

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / v1-186-226
.pdfГилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
216_______________ ГЛАВА 6_______________________________________________________________________________
Рис. 6.36. Последовательность смены мест кроветворения у зародышей мыши. Стрелками показаны возможные пути миграции полипотентных кроветворных стволовых клеток
(СК).
положность этому наличие стволовых клеток в печени зародыша начинает выявляться только после 11 сут. Эти исследователи утверждали, что стволовые клетки мигрируют из желточного мешка в печень зародыша, а затем в селезенку и костный мозг. Клетки желточного мешка зародыша также выполняют функции лимфоцитов до того, как начнет выявляться какая-либо активность стволовых клеток в собственно зародыше (Hofman, Globerson, 1973; Dahl et al., 1980). Таким образом,
считается, что исходная полипотентная стволовая клетка образуется в пределах желточного мешка и мигрирует из него сначала в печень, а затем в кроветворные органы взрослой особи (рис. 6.36).
Современные данные подтверждают представление о том, что стволовые клетки в костном мозге взрослой особи происходят из потомков этих эмбриональных стволовых клеток. Ванг и др. (Wong et al., 1986)
культивировали стволовые клетки из желточного мешка мыши и обнаружили, что некоторые из них дают начало эритроцитам, в которых синтезируется гемоглобин взрослого типа, а не эмбриональный. Напротив, результаты изучения формирования клеток крови у человека (Peschle et al., 1985) показали, что клетки печени 6-недельного зародыша совершенно так же, как клетки желточного мешка 5-недельного зародыша, синтезируют эмбриональные формы гемоглобина (рис. 6.37).
Однако другая группа исследователей (Rossant et al., 1986) утверждает, что миграция стволовых клеток может быть не столь простой, как полагали ранее. Для того чтобы определить идентичность клеточных клонов, эти исследователи использовали химерных мышей; оказалось, что большинство, если не все клоны кроветворных клеток в постнатальной печени, продуцировали только одну линию клеток.
Рис. 6.37. Ранние клетки крови человека. А. Эритробласты из желточного мешка на сроке 5 нед после зачатия. Б. Срез печени 6-недельного зародыша человека; видны крупные красные кровяные клетки (мегалобласты). (Из Peschle et al., 1985: фотографии с любезного разрешения С. Peschle.)
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ МЕЗОДЕРМА И ЭНТОДЕРМА____________________________________ 217
Другими словами, данный клон был либо миелоидным, либо лимфоидным клоном. Этот результат свидетельствует либо о том, что стволовые клетки, которые мигрируют в печень из желточного мешка, являются коммитированными, а не полипотентными, либо о том, что за I7 дней, в течение которых стволовые клетки находились в печени, одна клеточная линия стала преобладающей (доминирующей). Дальнейшие исследования, по-видимому, позволят сделать выбор между этими двумя в равной степени увлекательными возможностями.
