начатых в день введения аллоксана, т.е. элект- р овоздействие имеет длительное последействие. Аналогичное последействие отмечено нами и в отношении восстановления функции и р егенер ации токсически повр ежденных гепатоцитов [8].
Известно, что тр анскр аниальной электр остимуляции в пр именяемом р ежиме активир ует выделение β-эндофина [1], а ее антигипер -
142
гликемический эффект не пр оявлялся на фоне введения налоксона – блокатор а опиоидных р ецептор ов [7]. Поскольку β-клетки обладают опиоидными р ецептор ами [30], пр едставлялось естественным пр едположить, что наблюдаемые эффекты обусловлены непоср едственной их эндор финер гической активацией.
Действительно показано, что хотя пр и отсутствии повр еждений β- клеток введение β-эндофина тор мозит выр аботку инсулина и способ ствует р азвитию гипер гликемии [18], то на фоне повр еждения β-клеток отмечен обр атный эффект. Интер есно ср авнить р езультаты двух исследований, в котор ых пр и остр ом стр ессе и сохр анных β-клетках β-эндо- фин вызывал р азвитие гипер гликемии и снижение ур овня инсулина в кр ови [19], а пр и экспер иментальном диабете в условиях стр есса вызывал гипогликемический эффект, чего не наблюдалось у интактных особей [23]. Отмечены и др угие особенности функционир ования эндор финер гических механизмов на фоне повр еждения β-клеток. Так, на этом фоне снижается пр одукция β-эндофина в гипофизе [14], и только у животных с экспер иментальным диабетом некотор ые мор финоподобные вещества способны повышать ур овень β-эндофина кр ови [21]. Интер есно, что у гер оиновых нар команов также отмечен повышенный базовый ур овень инсулина [25].
Молекуляр ный механизм действия β-эндофина на повр ежденные инсулиноциты не изучен хотя бы по той пр ичине, что механизм пов- р еждения этих клеток аллоксаном или стр ептозотоцином не вполне ясен. Пр едполагается, что пер вичной мишенью аллоксана в β-клетках являются те элементы, котор ые участвуют в стимулир ующем влиянии глюкозы на пр одукцию инсулина – глюкозный тр анспор тер -2 (GLUT-2) и глюкокиназа, а для стр ептозотоцина – в основном GLUT- 2 [28]. Поскольку антигипер гликемический эффект тр анскр аниальной электр остимуляции пр оявляется не только пр и аллоксановом [6], но и пр и стр ептозотоциновом диабете [наши неопубликованные данные], можно пр едполагать, что β-эндофин может активир овать GLUT-2 [16].
Влияние β-эндофина на гомеостатическую р егуляцию ур овня глюкозы не огр аничивается, вер оятно, только р епар ацией повр ежденных β-клеток. Имеются данные о том, что β-эндофин может усиливать пот- р ебление глюкозы мышцами с помощью механизма, независимого от инсулина [17]. Вер оятно поэтому метфор мин, котор ый не влияет на пр одукцию инсулина, но стимулир ует инсулинзависимое поступление глюкозы в ткани и тор можение глюконеогенеза в печени [22], сущест венно усиливает антигипер гликемический эффект тр анскр аниальной электр остимуляции [7]. В то же вр емя данные о влиянии β-эндофина на глюконеогенез достаточно пр отивор ечивы, и, следовательно, этот эффект скор ее не может оказывать влияния на ур овень глюкозы кр ови
[13, 24].
143
Таким обр азом, полученные данные показывают, тр анскр аниальная электр остимуляция эндор финных стр уктур мозга может р ассмат- р иваться как пер спективное немедикаментозное неинвазивное воздействие для лечения диабета у человека, тр ебующее дальнейшего исследования. Не следует, конечно, напр ямую пер еносить данные, полученные пр и экспер иментальном аллоксановом и стр ептозотоциновом диабете у кр ыс и мышей, на патологию человека, поскольку имеются отчетливые межвидовые отличия – β-клетки человека, напр имер, гор аздо менее чувствительны к диабетогенным веществам, пр именяемым в экспер иментах [15].
