

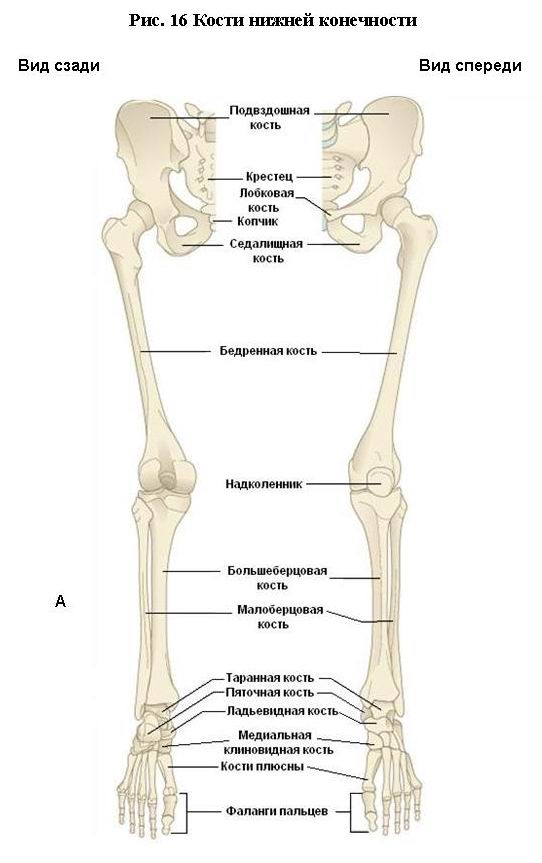
Кости нижней конечности
Нижняя конечность человека является органом опоры и передвижения, и ее строение наилучшим образом приспособлено к выполнению этой важной функции. Скелет нижней конечности, являющейся у человека органом опоры и перемещения тела в пространстве, состоит из более толстых и массивных костей, соединенных между собой менее подвижными сочленениями, чем у верхних конечностей.
Нижняя конечность человека состоит из пояса (это тазовые кости, между которыми сзади как бы вклинивается крестец) и свободной нижней конечности. Таким образом, образуется прочный таз (пояс нижних конечностей), имеющий арочное строение, несущий на себе тяжесть туловища и передающий ее массивным костям свободной нижней конечности.
Пояс нижних конечностей образован двумя тазовыми костями, каждая из которых сзади сочленяется с крестцом, а спереди друг с другом. Скелет свободной нижней конечности гомологичен со скелетом верхней конечности и также состоит из трех сегментов: проксимального (бедро), среднего (две кости голени: большеберцовая и малоберцовая) и дистального. В области коленного сустава имеется крупная сесамовидная кость - надколенник.
Дистальный сегмент свободной нижней конечности - стопа также подразделяется на три части: предплюсну, плюсну и фаланги пальцев (рис. 16).
Кости пояса нижней конечности. Тазовая кость (os сохае) - парная плоская кость, образована подвздошной, лобковой и седалищной костями, срастающимися между собой в области вертлужной впадины - глубокой ямки, сочленяющейся с головкой бедренной кости. Подвздошная кость расположена над вертлужной впадиной, лобковая кпереди и книзу, седалищная - книзу и сзади от нее. Седалищная и лобковая кости ограничивают запирательное отверстие овальной формы и больших размеров, затянутое соединительнотканной запирательной мембраной.
У новорожденного вертлужная впадина уплощена (больше у девочек), кости в этом месте соединены между собой прослойками хряща. Хрящевая пластинка между седалищной и лобковой костями исчезает примерно в 6-летнем возрасте. Окончательное сращение трех костей происходит в 12 - 15 лет у девочек и в 13 - 16 лет у мальчиков. С возрастом впадина углубляется.
Подвздошная кость (osilium) состоит из массивного тела и тонкого крыла, оканчивающегося вверху подвздошным гребнем. Концы гребня выступают спереди и сзади в виде верхних и нижних передних и задних подвздошных остей. На гребне прикрепляются широкие мышцы живота. Там, где сходятся края подвздошной и седалищной костей, под нижней задней подвздошной остью располагается большая седалищная вырезка, ограниченная снизу седалищной костью. Вогнутая внутренняя поверхность крыла подвздошной кости формирует подвздошную ямку, которая снизу ограничена дугообразной линией, простирающейся от ушковидной поверхности до лобкового гребня. На ягодичной поверхности подвздошной кости имеются три ягодичные шероховатые линии (передняя, задняя, нижняя), к которым прикрепляются ягодичные мышцы.
Крестцово-тазовая поверхность несет на себе ушковидную поверхность, сочленяющуюся с одноименной поверхностью крестца.
Седалищная кость (os ischii) имеет тело, которое участвует в формировании вертлужной впадины. Ветвь седалищной кости участвует в ограничении запирательного отверстия и образует мощный седалищный бугор, сзади и выше которого располагается малая седалищная вырезка.