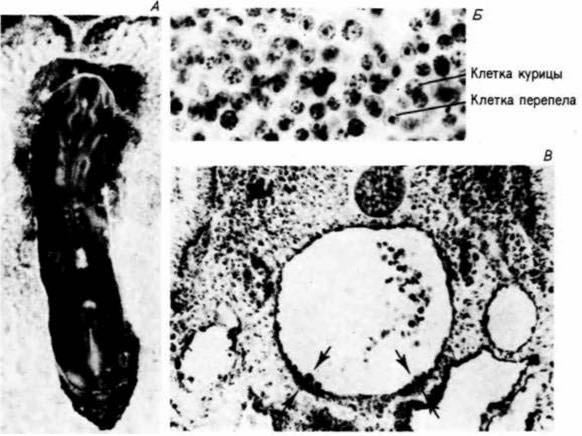
Полагают, что сходная ситуация наблюдается у зародышей птиц, но у них, видимо, имеются два разных источника кроветворных стволовых клеток. Первый, как и у зародышей млекопитающих, локализован в стенке желточного мешка. Эти клетки, однако, представляют собой временную популяцию. Главным источником стволовых клеток являются локальные островки, которые формируются в собственно зародыше. Это открытие было сделано в серии изящных опытов Дитерлен-Ливр, которая пересаживала бластодерму японского перепела на желточный мешок куриного зародыша (рис. 6.38). Клетки курицы легко отличить от клеток перепела, ядра которых значительно сильнее окрашиваются и поэтому служат постоянным маркером для различения клеток двух типов. Использование таких «химерных желточных мешков» показало (DieterlenLievre, Martin, 1981), что стволовые клетки желточного мешка не поставляют клетки взрослому животному; настоящие стволовые клетки формируются в узелках мезодермы, которая выстилает брыжейки и главные кровеносные сосуды. До сих пор считают, что у млекопитающих желточный мешок является
Рис. 6.38. Маркировка клеток крови посредством создания химер «курица — перепел». Α. Фотография «химерного желточного мешка», т.е. бластодермы куриного зародыша, пересаженной на желточный мешок перепела. Б. Фотография, на которой видны клетки курицы и перепела в тимусе химерного животного, различающиеся по окраске ядер. Все лимфоидные клетки куриного происхождения, тогда как структурные клетки тимуса происходят из клеток бластодермы перепела, Б. Срез аорты трехдневного куриного зародыша; стрелками указаны клетки, которые дадут начало кроветворным стволовым клеткам. Если клетки из этой области взять у зародышей перепела и поместить на куриный зародыш, то кровь у куриного зародыша будет кровью перепела. (Из Martin et al., 1978; DieterlenLièvre, Martin, I981;
фотографии с любезного разрешения F. Dieterlen-Lievre.)
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
218_______________ ГЛАВА 6________________________ ;______________________________________________________
.
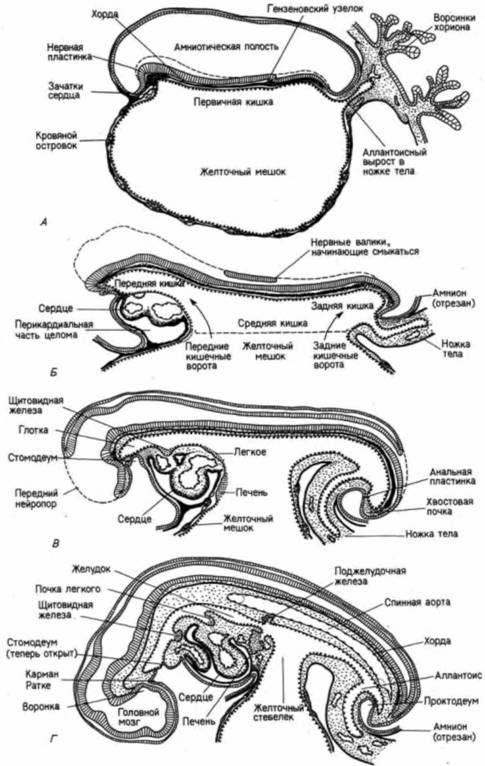
Рис. 6.39. Формирование пищеварительной системы у человека. Схематические изображения сагиттальных срезов зародышей в возрасте примерно: А — 16 сут; Б — 18 сут; В — 22 сут; Г — 28 сут. (Из Carlson, 1981.)
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
_______________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. МЕЗОДЕРМА И ЭНТОДЕРМА____________________________________ 219
источником всех кроветворных стволовых клеток; однако результаты некоторых исследований (Kubai, Auerbach, 1983) показывают, что эти клетки-предшественники стволовых клеток желточного мешка сами могут происходить из стволовых клеток, которые находятся в выстилке брюшной полости.