Тем не менее, можно наметить некотор ые напр авления изучения этого вопр оса в клинической пр актике, поскольку пр облема лечения диабета весьма актуальна. Возможно, что пр именение электр овоздействия может быть эффективным пр и диабете II типа в ситуации, когда пер ор альные сахар оснижающие пр епар аты уже не могут устр а- нить гипер гликемию, поскольку инсулинпр одуцир ующая функция β- клеток значительно снижена, и пр иходится начинать введение малых доз инсулина. Не исключено пр именение этого метода и в некотор ых случаях диабета I типа, поскольку в биоптатах поджелудочной железы значительной части таких больных выявлены р асположенные в ацинусах отдельные β-клетки с высокой степенью секр етор ной активности и без пр изнаков дегенер ации, пр ичем наличие их не коррр елир овало ни с полом, ни с возр астом, ни ср оками манифестации и длительностью заболевания [11]. Именно они могут стать источником р епар ативной р егенер ации остр овков.
В последнее вр емя интенсивно изучается возможность лечения диабета с помощью имплантации β-клеток, котор ые, однако, быстр о становятся нежизнеспособными [6]. Не исключено, что и в этом случае пр именение тр анскр аниальной электр остимуляции может быть полезным не только для поддер жания их функции, но и их мультипликации.
Заключение
Установлено, что тр анскр аниальная электр остимуляция эндор- финер гических стр уктур мозга у кр ыс с аллоксановым диабетом спо собствует р епар ативной р егенер ации β-клеток остр овков Лангер ганса поджелудочной железы и восстанавливает их инсулинпр одуцир у- ющую функцию. Рассматр иваются возможные механизмы наблюдаемого эффекта и пр едполагаемые напр авления его лечебного пр именения.
144
Литература
1. Айрапетов Л. Н., Зайчик А. М., Трухманов М. С., Лебедев В. П. и др.
Об изменении ур овня β-эндор фина в мозге и спинномозговой жидкости пр и тр анскр аниальнойэлектр оанальгезии//Физиол.жур н.СССРим.И.М.Сеченова. –
1985.–Т. 71, № 1. – С. 56–63.
2.АлександроваВ.А.,РычковаС.В.,ЛебедевВ.П.идр.Влияние тр анскр а- ниальной электр остимуляции опиоидных стр уктур головного мозга на р егене- р ацию язвенных дефектов слизистой оболочки желудка и двенадцатипер стной кишки. // Междунар од. мед. обзор ы. – 1994. – Т. 1 – С. 41–46.
3.Алиева Г. Н., Киясов А. П., Миннебаев М. М., Бурыкин И. М., Хафизь-
янова Р. Х. Динамика β- и β-клеточных популяций поджелудочной железы
исодер жания глюкозы в кр ови кр ыс пр и аллоксановом диабете // Бюл. эксп. биол. и мед. – 2002. – Т. 133, № 2. – С. 151–153.
4.Баранов В. Г., Соколоверова Н. М., Гаспарян Э. Г., Ярошевский Ю. А.
Экспер иментальный сахар ный диабет / Л.: «Наука». 1983.
5.Дорше Х. Х. фон, Фалкмер С. Онтогенез остр овков Лангер ганса человека. Обзор некотор ых данных световой и электр онной микр оскопии, иммуногистохимических и функциональных данных о фетальном р азвитии эндокр инной поджелудочной железы // Жур н. эволюц. биохим. физиол. – 2000. – Т. 36, № 6 – С. 527–552.
6.Кудряшев Б. А., Ульянов А. М. Стабильная р едукция пр одукции инсулина в ор ганизме пр и экспер иментальном инсулин-зависимом диабете // Вопр . мед. хим. – 1998. – Т. 37, № 4. – С. 40–43.
7.Лебедев В. П., Биличенко С. В., Малыгин А. В., Нечипоренко С. П., Кол-
басов С. Е., Мелихова М. В. Тр анскр аниальная электр остимуляция нор мализует ур овень сахар а кр ови пр и аллоксановом диабете у кр ыс // Рос. физиол. жур н. им. И. М. Сеченова. – 2004. – Т. 90, № 11. – С. 1426–1429.
