

Наивные cd8 т-клетки: способы активации.
Инициация дифференцировки наивных CD8 T-клеток в зрелые цитотоксические Т-лимфоциты осуществляется несколькими способами.
I.Наиболее простой из них связан с дендритными клетками . Этот тип антигенпрезентирующих клеток характеризуется выраженной экспрессией костимулятора В7 , что само по себе является достаточным условием активации синтеза ИЛ-2 . Данный цитокин обеспечивает активную пролиферацию и дифференцировку наивных Т-клеток, используя аутокринный путь стимуляции (рис. 13).
Рис. 13.
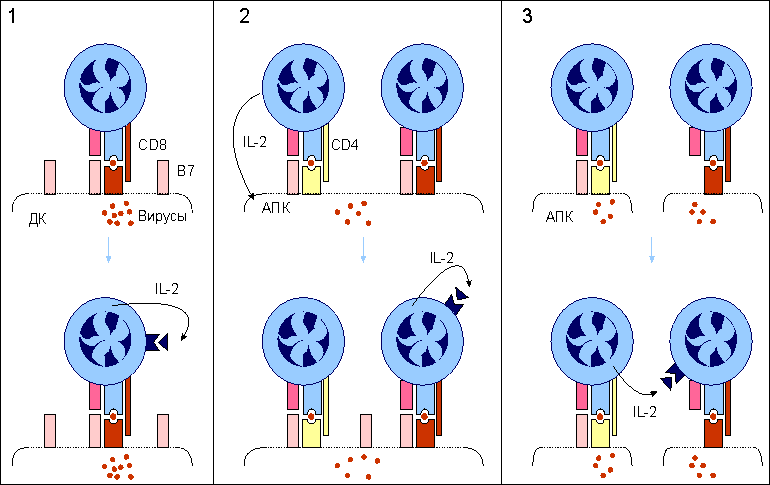
Активация наивных CD8 Т-клеток.
1. Наиболее простой способ активации CD8 Т-клеток наблюдается при распознавании этими клетками иммуногенного комплекса на поверхности дендритных клеток (ДК), обладающих выраженной экспрессией костимулятора В7. Формирование второго сигнала с помощью В7 обеспечивает активную продукцию интерлейкина-2 (IL-2) CD8 Т-клетками. Данный цитокин аутокринным способом активирует наивные CD8 Т-клетки к дифференцировке.
2. В условиях, когда антигенпрезентирующие клетки (АПК) не имеют В7 на своей поверхности, в процесс активации вступают CD4 Т-клетки. Секретируемый CD4 Т-клетками IL-2 после распознавания иммуногенного комплекса на поверхности АПК стимулирует экспрессию В7 этими клетками. Дальнейшее развитие событий неотличимо от первого случая.
3. В условиях, когда CD8 Т-клетки распознают иммуногенный комплекс на клетках, не способных к экспрессии В7 даже при стимуляции, запуск дифференцировки с помощью IL-2 осуществляется по принципу дистанционного управления.
II.В то же время цитотоксический ответ к некоторым вирусам и чужеродным трансплантатам требует присутствия CD4 T-клеток . Возможно, это связано с недостаточной иммуногенностью вирусных и трансплантационных антигенов для формирования первого сигнала от Т-клеточного рецептора или слабой экспрессией В7 , формирующего второй сигнал . При включении в ответ CD4 Т-клеток, которые способны стимулировать антигенпрезентирующие клетки к синтезу В7, создаются нормальные условия для двойной стимуляции наивных CD8 Т-клеток.
III.Третий способ стимуляции наивных CD8 T-клеток к дифференцировки связан с прямым действием на них ИЛ-2 , секретируемого распознавшими антиген CD4 T-клетками . Этот способ проявляется в условиях отсутствия индуцируемой экспрессии В7 под влиянием CD4 Т-клеток.
Какой из этих способов доминирует в организме при формировании цитотоксического ответа, пока неизвестно.
Наивные cd4 т-клетки: дифференцировка в th1 и th2
CD4 T-клетки , покинувшие тимус, не могут непосредственно встать на путь созревания до зрелых эффекторов. В отличие от наивных CD8 T-клеток , они должны пройти дополнительный этап развития на периферии , который заканчивается формированием незрелых ТН0 -клеток. Именно от этих коммитированных клеток в зависимости от ситуации формируются зрелые эффекторы: ТН1-клетки воспаления (CD4 Т-клетки воспаления) и хелперные ТН2-клетки (хелперные CD4 Т-клетки) . Они не различаются по фенотипическим маркерам клеточной поверхности, но известны их разные возможности в синтезе цитокинов .
ТН0 продуцируют интерлейкин-2 и интерлейкин-4 .
ТН1 продуцируют интерлейкин-2 , интерлейкин-3 , гранулоцитарно-макрофагальный колониестимулирующий фактор , интерферон-гамма , фактор некроза опухолей-бета (лимфотоксин) .
ТН2 продуцируют интерлейкин-3 , интерлейкин-4 , интерлейкин-5 , интерлейкин-6 , интерлейкин-10 и гранулоцитарно-макрофагальный колониестимулирующий фактор .
Интерлейкин-3 и гранулоцитарно-макрофагальный колониестимулирующий фактор являются общими для ТН1 и ТН2. Эти два цитокина принимают участие в качестве факторов гемопоэтической дифференцировки в костном мозге . Остальные цитокины являются характерными только для определенной субпопуляции.
Цитокины, свойственные ТН1, активируют в основном макрофаги . Цитокины, свойственные ТН2, активируют В-клетки . Активность именно этих цитокинов определяет разную эффекторную функцию двух субпопуляций.
Механизмы, направляющие дифференцировку Т-клеток в зависимости от иммунологической ситуации пока недостаточно понятны. Однако ряд экспериментальных данных позволяет представить следующую картину.
Макрофаги, захватившие бактерии или вирусы, начинают продукцию интерлейкина-12 , для которого клеткой-мишенью являются наивные Т-клетки. При этом стимулированные макрофагами нормальные киллеры секретируют интерферон-гамма . Два этих цитокина при совместном действии на наивные Т-клетки определяют их развитие в сторону образования CD4 Т-клеток воспаления .
В то же время наивные Т-клетки, стимулированные интерлейкином-4 , дифференцируются в хелперные CD4 T-клетки . Источником интерлейкина-4 могут быть базофилы и тучные клетки . При этом интерлейкин-4 является ингибитором развития Т-клеток воспаления .