ЭНТОДЕРМА
Из энтодермы развиваются органы пищеварения и связанные с ними железы, а также органы дыхания (рис. 6.39). По мере того как концы энтодермы загибаются в вентральном направлении и перемещаются к центру зародыша, формируются передняя кишка и задняя кишка, представляющие собой части будущей пищеварительной трубки. На месте будущего ротового отверстия образуется эктодермальное углубление
— стомодеум, которое потом приходит в контакт со стенкой передней кишки. Область их объединения называется ротовой пластинкой. Эта пластинка позднее (у эмбриона человека в возрасте около 22 сут) разрывается, а стомодеум превращается в ротовую полость. Эктодермальный эпителий крыши ротовой полости образует выступ (карман Ратке), который растет навстречу нейральному выросту дна зачатка промежуточного мозга — воронке или инфундибулюму. Эти два эктодермальных зачатка взаимодействуют друг с другом и вместе образуют гипофиз. Из кармана Ратке развивается железистая часть гипофиза, а из воронки — его нервная часть. Таким образом, гипофиз имеет двойное происхождение, и эта двойственная природа гипофиза отражается на его функции во взрослом организме.
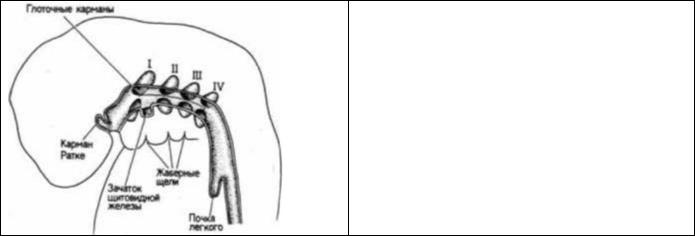
Энтодермальная часть пищеварительной трубки начинается в глотке. Здесь у зародыша млекопитающих образуются четыре глоточных кармана. Борозды между этими карманами иногда называют жаберными щелями, потому что они напоминают такие же структуры у зародышей рыб (рис. 6.40). Однако вместо того, чтобы дать начало жабрам, развитие глоточных карманов у млекопитающих изменилось в соответствии с наземной средой обитания. Первая пара глоточных карманов преобразуется в слуховые полости среднего уха и связанные с ними евстахиевы трубы. Из второй пары карманов развиваются стенки миндалин. Тимус образуется из третьей пары глоточных карманов. На более поздних стадиях развития эта железа будет обеспечивать дифференцировку лимфоцитов. Из этой же пары глоточных карманов происходит одна пара паращитовидных желез, а вторая их пара возникает из четвертой пары глоточных карманов. В дополнение к этим парным карманам между глоточными карманами второй пары на дне глотки формируется маленький центрально расположенный вырост (дивертикул). Этот энтодермально-мезенхимный дивертикул отпочковывается
Рис. 6.40. Глоточные карманы и жаберные щели у 5- недельного зародыша человека. (По Carlson, 1981.)
от глотки и мигрирует в область шеи, где дифференцируется в щитовидную железу. Здесь же от стенки глотки отшнуровывается зачаток дыхательной трубки — ларинготрахеальная борозда.
Пищеварительная трубка и ее производные
Позади глотки пищеварительная трубка сужается, переходя в пищевод, за которым следуют желудок, тонкий кишечник и толстый кишечник. Энтодермальные клетки образуют только выстилку пищеварительной трубки и ее желез, тогда как мезодермальные мезенхимные клетки, которые окружают эту трубку, дифференцируются в мышцы, обеспечивающие перистальтику.
На рис. 6.41 видно, что желудок развивается подобно глотке, как расширенный участок трубки. Каудальнее формируется кишечник, и связь между ним и желточным мешком в конечном счете прерывается. На каудальном конце кишечника возникает углубление, в котором энтодерма встречается с лежащим над ней покровным эпителием. Тонкая клоакальная мембрана разделяет эти две ткани. Впоследствии она разрывается, и образуется отверстие, которое станет анальным.