Лобковая кость (os pubis) также имеет тело, участвующее в формировании вертлужной впадины, и две ветви - верхнюю и нижнюю, - соединяющиеся между собой под углом. На медиальной поверхности угла имеется сuмфизиальная поверхность, которая, соединяясь с такой же поверхностью противоположной кости, образует лобковый симфиз. Задний край верхней ветви заострен. Это лобковый гребень, который переходит в дугообразную линию подвздошной кости, тем самым, образуя пограничную линию, отделяющую большой таз от малого. На расстоянии 1,5 - 2 см от симфиза гребень утолщается, образуя лобковый бугорок. На границе с подвздошной костью находится подвздошно-лобковое возвышение.
Кости свободной нижней конечности. Бедренная кость (os femoris) - наиболее крупная, массивная трубчатая кость, изогнутая кпереди. Шаровидная головка бедренной кости, сочленяющаяся с вертлужной впадиной тазовой кости, несет на своей поверхности ямку, куда прикрепляется круглая связка. Длинная шейка, соединяющая головку с телом кости, расположена под углом к последнему.
У мужчин этот угол тупой (около 130 град.), у женщин - почти прямой. Тотчас под шейкой латерально расположен большой вертел, у основания которого имеется углубление - вертельная ямка, обращенная к шейке бедренной кости. С медиальной стороны находится малый вертел. Оба вертела соединены спереди межвертельной линией, сзади - межвертельным гребнем. Тело бедренной кости имеет цилиндрическую форму, спереди его рельеф гладкий, сзади проходит шероховатая линия, в которой различают латеральную губу, вверху оканчивающуюся ягодичной бугристостью, и медиальную, переходящую проксимально в гребенчатую линию. К описанным буграм, линиям, гребням прикрепляются мышцы, их работа и обусловливает появление на кости данных структур. Внизу обе губы также расходятся, образуя треугольную подколенную поверхность.
Более сложно устроен нижний эпифиз бедренной кости. На нем выделяются два мощных мыщелка: медиальный и латеральный, которые несут на своих боковых поверхностях одноименные надмыщелки. Мыщелки, причем медиальный больше латерального, разделены глубокой межмыщелковой ямкой, переходящей впереди в надколенниковую поверхность, к которой прилежит надколенник.
Надколенник (patella) представляет собой сесамовидную кость, лежащую в толще сухожилия четырехглавой мышцы бедра. Верхушка надколенника обращена вниз, основание - вверх, суставная поверхность, покрытая хрящом, - назад. Надколенник легко прощупывается у живого человека.
Большеберцовая кость (tibia) - единственная из двух костей голени, которая сочленяется с бедренной. Это обусловило ее большую массивность. Верхний эпифиз мощный, широкий, имеет два мыщелка: медиальный и латеральный, - несущие на своих проксимальных концах слегка вогнутые суставные поверхности, разделенные межмыщелковым возвышением. На латеральной поверхности одноименного мыщелка большеберцовой кости имеется малоберцовая суставная поверхность.
Тело большеберцовой кости имеет трехгранную форму. Острый передний kрай возле верхнего эпифиза переходит в выраженную бугристость большеберцовой кости - место прикрепления сухожилия четырехглавой мышцы бедра. Он отделяет медиальную поверхность от латеральной. К латеральному (межкостному) краю прикрепляется межкостная перепонка голени. Медиальный край разграничивает медиальную и заднюю поверхности. Дистальный эпифиз примерно четырехугольной формы, несет на себе нижнюю суставную поверхность для сочленения с таранной костью стопы.
Медиальный конец его оттянут книзу и образует медиальную лодыжку. На латеральной стороне нижнего эпифиза имеется малоберцовая вырезка для сочленения с малоберцовой костью.
Малоберцовая кость (fibula) - тонкая длинная трубчатая кость. Верхний эпифиз - головка - несет на себе суставную поверхность для сочленения с верхним эпифизом большеберцовой кости и заканчивается заостренной верхушкой. Посредством шейки головка переходит в тело трехгранной формы, которое внизу оканчивается утолщенной латеральной лодыжкой, снабженной суставной поверхностью лодыжки. Нижняя суставная поверхность большеберцовой кости и суставные поверхности лодыжек образуют вилку, которая охватывает блок таранной кости сверху и с боков.
Кости (скелет) стопы (ossa pedis). В стопе различают предплюсну, плюсну и пальцы. Стопа человека выполняет строго специализированную функцию передвижения и опоры. С этим связано ее строение по типу прочной и упругой сводчатой арки с короткими пальцами. Основные особенности стопы современного человека - это наличие сводов, прочность, пронированное положение, укрепление медиального края, укорочение пальцев, укрепление и приведение I пальца, который не противопоставляется остальным, и расширение его дистальной фаланги. Кости предплюсны, испытывающие большую нагрузку, массивные, прочные.
Предплюсна (tarsus). Представлена семью короткими костями, расположенными в два ряда. В проксимальном (заднем) ряду находятся таранная и пяточная кости, в дистальном (переднем) латерально располагается кубовидная кость, медиально - узкая ладьевидная, впереди нее три клиновидные кости: медиальная, промежуточная и латеральная.
Таранная кость (talus) состоит из тела, шейки и головки.