8.Лебедев В. П., Мелихова М. В., Колбасов С. Е., Стройкова Г. С., Замуру-
ев О. Н. Влияние тр анскр аниальной электр остимуляции эндор финных стр уктур мозга кр ыс на функциональную активность гепатоцитов пр и их токсическом повр еждении // Рос. физиол. жур н. им. И. М. Сеченова. – 2000. – Т. 86, № 11. – С. 1449–1457.
9.Пузырев А. А. Обр азование эндокр инных клеток поджелудочной железы человека из эпителия пр отоков и ацинусов // Ар х. анат., гист. и эмбр . – 1979. – Т. 76, № 1. – С. 20–25.
10.Пузырев А. А. Диффер енцир овка эндокр инных клеток поджелудочной железы белой кр ысы в составе эпителия выводных пр отоков. Ар х. анат., гист.
иэмбр . – 1982. – Т. 82, № 3. – С. 83–90.
11.Севергина Э. С., Дюжева Т. Г., Разгулина Л. Е., Стахеев И. Б. Локализация β-клеток в ацинусах – обычное явление или пр изнак компенсатор ного пр оцесса пр и инcулин-зависимом сахар ном диабете // Ар х. пат. – 1992. – Т. 52, № 12. – С. 18–23.
12.Тиц Н. У. Энциклопедия клинических и лабор атор ных тестов / М. «Лабинфор м». – 1997.
13.Brubaker P. L., Sun A., Vranic M. Lack of effect of beta-endorphin on basal or glucagon-stimulated hepatic glucose production in vitro // Metabolism. – 1987. – Vol. 36, № 5. – P. 432–437.
14.Cheyng C. Y., Tang F. The effect of streptozotocin-diabetes on betaendorphin le-vel and proopiomelanocortin gene expression in the rat pituitary // Neurosci. Lett. – 1999. Vol. 26, № 1–2 – P. 118–120.
145
10. Заказ № 282
15.Eizirik D. L., Pipeleers D. G., Ling Z., Welsh N. et al. Major species differences between humans and rodents in the susceptibility to pancreatic beta-cell injury // Proc. Natl. Acad. Sci. USA. – 1994. – Vol. 91, № 20. – P. 9253–9256.
16.Elsner M., Tiedge M., Guldbakke B., Munday R., Lenzen S. Importance of the GLUT2 glucose transporter for pancreatic beta cell toxicity of alloxan // Diabetologia. – 2002. – Vol. 45, № 11. – P. 1542–1549.
17.Evans A. A., Khan S., Smith M. E. Evidence for a hormonal action of betaendorphin to increase glucose uptake in resting and contracting skeletal muscle // J. Endocrinol. – 1997. – Vol. 155, № 2. – P. 387–392.
18.Fatouros I. G., Goldfarb A. H., Jamurtas A. Z., Angelopoulos T. J., Gao J.
Beta-endorphin infusion alters pancreatic hormone and glucose levels during exercise in rats // Eur. J. Appl. Physiol. Occup Physiol. – 1997. – Vol. 76, № 3. – P. 203–208.
19.Gunoz H., Dindar A., Neyzi O. beta-Endorphin and some hormonal levels in children with acute stress hyperglycaemia // Diabetes Res. Clin. Pract. – 1994. – Vol. 24, № 2. – P. 97–101.
20.Izumida Y., Aoki T., Yasuda D., Koizumi T., Suganuma C. et al. Hepatocyte growth factor is constitutively produced by donor-derived bone marrow cells and promotes regeneration of pancreatic beta-cells. // Biochem Biophys Res Commun.– 2005. – Vol. 333, № 1. – P. 273–282.
21.Kamei J. Antinociceptive effects of the enantiomorphs and its main metabolite in streptozotocin-induced diabetic mice // Nihon Shinkei Seishin Yakurigaku Zasshi.– 2000. – Vol. 20, № 1. – P. 11–16.
22.Kirpicnikov D., McFarlane S. I., Sowers J. R. Metformin: an update // Ann. Intern. Med. 2002. – Vol. 137, № 1. – P. 25–33.