Печень, поджелудочная железа и желчный пузырь
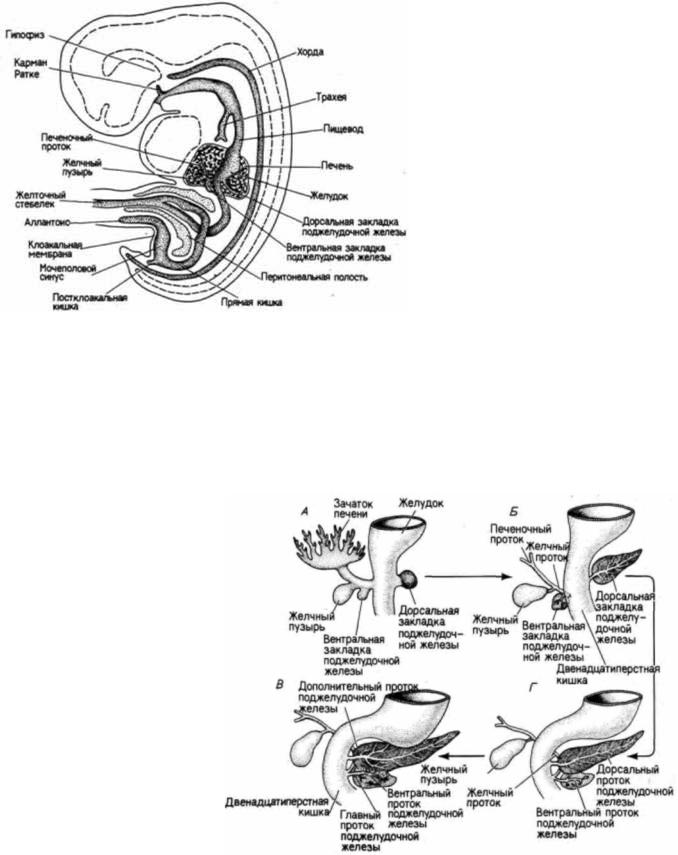
Энтодерма также образует выстилку трех дополнительных органов, которые развиваются непосредственно за желудком, каудальнее его. Печеночный дивертикул представляет собой энтодермальную трубку, которая простирается от передней кишки в окружающую мезенхиму. Мезенхима индуцирует
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
220_______________ ГЛАВА 6_______________________________________________________________________________
Рис. 6.41. Пищеварительная система 6-недельного зародыша человека. Область желудка в пищеварительной трубке начала расширяться; поджелудочная железа представлена двумя закладками, которые впоследствии сольются. (По
Langman, 1981.)
энтодерму, которая в ответ на ее воздействие пролиферирует, ветвится и формирует железистый эпителий печени. Часть печеночного дивертикула (область, ближайшая к пищеварительной трубке) продолжает функционировать как дренажный проток печени, а из ответвления от этого протока образуется желчный пузырь (рис. 6.42). Поджелудочная железа развивается в результате слияния четко выраженных дорсального и вентрального дивертикулов. Оба этих зачатка возникают из области энтодермы, расположенной непосредственно за желудком; по мере их роста они сближаются и в конце концов сливаются. У взрослого человека сохраняется только вентральный проток, несущий
Рис. 6.42. Развитие поджелудочной железы у человека. На 30 сут. развития зародыша (А) вентральная закладка поджелудочной железы расположена вблизи зачатка печени. К 35 сут. железа начинает мигрировать назад (Б). В течение шестой недели развития вентральная закладка поджелудочной железы приходит в контакт с дорсальной закладкой (В). У большинства людей дорсальная закладка железы утрачивает свой проток в двенадцатиперстную кишку. Однако примерно у 10% популяции сохраняется система двух протоков (Г). (По Langman, 1969.)
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. МЕЗОДЕРМА И ЭНТОДЕРМА 221
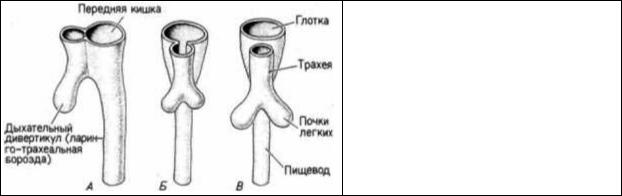
Рис. 6.43. Разделение передней кишки на пищевод и дыхательный дивертикул (трахею) в течение третьей и четвертой недель беременности у человека. А. Вид сбоку (конец третьей недели). Б, В. Вид с брюшной стороны (четвертая неделя беременности). (По
Langman, 1969.)