На верхней поверхности тела расположен блок, имеющий три суставные поверхности (верхнюю, медиальную и латеральную лодыжковые), сочленяющиеся с соответствующими поверхностями костей голени. На нижней поверхности таранной кости находятся три пяточные суставные поверхности: задняя, средняя и передняя. Между задней и средней поверхностями проходит борозда таранной кости; позади блока отходит задний отросток. В борозде, разделяющей задний отросток на две части, проходит сухожилие длинного сгибателя большого пальца. Головка таранной кости овальной формы, сочленяется с ладьевидной костью.
Наиболее крупная пяточная кость (calcaneus), сочленяясь с таранной костью вверху и кубовидной спереди, несет на себе соответствующие суставные поверхности. Важной структурой является опора таранной кости - костный выступ, поддерживающий головку таранной кости. Между средней и задней таранными суставными поверхностями проходит борозда пяточной кости, которая, соединяясь с соответствующей бороздой таранной кости, формирует пазуху предплюсны, где находится мощная связка, удерживающая пяточную и таранную кости. Вход в пазуху открывается на латеральной стороне тыла стопы. Кзади пяточная кость заканчивается мощным пяточным бугром.
Ладьевидная, кубовидная и три клиновидные кости соединяются между собой, а первые две, кроме того, с пяточной и таранной; клиновидные и кубовидная - с плюсневыми костями.
Ладьевидная кость (os naviculare) лежит медиально. Ее проксимальная вогнутая суставная поверхность сочленяется с головкой таранной кости, а выпуклая дистальная несет на себе три плоские суставные поверхности для соединения с клиновидными костями. На медиальном крае ладьевидной кости расположена ее бугристость, к которой прикрепляется задняя большеберцовая мышца.
Три клиновидные кости (ossa cuneiformia) лежат кпереди от ладьевидной кости, занимают медиальную часть предплюсны и сочленяются с основаниями плюсневых костей: наиболее крупная медиальная клиновидная кость - с первой плюсневой костью, промежуточная - со II, латеральная - с III.
Кубовидная кость (os cuboideum) занимает латеральный край предплюсны, она лежит между пяточной и IV - V плюсневыми костями, с которыми сочленяется. На подошвенной поверхности кубовидной кости располагается ее бугристость.
Плюсна (ossa metatarsi). Представлена пятью короткими трубчатыми костями, в каждой из которых различают основание, тело и головку. I плюсневая кость наиболее короткая и толстая, II - наиболее длинная. Тела плюсневых костей выпуклые в сторону тыла стопы, форма их призматическая. Своими основаниями плюсневые кости сочленяются с клиновидными и кубовидной костями, а головками - с основаниями соответствующих проксимальных фаланг.
Скелет пальцев образован фалангами (рhаlаnх) - короткими трубчатыми костями. Количество их соответствует фалангам пальцев кисти, однако они отличаются небольшими размерами. Фаланги I пальца, особенно дистальная, имеют большие размеры, чем фаланги II - V пальцев стопы. Каждая фаланга состоит из основания, тела и головки. Отличительной особенностью дистальных фаланг является наличие бугристости. Каждая проксимальная фаланга своим основанием сочленяется с соответствующей плюсневой костью, а головкой - со средней фалангой. Средние фаланги сочленяются с основаниями дистальных фаланг.
|
|
|
|
|||
|
Скелет вместе с мышцами выполняет функции опоры и движения благодаря тому, что все кости соединены между собой и образуют подвижные костные рычаги. При этом кости образуют пассивную, а мышцы - активную часть опорно-двигательного аппарата. Характер соединений зависит от функции того или иного костного звена.
Кости низших позвоночных, ведущих водный образ жизни, соединены посредством непрерывных соединений. Выход на сушу привел к изменению характера движений, в связи с этим сформировались переходные формы (симфизы) и наиболее подвижные прерывные соединения - суставы. В соответствии с этим в онтогенезе человека все соединения костей проходят две стадии развития, напоминающие таковые в филогенезе, вначале непрерывные, затем синовиальные (суставы).
В теле человека все соединения костей делятся на три большие группы: непрерывные, полусуставы (симфизы) и прерывные, или синовиальные (суставы).
Непрерывные соединения - это соединения костей, осуществляющиеся с помощью различных видов соединительной ткани. При этом отсутствует суставная щель или полость между соединяющимися костями. Непрерывные соединения весьма прочны, однако их подвижность ограничена или вообще отсутствует. В зависимости от характера ткани, соединяющей кости, различают фиброзные, хрящевые и костные соединения (рис. 17).
В прочных фиброзных соединенuях (сuндесмозах) кости соединены между собой плотной волокнистой соединительной тканью. Сuндесмоз (syndesmosis) - это соединение костей с помощью связок, мембран, швов, «вколачивания», коллагеновые волокна которых срастаются с надкостницей, переходят в нее без четкой границы. Связки представляют собой толстые, образованные плотной волокнистой соединительной тканью пучки или пластины, которые перекидываются от одной кости к другой, укрепляя суставы или ограничивая их движения. Большинство связок образовано пучками коллагеновых волокон. Однако встречаются связки, состоящие из пучков эластических волокон, например желтые связки, натянутые между дугами позвонков. Они растягиваются при сгибании позвоночного столба и благодаря своей эластичности вновь укорачиваются, способствуя разгибанию позвоночника.