23.Liu I. M., Niu C. S., Chi T. C., Kuo D. H., Cheng J. T. Investigations of the mechanism of the reduction of plasma glucose by cold-stress in streptozotocininduced diabetic rats // Neuroscience. – 1999. – Vol. 92, № 3. – P. 1137–1142.
24.Matsumura M., Fukushima T., Saito H., Saito S. In vivo and in vitro effects of beta-endorphin on glucose metabolism in the rat // Horm. Metab. Res. – 1984. – Vol. 16, № 1. – P. 27–31.
25.Passariello N., Giugliano D., Quatraro A., Consoli G., Sgambato S., Torella R., D’Onofrio F. Glucose tolerance and hormonal responses in heroin addicts. A possible role for endogenous opiates in the pathogenesis of non-insulin- diabetes dependent Metabolism. – 1983. – Vol. 32, № 12. – P. 1163–1165.
26.Slack J. M. Developmental biology of pancreas // Development. – 1995 .– Vol. 121, № 6. – P. 1569-1580.
27.Waguri M., Yamamoto K., Miygawa J. Y., Tochino Y., Yamamori K., et al.
Demonstration of two different processes of beta-cell regeneration in a new diabetic mouse model induced by selective perfusion of alloxan // Diabetes. – 1997. – Vol. 6, № 8. – P. 1281–1290.
28.Walde S. S., Dohle C., Schott-Ohly P., Gleichmann H. Molecular target structures in aloxaninduced diabetes in mice // Lfe Sci. – 2002. – Vol. 71, № 14. – P. 1681–1694.
29.WangR.,YashpalN.,BacchusF.,LiJ.J.Hepatocyte growth factor regulates proliferation and differentiation of epithelial monolayers derived from islets of postnatal rat pancreas // Endocrinol. – 2004. – Vol. 183, № 1. – P. 163–171.
30.Zhang M., Zheng M., Schleicher R. L. Autoradiographic localization of beta-endorphin binding in the pancreas // Mol. Cell Neurosci. – 1994. – Vol. 5, № 6. – P. 684–690.
19. Эффективность транскраниальной электростимуляции эндорфинергических структур мозга (ТЭС-терапии)
вкоррекции нарушений углеводного обмена
убольных сахарным диабетом типа 2
Рогова Н. В., Петров В. И.
Волгогр адский государ ственный медицинский универ ситет, Волгогр ад
Резюме
Оценивали эффективность ТЭС-тер апии пр и включении её в комплексное лечение сахар ного диабета тип 2 (СД2) у пациентов без ожир ения (ПбО) и пациентов с ожир ением (ПсО) по влиянию на ур о- вень гликемии и выр аботке инсулина. ТЭС-тер апия оказалась весьма эффективной у гр уппы ПбО, у котор ых имело место снижение функциональной активности β-клеток. Эффект ТЭС-тер апии пр евосходил по важным показателям эффекты глибенкламида (по нор мализации фаз секр еции инсулина, отсутствию гипогликемических состояний). У гр уппы ПсО, несмотр я на выр аженную гипер инсулинемию и инсулинор езистентность, ТЭС-тер апия не вызывала р оста гипер инсулинемии и снижала постпр андиальную гликемию, что свидетельствует, возможно, о способности ТЭС-тер апии снижать инсулинор езистентность. Общий эффект ТЭС-тер апии в моновар ианте был менее выр ажен по ср авнению с сиофор ом. Но комбинация ТЭС-тер апия с сиофор ом обеспечивала полную компенсацию нар ушений углеводного обмена у ПсО с СД2. Полученные данные показывают, что ТЭС-тер апия в комплексном лечении больных СД2 имеет опр еделенные пер спективы шир окого клинического пр именения пр и этой патологии.
Введение
Сахар ный диабет типа 2 (СД 2) пр едставляет собой серь езную медико-социальную пр облему, что обусловлено его высокой р аспр о- стр аненностью, тенденцией к р осту числа больных, хр оническим течением, опр еделяющим кумулятивный хар актер заболевания, высокую инвалидизацию [1]. Экспер тная оценка р аспр остр аненности сахар ного диабета позволяет считать, что к 2010 году количество больных сахар - ным диабетом будет увеличиваться и достигнет более чем 230 млн, из котор ых 85–90 % больных будут составлять больные диабетом типа 2 [2].