пищеварительные ферменты в кишечник. У других видов (например, у собак) оба протока — дорсальный и вентральный — открываются в кишечник.
Дыхательная трубка
Легкие также являются производными пищеварительной трубки, хотя они совсем не участвуют в пищеварении. В центре задней области глотки между четвертой парой глоточных карманов появляется ларинго-трахеальная борозда, которая тянется по вентральной поверхности передней кишки (рис. 6.43). Эта борозда затем раздваивается на две главные ветви, которые образуют легкие. Ларинготрахеальная энтодерма становится выстилкой трахеи, двух бронхов и воздушных мешков (альвеол) легких. Как мы увидим в одной из последующих глав, ветвление этой энтодермальной трубки зависит от взаимодействий между разными типами мезодермальных клеток на ее пути. Легкие являются эволюционным новшеством, и они последними из органов млекопитающих завершают свою дифференцировку. Легкие должны быть способны к поглощению кислорода с первым вдохом ребенка. Чтобы это осуществить, альвеолярные клетки секретируют в жидкость, омывающую легкие, поверхностноактивное вещество — сурфактант. Этот сурфактант, состоящий из фосфолипидов, таких, как сфингомиелин и лецитин, секретируется на поздних стадиях беременности, и необходимый физиологически полезный уровень их содержания достигается у человека обычно на 34-й неделе беременности. Эти вещества дают альвеолярным клеткам возможность соприкасаться не слипаясь. Дети, родившиеся преждевременно, часто испытывают затруднения в дыхании, и их приходится держать в респираторных камерах (где длительное время поддерживается искусственное дыхание) до тех пор, пока не созревают клетки, секретирующие сурфактант.
Дополнительные сведения и гипотезы: Тератокарцинома
Половая клетка млекопитающих или ранний бластомер содержит в себе всю информацию, необходимую для последующего развития. Что может случиться, если такая клетка станет злокачественной? В этом случае возникает тератокарцинома. Опухоли этого типа иногда спонтанно развиваются в яичнике или в семеннике. Образование тератокарцином можно вызвать экспериментально, имплантируя бластоцисту не в матку, а в иное место (например, внутрь соединительнотканной капсулы, окружающей почку). Независимо от того, возникла ли тератокарцинома спонтанно или в результате экспериментального вмешательства, она содержит популяцию недифференцированных стволовых клеток, которые по биохимическим свойствам и по способностям к развитию похожи на клетки внутренней клеточной массы (Graham, 1977). Эти стволовые клетки (называемые клетками эмбриональной карциномы) не только делятся, но могут также дифференцироваться в самые разнообразные ткани, в том числе в кишечный и дыхательный эпителий, мышечную, нервную, хрящевую и костную ткани (рис. 6.44). Образующиеся дифференцированные клетки больше не делятся и потому не являются злокачественными. Итак, эти опухоли могут дать начало самым разнообразным тканям, встречающимся в организме.