Межкостные перепонки (membranae interоsseae) представляют собой соединительнотканные пластины, натянутые, например, между диафизами длинных трубчатых костей предплечья и голени. Они прочно удерживают одну кость возле другой, служат местом начала многих мышц. Межкостные перепонки сформированы пучками коллагеновых волокон, образующих слои, направленные от одной кости к другой.
Швы (suturae) - это соединения краев костей крыши и лицевого отдела черепа между собой с помощью тонких прослоек волокнистой соединительной ткани. Надкостница, не прерываясь, покрывает линию шва. Между соединяющимися краями костей черепа находится тонкая прослойка волокнистой соединительной ткани. С возрастом коллагеновые волокна кальцифицируются и волокнистая соединительная ткань превращается в ретикулофиброзную (грубоволокнистую) костную ткань. В зависимости от конфигурации краев соединяющихся костей различают зубчатый, плоский и чешуйчатый швы. Разновидностью фиброзного соединения является вколачивание (gomphosis) и зубоальвеолярное соединение (articulatio dеntоаlvеоlаris) - соединение зуба с костной тканью зубной альвеолы. Швы, а также «вколачивание» представляют собой прочные, эластичные, малоподвижные, даже практически неподвижные соединения костей черепа.
Хрящевые соединения, или синхондрозы (synchondrosis), представляют собой соединения костей с помощью волокнистой хрящевой ткани. Волокнистый хрящ, подобно другим типам хряща, состоит из небольшого количества хондроцитов и межклеточного вещества. Хондроциты не отличаются от описанных ранее. Они расположены в узких лакунах. Межклеточное вещество образовано пучками коллагеновых волокон, имеющих весьма упорядоченное строение, которое определяется направлением сил натяжения и сжатия. Волокна, образующие пучки толщиной 40 - 70 нм, соединяются тонкими волокнами, а количество аморфного вещества волокнистого хряща невелико.
Синхондрозы отличаются прочностью, упругостью и малой подвижностью, степень которой зависит от толщины и структуры хрящевой прослойки между костями. Крайне редко хрящ между соединяющимися костями сохраняется в течение всей жизни. Такие синхондрозы являются постоянными (например, между ребрами и грудиной). Большинство синхондрозов временные, ибо хрящевая прослойка между костями сохраняется лишь до определенного возраста, после чего хрящ замещается костной тканью.
Костные соединения - синостозы (synostosis) - появляются по мере окостенения синхондрозов между отдельными костями основания черепа, костями, составляющими тазовую кость, и др.
При этом в межклеточном веществе волокнистого хряща откладываются кристаллы гидроксиапатита и аморфного трикальцийфосфата (В. Н. Павлова и др., 1988). Кристаллы гидроксиапатита ориентированы вдоль продольной оси коллагеновых волокон, они расположены в промежутках между молекулами тропоколлагена и на поверхности волокон. Неколлагеновые белки аморфного вещества также связывают ионы кальция и фосфата и кристаллы гидроксиапатита. Важную роль в кальцификации играют матриксные пузырьки, которые представляют собой округлые мембранные гранулы диаметром 30-100 нм. Пузырьки образуются из отростков хондроцитов. Первые кристаллы гидроксиапатита в кальцифицирующемся хряще обнаруживаются в пузырьках.
Симфизы (от греч. symphysis - срастание) также представляют собой хрящевые соединения, лишенные суставной капсулы. В толще хряща имеется небольшая, заполненная жидкостью щелевидная полость, синовиальная оболочка отсутствует. К этому типу соединений относятся межпозвоночные симфизы, лобковый сuмфuз и сuмфиз рукоятки грудuны Соединяющиесякости связаны волокнистым хрящом, образованным переплетающимися между собой мощными S-образными пучками коллагеновых волокон, которые вплетаются в надкостницу и в гиалиновый хрящ, покрывающий обращенные друг к другу симфизиальные поверхности.
Суставы, или синовиальные соединения (аrticulаtionеs synoviales), представляют собой прерывные соединения костей, у которых между соединяющимися костями всегда имеется «прерывность» - суставная щель. Каждый сустав имеет следующие анатомические элементы: суставные поверхности костей, покрытые суставным хрящом, суставную капсулу, суставную полость, синовиальную жидкость (рис. 18). Суставные поверхности покрыты, как правило, гиалиновым хрящом (рис.19), лишь у височно-нижнечелюстного и грудино-ключичного суставов он волокнистый. Толщина хряща колеблется от 0,2 до 6,0 мм и находится в прямой зависимости, от функциональной нагрузки, испытываемой суставом. Чем больше нагрузка, тем толще хрящ.