Пр ичинами р азвития самого СД 2 является инсулинор езистентность и нар ушение секр еции инсулина, фор мир ование котор ых обус-
147
ловлено комбинацией вр ожденных и пр иобр етенных фактор ов. Уст- р анение инсулинор езистентности (ИР) и сохр анение функциональной активности β-клеток поджелудочной железы являются важными аспектами пр еодоления системного дисметаболизма.
Пр облема комплексной тер апии СД 2 актуальна, так как для достижения компенсации углеводного обмена тр удно обойтись монотер апией, и лечение таких больных сопр овождается назначением большого количества лекар ственных ср едств [3].
В настоящее вр емя р азр абатываются комбинир ованные пр епар а- ты и начинается поиск новых нелекар ственных методов лечения СД. Показано, напр имер , что тр анскр аниальная электр остимуляция (ТЭС) защитных (эндор финных) механизмов мозга может быть пер спективным методом для лечения СД [4, 5]. Действительно, в экспер иментах на моделях с аллоксановым и стр ептозотоциновом диабетом было пр о- демонстр ир овано, что ТЭС обладает выр аженным антигипер гликемическим эффектом, способствует восстановлению стр уктур ы повр ежденных β-клеток и их способности выр абатывать инсулин. Кр оме того, установлено, что ТЭС эффективно потенцир овал антигипер гликемические эффекты сахар оснижающих пр епар атов.
Этиданныепозволяютпр едположить,чтовключениеТЭС-тер апии в пр актику лечения СД 2 позволит повысить эффективность лечения и уменьшить объем пр именения медикаментов. Конкр етная цель настоящей р аботы – изучить и оценить влияние ТЭС-тер апии на ур овень инсулина в плазме кр ови у больных СД 2 для последующей оптимизации фар макотер апии этой гр уппы пациентов.
Методы и материалы
По дизайну исследование откр ытое, пр остое, р андомизир ованное. В исследование включены мужчины и женщины от 18 до 70 лет. Состав гр упп по полу, возр асту и тяжести заболевания существенно не р азличается. Диагноз СД устанавливался на основании кр итер иев ВОЗ (1999). Все пациенты к моменту включения в исследование находились в состоянии субкомпенсации или декомпенсации углеводного обмена.
Контр ольную гр уппу (гр уппа 1) составили пациенты с гипер тонической болезнью I степени без нар ушений углеводного обмена и без ожир ения.
Всем пациентам с СД 2 после включения в исследование на 14 дней отменяли гипотензивные ср едства, а с 7-го дня этой отмены снимали сахар оснижающие ср едства (на этот пер иод пациенты госпитализир о- вались в стационар , т. е. с 7-го по 14-й день отмены пр епар атов гипотензивных).
После «отмывочного пер иода» все пациенты с СД 2 р азделялись по ИМТ на 2 гр уппы (без ожир ения – ИМТ < 25 кг/м2 и с ожир ением
148
ИМТ >25 кг/м2), затем в каждой из этих двух гр упп пр оводилась р андомизация пациентов. В гр уппе СД без ожир ения на две гр уппы (2-я и 3-я гр уппы, 2-я получала кур с ТЭС-тер апии; 3-я – микр онизир ованный глибенкламид (1,75 мг 2 р аза в день). В гр уппе СД 2 с ожир ением пр о- водилась р андомизация на 3 гр уппы (4, 5, 6). 4-я гр уппа получала кур с ТЭС-тер апии; 5-я гр уппа – сиофор (850 мг, 2 р аз в день); 6-я гр уппа – ТЭС ± сиофор (850 мг, 1 р аз в день). Далее всем выполнялось обследование и назначался 14-дневный кур с тер апии, затем пр оводилось контр ольное обследование – после лечения. ТЭС-тер апию осуществляли аппар атом ТРАНСАИР по общепр инятой схеме.