Эмбриональные карциномы можно переносить из одной мыши в другую; их клетки можно индуцировать к образованию эмбриоидных тел. инъецируя эти клетки в перитонеальную (брюшную) полость.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
222 ___________ ГЛАВА 6___________________
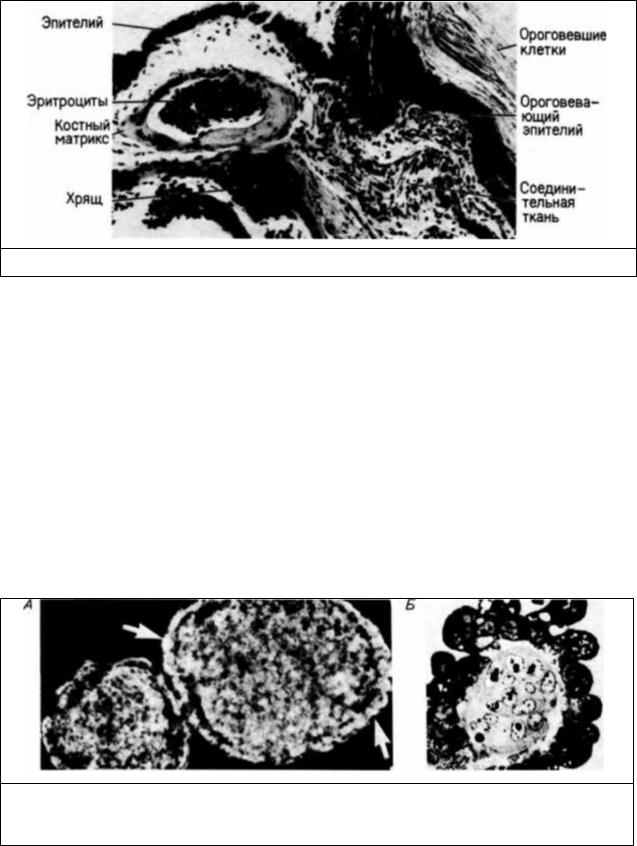
Рис. 6.44. Микрофотография среза карциномы. Видны дифференцированные клетки многих типов. (Из Gardner, 1982; фотография из С. Graham с любезного разрешения R. L. Gardner.)
где затем появляются аморфные скопления дифференцированных клеток (в том числе кроветворные колонии) в «энтодерме» (рис. 6.45). Было показано (Adamson et al., 1977), что эта «энтодерма» способна синтезировать α- фетопротеин, белок, обычно синтезируемый нормальными энтодермальными клетками млекопитающих. Следовательно, клетки эмбриональной карциномы имитируют раннее развитие млекопитающих, но развитие формируемой ими опухоли имеет случайный, неупорядоченный характер.
Эти опухоли, вероятно, происходят из половых клеток, однако по своим биохимическим, морфологическим и поверхностным свойствам образующие ее клетки напоминают бластомеры внутренней клеточной массы. Стюарт и Минц (Stewart, Mintz, 1981) получили мышь из клеток, часть которых происходила из стволовой клетки тератокарциномы! Стволовые клетки из тератокарциномы мышей линии агути культивировали в течение нескольких клеточных поколений и убедились, что они сохраняют хромосомный набор, характерный для родительской мыши. Затем изолированные стволовые клетки этого типа инъецировали в бластоцисты белых мышей. Бластоцисты переносили в матку приемной матери, которая родила живых мышат. Шкурки у некоторых мышат были двухцветными; это свидетельствовало о включении опухолевой клетки в развитие зародыша (рис. 6.46). Более того, когда химерную мышь спаривали с мышью, несущей соответствующий маркер, некоторые из рожденных ею мышат имели черты, характерные для фенотипа опухолевого «родителя». Злокачественная клетка эмбриональной карциномы образовала многие, если не все, типы нормальных соматических клеток и даже нормально функционирующие половые клетки! Если спаривали
Рис. 6.45. Эмбриоидные тела. А. Живые эмбриоидные тела в культуре (фотографирование с помощью фазовоконтрастного устройства). Стрелки указывают области, в которых можно видеть клетки двух разных типов (внутренние и наружные). Б. Срез такого эмбриоидного тела: препарат окрашен гематоксилином и эозином. Стволовые клетки (в центре) окружены клетками, очень напоминающими раннюю эмбриональную энтодерму. (Фотографии с любезного разрешения G. Martin.)