Суставной хрящ лишен кровеносных сосудов и надхрящницы. Он состоит на 75 - 80 % из воды, и 20 - 25 % массы приходится на сухое вещество, около половины которого составляет коллаген, соединенный с протеогликанами. Первый придает хрящу прочность, вторые - упругость. Через межклеточное вещество путем диффузии из синовиальной жидкости в хрящ свободно проникают вода, питательные вещества и т. д., оно непроницаемо для, крупных молекул белка. Наряду с этим питание хряща осуществляется за счет диффузии из капилляров подлежащей субхондральной костной пластинки. В хряще имеются микроциркуляторные пути, расположенные в основном веществе между коллагеновыми пучками по ходу макромолекул протеогликанов.
Хрящ отделен от кости извилистой электронно-плотной линией, образующей множество выпячиваний, направленных в сторону хряща, в которые проникают синусоидные кровеносные капилляры. При этом в норме всегда между хрящом и капиллярами кости находятся пластинки остеоидной ткани. В суставном хряще различают три зоны: поверхностную, промежуточную и базальную(рис. 25). Непосредственно к кости прилежит слой хряща, пропитанный солями кальция. Над ним в основном веществе располагаются изогенные группы клеток - хондроцитов, залегающих в своих ячейках. Изогенные группы располагаются в виде колонок, перпендикулярных поверхности хряща. Над слоем изогенных групп находится тонкий волокнистый слой, а над ним - поверхностный слой. Со стороны суставной полости хрящ покрыт слоем аморфного вещества. Хондроциты секретируют гигантские молекулы, которые образуют межклеточное вещество.
Суставной хрящ защищает суставные концы кости от механических воздействий, уменьшая давление и равномерно распределяя его по поверхности. Деформация хряща, возникающая при движениях в суставе, обратима.
Суставная капсула (capsula articularis), прикрепляющаяся вблизи краев суставных поверхностей сочленяющихся костей или на некотором расстоянии от них, прочно срастается с надкостницей, образуя замкнутую суставную полость. Капсула состоит из двух слоев. Наружный слой представлен толстой прочной фиброзной мембраной, образованной волокнистой соединительной тканью, коллагеновые волокна которой направлены преимущественно продольно. Местами фиброзная мембрана образует утолщения - связки, укрепляющие суставную капсулу, связки могут располагаться также в толще капсулы (капсульные связки) или вне ее (внекапсульные связки). Кроме того, встречаются внутрикапсульные связки, расположенные в глубине сустава, покрытые снаружи синовиальной мембраной. Толщина и форма связок зависят от особенностей строения сустава и действующей на него силы тяжести. Связки не только укрепляют сустав, но и направляют, а также ограничивают движения. Они чрезвычайно прочны. Так, например, прочность на разрыв подвздошно-бедренной связки достигает 350 кг, а длинной связки подошвы - 200 кг. Связки выполняют также функцию пассивных тормозов, ограничивая движения в суставе. Связки вплетаются в надкостницу в зоне прикрепления суставной капсулы к кости.
Подобно надкостнице, суставная капсула богата сосудами и нервными окончаниями, особенно пластинчатыми тельцами (Фатера-Пачини) и колбами Краузе. Нервные окончания проникают в синовиальный слой.
Внутренний слой суставной капсулы образован тонкой гладкой блестящей синовиальной мембраной, которая выстилает изнутри фиброзную мембрану и продолжается на поверхность кости, не покрытой суставным хрящом. Синовиальная мембрана состоит из плоской и ворсинчатой частей. Последняя имеет множество небольших выростов, обращенных в полость сустава - синовиальные ворсинки, очень богатые кровеносными сосудами. Разветвленные ворсинки значительно увеличивают поверхность синовиальной мембраны. Количество ворсинок и складок синовиальной оболочки прямо пропорционально степени подвижности сустава (И. П. Каллистов, 1951). Через ворсинки осуществляются ультрафильтрация из кровеносного русла в полость сустава и резорбция жидкости из нее. Лимфатические сосуды не проникают в ворсинки, они образуют в плоской части синовиальной мембраны сплетение, начинающееся расширенными лимфатическими капиллярами - лакунами. Если сочленяющиеся поверхности не очень соответствуют друг другу (инконгруэнтны), синовиальная мембрана обычно образует различной величины и формы синовиальные складки.
В наиболее крупных складках, например в коленном суставе, имеются скопления жировой ткани. Синовиальная мембрана состоит из синовиальной собственной пластинки, образованной ретикулярными и коллагеновыми фибриллами, на которых располагается слой синовиальных клеток синовиоцитов. Синовиальные клетки лежат на тонком прерывистом слое основного вещества, похожего на базальную мембрану, но не являющегося ею.
Различают два типа синовиоцитов (В.Н. Павлова, 1980): секреторные и фагоцитарные. Секреторные синовиоциты вырабатывают синовиальную жидкость и ее главный компонент гиалуроновую кислоту. Синовиальная жидкость содержит 95 % воды, остальная часть - белки, мукополисахариды и соли. Фагоцитарные синовиоциты обладают свойствами макрофагов, они имеют множество цитоплазматических отростков и микроворсинок, богаты лизосомами и фагосомами.
