

Интерлейкин-2: участие в процессе созревания т-клеток.
Встреча организма с антигеном требует быстрого формирования гуморального или клеточного ответа. Одним из основных участников реализации специфического сигнала от антигена и костимулирующего сигнала от В7 антигенпрезентирующих клеток является интерлейкин-2 (ИЛ-2) .
В результате двухсигнальной активации клона наивных T-клеток начинаются синтез и секреция этого цитокина и одновременная экспрессия на клеточной поверхности его рецептора. ИЛ-2, взаимодействуя с собственным рецептором, обеспечивает быстрое размножение и последующую дифференцировку наивных Т-клеток до зрелых эффекторов.
В процессе инициации синтеза ИЛ-2 ключевая роль принадлежит костимулятору В7. Распознавание антигена Т-клеточным рецептором индуцирует несколько транскрипционных факторов.
Один из них - ядерный фактор активации у Т-клеток (англ. сокр. NF-AT ), который, взаимодействуя с промотором ИЛ-2-гена, обеспечивает транскрипцию соответствующего гена. Однако дерепрессия ИЛ-2-гена не приводит к активной продукции этого цитокина, так как цитокиновые мРНК нестабильны. Взаимодействие CD28 с В7 формирует сигнал, который стабилизирует мРНК этого интерлейкина. В результате такой стабилизаци РНК происходит усиление синтеза ИЛ-2 в 20-30 раз. Т-клетки начинают активно пролиферировать и дифференцироваться в зрелые эффекторные клетки.
Если распознавание антигена Т-клетками происходит в отсутствие костимулирующего сигнала от В7:CD28, то продукция ИЛ-2 крайне низка, и Т-клетки не могут адекватно ответить на антиген.
Важное значение ИЛ-2 в инициации специфического иммунного ответа хорошо иллюстрируется применением лекарственных препаратов, подавляющих отторжение трансплантатов . Хорошо известный в клинике циклоспорин А подавляет продукцию ИЛ-2, нарушая прохождение сигнала от Т-клеточного рецептора , провзаимодействовавшего с антигеном. В итоге формирование антигенспецифического клона Т-клеток подавляется, что обеспечивает длительное выживание трансплантированных органов.
Изменение экспрессии поверхностных молекул T-клеток.
Взаимодействие наивных Т-клеток с антигенным комплексом и костимулятором В7 на поверхности антигенпрезентирующих клеток инициирует полноценный синтез и секрецию ИЛ-2 . Этот цитокин аутокринным способом стимулирует наивные Т-клетки к пролиферации и дифференцировке. После пролиферативной фазы, длящейся 4-5 дня, эти клетки дифференцируются в зрелые эффекторные Т-лимфоциты , которые способны синтезировать все белки, требуемые для выполнения специализированных функций.
Одно из следствий прошедшей дифференцировки Т-клеток состоит в прямом их действии на чужеродные в антигенном отношении клетки без использования каких-либо костимулирующих сигналов.
Обеспечивается подобное прямое эффекторное действие количественным и качественным изменением состава поверхностных молекул T-клеток. Это относится ко всем зрелым Т-клеткам независимо от деления на субпопуляции.
Завершившие дифференцировку Т-клетки характеризуются, во-первых, усилением экспрессии LFA-1 и CD2 молекул, которые позволяют T-клеткам более эффективно взаимодействовать с клетками-мишенями, экспрессирующими чужеродные антигены. Такое усиление особенно существенно для цитотоксических CD8 T-клеток , так как большинство клеток организма характеризуется незначительным уровнем адгезинов ICAM и LFA-3 , которые хорошо представлены только на антигенпрезентирующих клетках .
Второе изменение касается антигенраспознающего Т-клеточного рецепторного комплекса . Тирозинспецифическая фосфатаза ( СD45 ), выполняющая важную функцию в передаче сигнала внутрь клетки с Т-клеточного рецепторного комплекса, представлена на поверхности Т-клеток разными изоформами. Одна из функций этих изоформ - связывание Т-клеточного рецептора с корецепторами CD4 или CD8 , что обеспечивает эффективное прохождение сигнала от антигена внутрь клетки.
И, наконец, зрелые эффекторные Т-клетки теряют L-селектин , который был необходим наивным Т-клеткам при заселении лимфоидной ткани, но оказывается не нужным и даже вредным при развитии иммунного ответа, так как мешает миграции в зону проникновения патогена. Экспрессия L-селектина заменяется интегрином VLA-4 .
Этот адгезин позволяет Т-клеткам связываться с сосудами в зоне воспаления и выполнять тем самым свою антигеннейтрализующую функцию (рис.12.).
Рис. 12.
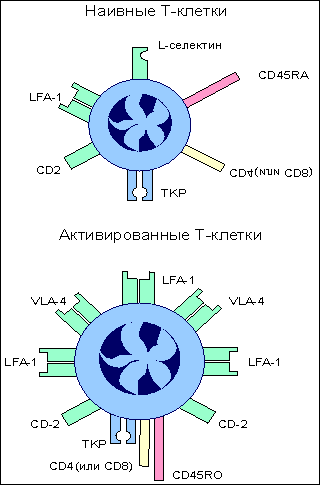
Изменение экспрессии поверхностных молекул при дифференцировке наивных Т-клеток в зрелые эффекторы.
При созревании наивных Т-клеток происходит изменение экспрессии поверхностных молекул, обладающих различными функциональными проявлениями в иммунном ответе. ТКР образует комплекс с ко-рецептором CD4 (или CD8) при участии тирозин-специфической фосфатазы клеточной поверхности CD45; усиливается экспрессия адгезинов LFA-1 и CD2 (подобное усилие особенно важно для цитотоксических CD8 Т-клеток); появляется новый адгезин VLA-1, помогающий связыванию Т-клеток с сосудами в зоне проникновения антигена; в то же время подавляется экспрессия L-селектина, который необходим наивным Т-клеткам при заселении лимфоидной ткани, но становится помехой при миграции зрелых Т-клеток в зону проникновения антигена.