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
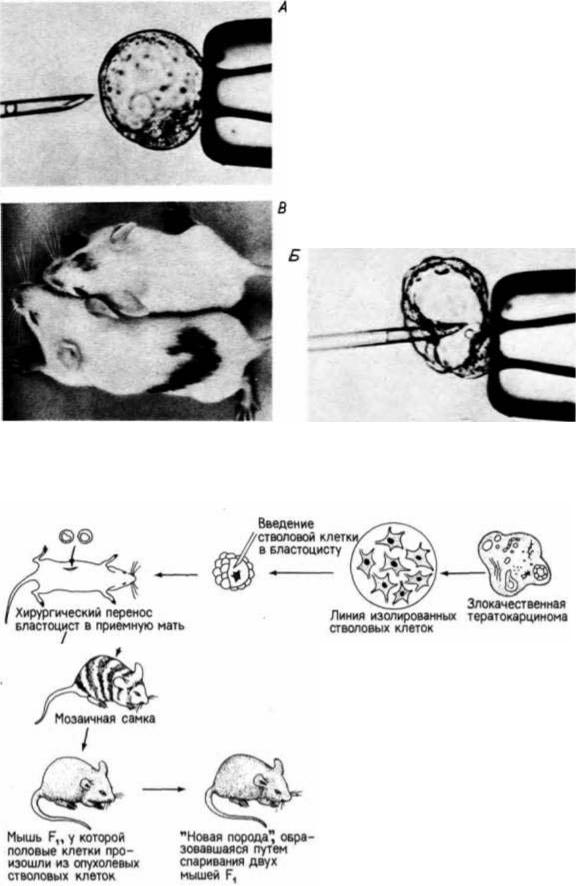
Рис. 6.46. Введение клетки эмбриональной карциномы в бластоцисту мыши. А. Микропипетка со стволовой клеткой тератокарциномы приближается к бластоцисте, прочно удерживаемой обычной пипеткой. Б. Инъекция стволовой клетки во внутреннюю клеточную массу. В. Мыши, развившиеся в опыте, в котором стволовая клетка тератокарциномы из черной линии мышей интегрировалась во внутреннюю клеточную массу бластоцисты мышей белой линии. На голове и спине мышей можно видеть черный мех, указывающий на то, что по крайней мере пигментные клетки были опухолевого происхождения. (А и Б — с с любезного разрешения K. Illmense; В – из Papaioannou, 1979; фотография с любезного разрешения V.E. Papaioannou. )
Рис. 6.47. Схема опыта с получением мышей, гены которых принадлежат главным образом опухолевым клеткам. Из злокачественной карциномы мыши изолировали линию стволовых клеток и вводили их в бластоцисты мыши другой линии. Химерные бластоцисты помещали для развития в матку приемной матери. Если опухолевые клетки интегрируются в бластоцисту, то многие из клеток у родившейся мыши происходят из клеток опухоли. Если из стволовых клеток опухоли произошли и половые клетки, то мозаичную мышь можно спарить с нормальной мышью и получить потомство F1. Мыши F1 должны быть гетерозиготными по всем хромосомам опухолевой клетки. Спаривание между мышами F1 дает мышей F2, имеющих гомозиготные гены, происходящие из опухолевых клеток. (По Stewart, Mintz, 1981.)

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
224 |
ГЛАВА 6 |
мышей, получивших опухолевую клетку от одного из родителей, то в потомстве оказывались мыши, гомозиготные по большому числу генов опухолевой клетки (рис. 6.47).
Поскольку культивирование ранних зародышей млекопитающих является трудной задачей, клетки эмбриональной карциномы дают исследователям уникальную возможность для анализа раннего развития млекопитающих. Кроме того, благодаря способности клеток эмбриональной карциномы мутировать в культуре при инъекции таких клеток в бластоцисты можно также создать мышей, мутантных почти по любому гену.
На этом мы завершим обзор основных черт развития животных и обратимся к механизмам, которые делают возможным это развитие. Во втором томе книги мы рассмотрим различные способы, с помощью которых осуществляется дифференцировка клеток, а третий том будет посвящен вопросам взаимодействия клеток между собой при формировании тканей и органов.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ МЕЗОДЕРМА И ЭНТОДЕРМА____________________________________225