Бесклеточная пластинка поверхностной зоны суставного хряща покрыта протекторной пленкой синовии, толщина которой меняется в зависимости от нагрузки на сустав. Сuновиальная жидкость смачивает покрытые хрящом суставные поверхности; устраняет их трение друг о друга и осуществляет трофику суставного хряща. Количество и состав смазочного вещества - синовии - зависит от функциональной нагрузки на сустав. Даже в таких крупных суставах, как коленный или тазобедренный, ее количество не превышает 2-4 мл. Давление в полости сустава ниже атмосферного. Прозрачная синовия содержит воду, соли, гиалуроновую кислоту, глюкозу, мочевину. Содержание белка в синовиальной жидкости значительно колеблется в различных суставах.
В норме у живого человека суставная полость представляет собой узкую щель, расположенную между покрытыми хрящом суставными поверхностями и ограниченную синовиальной мембраной. Форма суставной полости зависит от формы сочленяющихся поверхностей, наличия или отсутствия внутри сустава вспомогательных образований (синовиальные складки, суставной диск или мениск) либо внутрикапсульных связок.
Суставные поверхности редко полностью соответствуют друг другу по форме. Для достижения конгруэнтности (от лат. соngruens - согласный между собою, соответствующий) в суставах имеется ряд вспомогательных образований - хрящевых дисков, менисков, губ (рис. 20). Так, например, в височно-нижнечелюстном суставе имеется хрящевой диск, сращенный с капсулой по наружному краю. Диск, как правило, разделяет суставную полость на два этажа. Мениски - это несплошные хрящевые или соединительнотканные пластинки полулунной формы, расположенные между суставными поверхностями. В коленном суставе имеются полукольцевые медиальный и латеральный мениски, которые расположены между суставными поверхностями бедренной и большеберцовой костей.
Диски и мениски состоят из волокнистого (коллагенового) хряща, образованного упорядоченными пучками коллагеновых волокон, которые ориентированы соответственно направлению сил натяжения и давления. Диски и мениски способны смещаться при движениях. Они как бы сглаживают неровности сочленяющихся поверхностей, делают их конгруэнтными, амортизируют сотрясения и толчки при передвижении.
Суставная губа, расположенная по краю вогнутой суставной поверхности, дополняет и углубляет ее. Она прикреплена своим основанием к краю суставной поверхности, а внутренней вогнутой поверхностью обращена в сторону полости сустава. Так, по краю полулунной суставной поверхности вертлужной впадины имеется вертлужная губа, благодаря которой суставная поверхность тазобедренного сустава углубляется и больше соответствует шаровидной головке бедренной кости. К вспомогательным образованиям суставов относятся и синовиальные сумки, синовиальные влагалища - небольшие полости, образованные синовиальной мембраной и заполненные синовиальной жидкостью.
В зависимости от количества суставных поверхностей, участвующих в образовании сустава и их взаимоотношений между собой, суставы делятся на простые (две суставные поверхности), сложные (более двух), комбинированные и комплексные. Если два (или более) анатомически самостоятельных сустава функционируют совместно, то они называются комбинированными, например оба височно-нижнечелюстных сустава.
Комплексные - это суставы, в которых между сочленяющимися поверхностями имеются диск или мениск, разделяющие полость сустава на два отдела. Движения в суставах совершаются вокруг фронтальной, сагиттальной и продольной осей вращения. Вокруг фронтальной оси выполняются сгибание и разгибание, при которых угол между сочленяющимися костями уменьшается или увеличивается (например, при сгибании в локтевом суставе угол между плечом и предплечьем уменьшается, а при разгибании увеличивается до 180 град., конечность выпрямляется).
Вокруг сагиттальной оси осуществляются приведение, при котором одна из сочленяющихся костей (конечность) приближается к срединной плоскости (к туловищу), и отведение, при котором кость (конечность) удаляется от нее. При вращении кость поворачивается вокруг своей продольной оси в ту или иную сторону. Круговое движение - это последовательное движение вокруг всех осей, при котором свободный конец движущейся кости или конечности, например кисть руки, описывает окружность. Чем больше разность угловых величин (в угловых градусах) сочленяющихся поверхностей, тем больше размах (объем) движений. При почти равной протяженности суставных поверхностей объем движений в суставах незначителен. На величину объема движений в суставах влияют также количество и расположение связок, укрепляющих сустав, положение и степень растяжимости мышц, окружающих сустав.
Форма сочленяющихся поверхностей определяет количество осей, вокруг которых может совершаться движение. В зависимости от этого суставы делятся на одно-, двух- и многоосные (рис. 21).
Для удобства форму суставной поверхности сравнивают с отрезком тела вращения. При этом каждая форма сустава допускает то или иное число осей движения. Так, суставы цилиндрической и блоковидной формы являются одноосными. При, вращении прямой линии вокруг параллельной ей прямой оси возникает цилиндрическое тело вращения. Цилиндрические суставы - это срединный атлантоосевой, проксимальный лучелоктевой. Блок представляет собой цилиндр с бороздой или гребнем, расположенными перпендикулярно оси цилиндра, на другой суставной поверхности имеется соответствующее углубление или выступ. Примерами блоковидных суставов являются межфаланговые суставы кисти. Разновидностью блоковидных суставов является винтообразный сустав. Отличие винта от блока в том, что борозда расположена не перпендикулярно оси, а по спирали. Примером винтообразного сустава может служить плечелоктевой сустав.