4–5-ти больным из каждой из 6-ти гр упп до и после лечения исследовали ур овень инсулина кр ови и глюкозы капилляр ной кр ови по 12 точкам. Вр емя забор а кр ови отмечено в таблице 1. Вр емя пр иема пищи стр ого устанавливалось: ЗАВТРАК – 7.45, ОБЕД – 11.45, УЖИН – 17.45. Остальным больным (100 человек, т. е. каждому из 16–20 человек в гр уппе) опр еделяли до и после лечения инсулин и глюкозу капилляр ной кр ови в 6.00 (натощак) и в 10.00 (постпр андиально).
Ур овень инсулина в сывор отке кр ови опр еделяли иммунологическим методом с помощью набор а пр оизводства фир мы «DRGDiagnostics» (Гер мания). Ур овень глюкозы опр еделяли в цельной капилляр ной кр ови из пальца с помощью пор тативного глюкометр а пр оизводства фир мы «Roche» (Гер мания).
Результаты исследования и их обсуждение
Результаты исследования пр иведены в таблицах № 1 и 2. Из полученных р езультатов исходно у пациентов с СД 2 имеются серь езные нар ушения в сточном пр офиле инсулинемии.
Кур с ТЭС-тер апии повышает базальную секр ецию инсулина во всех гр уппах без ожир ения и у здор овых (т. е. больных ГБ без нар у- шений углеводного обмена), смещает пик постпр андиальной секр еции ближе к р аннему пику (чер ез 15 мин. после еды небольшое увеличение, а чер ез 1 час после еды – пик), увеличивает величину пика постпр андиального инсулина. В этих гр уппах данные изменения сопр овождаются снижением ур овня глюкозы капилляр ной кр ови (пр ичем пик гликемии совпадает с пиком инсулинемии).
В гр уппе 3 (манинил) базальный ур овень инсулина повышается, но меньше, чем в гр уппе ТЭС, пик постпр андиальной секр еции остается почти таким же, как и без лечения, т. е. чер ез 2 часа после еды (без лечения – на 2,5 часа), однако амплитуда пика немного выше, чем пр и ТЭС, и шир ина пика больше, чем пр и ТЭС, что сопр овождается эпизодами гипогликемии (особенно ночью).
Кур с ТЭС-тер апии в гр уппах с ожир ением не изменяет ур овень базальной секр еции, смещает пик постпр андиальной секр еции ближе к
149
150
Та б л иц а 1
Суточные профили уровней инсулинемии (инс, мкЕД/мл) и гликемии (глк, ммоль/л) до и после курса лечения [группы 1 – 3]
Вр емя |
Показатель |
|
1 гр уппа ГБ I |
2 гр уппа СД |
± ТЭС |
3 гр уппа СД |
± манинил |
|||
(ч) |
|
|
|
|
|
|
|
|
|
|
|
До |
После |
До |
|
После |
До |
После |
|||
|
|
|
|
|
|
|
|
|
|
|
6 |
инс |
10,321 |
± 1,07 |
14,225 ± 0,4 |
6,168 ± 1,44 |
|
10,967 ± 0,3 |
7,218 ± 0,3 |
9,632 |
± 0,1 |
глк |
4,3 ± |
0,11 |
3,4 ± 0,03 |