Эллипсовидный, мыщелковый и седловидный суставы являются двухосными. При вращении половины эллипса вокруг его диаметра образуется тело вращения - эллипс. Лучезапястный сустав является эллипсовидным. Мыщелковый сустав по форме близок к блоковидному и эллипсовидному, его суставная головка – подобие эллипса, однако в отличие от первого суставная поверхность располагается на мыщелке. Например, коленный и атлантозатылочный суставы являются мыщелковыми (первый является также комплексным, второй - комбинированным).
Суставные поверхности седловидного сустава по форме напоминают два «седла» с пересекающимися под прямым углом осями. Седловидным является запястно-пястный сустав большого пальца, который характерен только для человека и обусловливает противопоставление большого пальца кисти остальным. Преобразование сустава в типично седловидный связано с трудовой деятельностью.
Шаровидный и плоский суставы - многоосные. При вращении половины круга вокруг его диаметра образуется шар. Кроме движения вокруг трех осей в этих суставах совершается еще и круговое движение (например, плечевой и тазобедренный суставы). Последний считают чашеобразным благодаря значительной глубине суставной ямки.
Плоские суставы также относятся к многоосным. Движения в них, хотя и могут производиться вокруг трех осей, отличаются малым объемом. Объем движения в любом суставе зависит от его строения, разности угловых размеров суставных поверхностей, а в плоских суставах величина дуги движения незначительна. К плоским относятся, например, межзапястные, предплюснеплюсневые суставы.
В раннем детском возрасте суставы развиваются интенсивно, окончательное Формирование всех элементов суставов заканчивается в возрасте 13 - 16 лет. Подвижность суставов больше у детей и молодых людей, у женщин она больше, чем у мужчин. С возрастом подвижность суставов уменьшается. Это связано со склерозированием фиброзной мембраны и связок, ослаблением мышечной активности. Лучшее средство для достижения высокой подвижности суставов и профилактики возрастных изменений – это постоянные физические упражнения.
Соединения костей черепа
Соединения костей черепа - это преимущественно фиброзные соединения (швы). В области лицевого черепа швы ровные, гладкие, плоские (гармоничные); в случае мозгового черепа швы зубчатые, между теменной костью и чешуей височной кости - чешуйчатый шов. В основании черепа у ребенка имеются синхондрозы, например клиновидно-затылочный, клиновидно-каменистый, каменисто-затылочный, которые с возрастом окостеневают, превращаясь в синостозы. Лишь нижняя челюсть образует с черепом синовиальное соединение - височно-нижнечелюстной сустав, подкрепленный связками.
Позвоночный столб образует с черепом атлантозатылочный, срединный и латеральные атлантоосевые суставы. Суставы укреплены связками.
Соединения костей туловища
Жизненная необходимость в высокой подвижности головы способствовала формированию между черепом, I и II шейными позвонками комплекса прочных и подвижных соединений. Между мыщелками затылочной кости и латеральными массами атланта образуется парный атлантозатылочный сустав. Капсула его прикрепляется по краю суставных поверхностей. По форме сустав эллипсовидный. Движения происходят по двум осям: вокруг фронтальной оси - наклоны головы вперед и назад; вокруг сагиттальной оси - наклоны головы в стороны. Между дугами атланта и затылочной костью проходят передняя и задняя атлантозатылочные мембраны.
Соединения между I и II шейными позвонками представлены срединным (непарным) и латеральным (парным) атлантоосевыми суставами (см. рис. 22). Срединный атлантоосевой сустав образован ямкой зуба на передней дуге атланта и зубом осевого позвонка. Суставная капсула прикрепляется по краю суставных поверхностей. По форме сустав цилиндрический, по функции - вращательный. Ось движения проходит вертикально через зуб осевого позвонка.
Латеральный атлантоосевой (парный) сустав расположен между нижними, суставными поверхностями латеральных масс атланта и верхними сочленовными поверхностями осевого позвонка. Суставная капсула прикрепляется по краю суставных поверхностей. Обширная капсула обеспечивает значительную подвижность сустава. По форме латеральный атлатоосевой сустав плоский. В целом во всех трех атлантоосевых суставах возможно движение вокруг одной оси, проходящей вертикально через зуб осевого позвонка (повороты головы в стороны). Суставы между затылочной костью, I и II шейными позвонками подкрепляются крыловидными связками, крестообразной связкой атланта и связкой верхушки зуба осевого позвонка.
Позвонки связаны между собой с помощью различного вида соединений. Между телами позвонков имеются межпозвоночные диски, толщина которых в грудном отделе 3-4 мм, в шейном - 5-6 мм, в поясничном - 10-12 мм. Диск состоит из расположенного в центре студенистого ядра (остаток хорды), близкого по своему строению гиалиновому хрящу, окруженному фиброзным кольцом, образованным волокнистым хрящом.