9,1 ± 0,22 |
|
6,0 ± 1,04 |
9,0 ± 1,15 |
6,5 ± |
0,11 |
|
|
|
|||||||||
8 |
инс |
15,276 ± 1,22 |
18,117 ± 0,7 |
10,492 ± 1,1 |
|
14,832 ± 1,4 |
9,055 ± 0,2 |
10,956 ± 0,3 |
||
глк |
4,8 ± |
0,07 |
3,5 ± 0,04 |
9,3 ± 0,03 |
|
8,0 ± 0,25 |
9,4 ± 0,03 |
8,9 ± 0,1 |
||
|
|
|||||||||
9 |
инс |
50,776 ± 1,14 |
57,433 ± 0,2 |
25,824 ± 2,6 |
|
51,828 ± 1,7 |
24,856 ± 1,3 |
26,164 ± 0,5 |
||
глк |
8,0 ± 0.07 |
7,8 ± 0,031 |
12,2 ± 1,01 |
|
8,5 ± 1,3 |
12,4 ± 1,2 |
10,3 ± 0,3 |
|||
|
|
|||||||||
10 |
инс |
27,155 ± 1,05 |
30,111 ± 0,1 |
54,726 ± 1,4 |
|
26,234 ± 2,5 |
56,121 ± 0,3 |
71,233 ± 2,6 |
||
глк |
5,1 ± 0,02 |
4,9 ± 0,014 |
12,0 ± 0,84 |
|
7,5 ± 1,3 |
12,2 ± 1,8 |
8,4 ± 1,2 |
|||
|
|
|||||||||
12 |
инс |
21,436 ± 2,03 |
26,335 ± 1,3 |
27,552 ± 1,1 |
|
24,616 ± 0,2 |
29,989 ± 1,1 |
38,137 ± 1,3 |
||
глк |
5,9 ± 0,01 |
6,0 ± 0,0028 |
10,1 ± 0,27 |
|
7,4 ± 1,1 |
10,5 ± 0,2 |
8,2 ± 0,1 |
|||
|
|
|||||||||
13 |
инс |
38,826 ± 2,12 |
43,133 ± 1,1 |
29,872 ± 1,4 |
|
44,056 ± 2,4 |
31,531 ± 0,4 |
32,292 ± 0,2 |
||
глк |
7,4 ± 0,03 |
7,2 ± 0,014 |
12,8 ± 1,3 |
|
8,7 ± 0,01 |
12,9 ± 1,7 |
8,9 ± 1,2 |
|||
|
|
|||||||||
14 |
инс |
40,961 ± 0,02 |
41,053 ± 0,01 |
32,345 ± 2,3 |
|
46,399 ± 1,6 |
34,125 ± 2,4 |
58,948 ± 2,5 |
||
глк |
7,3 ± 0,013 |
7,1 ± 0,02 |
12,3 ± 0,13 |
|
8,0 ± 1,34 |
12,4 ± 0,33 |
7,8 ± 1,3 |
|||
|
|
|||||||||
18 |
инс |
19,352 ± 1,15 |
24,332 ± 0,4 |
20,339 ± 0,3 |
|
22,845 ± 1,1 |
22,222 ± 2,9 |
31,521 ± 1,5 |
||
глк |
5,0 ± 0,011 |
4,9 ± 0,023 |
8,9 ± 1,03 |
|
6,8 ± 0,41 |
9,0 ± 1,06 |
6,2 ± 0,8 |
|||
|
|
|||||||||
19 |
инс |
40,136 ± 1,12 |
43,689 ± 0,4 |
34,545 ± 0,5 |
|
43,258 ± 1,2 |
36,125 ± 1,1 |
39,267 ± 0,3 |
||
глк |
7,4 ± 0,021 |
7,1 ± 0,015 |
10,6 ± 0,22 |
|
7,9 ± 0,3 |
11,1 ± 0,05 |
8,0 ± 1,2 |
|||
|
|
|||||||||
20 |
инс |
25,411 ± 1,17 |
27,331 ± 0,3 |
56,497 ± 1,7 |
|
32,865 ± 2,7 |
58,452 ± 2,1 |
62,245 ± 2,2 |
||
глк |
6,9 ± 0,018 |
6,5 ± 0,011 |
9,9 ± 1,02 |
|
7,3 ± 0,33 |
10,2 ± 0,6 |
7,6 ± 1,02 |
|||
|
|
|||||||||
21 |
инс |
24,121 ± 1,03 |
26,452 ± 0,2 |
45,831 ± 2,4 |
|
25,114 ± 1,9 |
47,106 ± 1,7 |
50,127 ± 1,4 |
||
глк |
6,3 ± 0,032 |
6,2 ± 0,024 |
7,8 ± 0,01 |
|
7,0 ± 0,12 |
7,9 ± 0,3 |
6,6 ± 0,1 |
|||
|
|
|||||||||
03 |
инс |
10,022 ± 0,004 |
9,367 ± 0,003 |
20,717 ± 0,2 |
|
10,246 ± 3,1 |
22,504 ± 1,4 |
14,126 ± 0,2 |
||
глк |
3,8 ± 0,003 |
4,0 ± 0,005 |
7,6 ± 0,12 |
|
6,0 ± 0,06 |
7,8 ± 1,2 |
4,5 ± 0,3 |
|||
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|