Благодаря такому строению диски прочные, упругие, а соединенные ими позвонки обладают некоторой подвижностью. В поясничном отделе позвоночника, который является наиболее подвижным, в межпозвоночных дисках нередко появляется полость, в результате чего образуются межпозвоночные симфизы.
Передняя и задняя продольные связки, образованные плотной волокнистой оформленной соединительной тканью, укрепляют соединения тел позвонков. Передняя связка проходит по передней поверхности тел позвонков, прочно срастается с межпозвоночными дисками, начиная от глоточного бугорка затылочной кости и переднего бугорка передней дуги атланта до 2 - 3-й поперечных линий тазовой поверхности крестца. Задняя продольная связка проходит вдоль задней поверхности тел позвонков в позвоночном канале, начиная от II шейного до I копчикового позвонка.
Суставные отростки позвонков соединены между собой и образуют дугоотросчатые суставы, остистые укреплены межостистыми и надостистой связками. Верхняя часть последней, натянутая между наружным гребнем затылочной кости и остистыми отростками шейных позвонков, называется выйной связкой. У человека в связи с прямохождением она утратила свою важную роль в поддержании головы. Поперечные отростки соединены между собой межпоперечными связками. Крестец соединен с копчиком с помощью межпозвоночного диска, а также ряда связок.
Длина позвоночного столба новорожденного ребенка составляет 40 % длины его тела. В первые 2 года позвоночник вырастает почти в 2 раза, до 15-1б лет рост замедленный, после 15-16 лет снова ускоряется. Развитие позвоночника завершается примерно к 23-25 годам. У взрослого человека позвоночный столб примерно в 3,5 раза длиннее (60-70 см), чем у новорожденного. Часть позвоночника, расположенная над крестцом, постепенно утолщается книзу, крестцово-копчиковая часть резко утончается. Число свободных (докрестцовых) позвонков у человека, как правило, 24 (в 5-8 % случаев 23 или 25). Число позвонков в крестцовом и копчиковом отделах также варьирует: крестец может состоять из 4 - 6 позвонков, копчик из 1 - 5 позвонков.
Межпозвоночные диски у детей относительно толще, чем у взрослых людей, с возрастом их толщина уменьшается, они становятся менее эластичными, студенистое ядро уменьшается в размерах, периферическая зона фиброзного кольца частично замещается хрящом и даже окостеневает.
Позвоночник человека имеет изгибы. Выпуклости позвоночного столба, обращенные вперед, называются лордозами, назад - кифозами. Шейный лордоз переходит в грудной кифоз, который в свою очередь сменяется поясничным лордозом, а затем крестцово-копчиковым кифозом. Функциональная роль изгибов очень велика. Благодаря ним удары, толчки и сотрясения, передающиеся позвоночнику при различных движениях, падении, ослабляются и не достигают черепа и, главное, мозга.
Движение позвоночника осуществляется вокруг трех осей: 1) поперечной оси - сгибание позвоночника вперед (флексия) и разгибание назад (экстензия); амплитуда этих движений 170 - 245°; 2) сагиттальной оси - боковое сгибание вправо и влево, общий размах движений около 165°; 3) продольной оси (вертикальной) - вращательные движения (ротация), общий размах - около 120° и круговое движение.
В шейном и поясничном отделах размах движений наибольший. Объем движений в шейном отделе: сгибание 70-79°, разгибание 95-105°, вращение 80-85°.
В грудном отделе подвижность позвоночника ограничена наличием ребер и грудины, тонкостью межпозвоночных дисков и частично направленными вертикально остистыми отростками. Сгибание, разгибание и боковые сгибания здесь невелики: сгибание до 35°, разгибание - до 50°, вращение - до 20°. В поясничном отделе толстые межпозвоночные диски способствуют большей подвижности (сгибание - до 60°, разгибание - до 45-50°), строение суставных отростков задерживает ротацию и боковые движения.
Наклоны позвоночника вправо и влево из исходного вертикального положения происходят при одновременном сокращении мышц-сгибателей и мышц-разгибателей туловища, а также квадратной мышцы поясницы соответствующей стороны (около55°).
Вращение (скручивание) позвоночника вокруг вертикальной оси (при стоянии на 90°, сидя - на 54°) осуществляют мышцы: поперечно-остистая, наружная косая - живота, лестничные (соответствующей стороны), внутренняя косая - живота, ременные - головы и шеи (противоположной стороны).
Разгибание выполняют мышцы: выпрямляющая позвоночник, трапециевидная, ременные головы и шеи. Сгибание - мышцы: прямые - живота, наружные и внутренние косые - живота, лестничные, длинные шеи, грудино-ключично-сосцевидные.
Грудная клетка. Грудная клетка образована соединенными между собой грудными позвонками, ребрами и грудиной. Ребра сочленяются с позвонками с помощью реберно-позвоночных суставов. С грудиной хрящи II - VII ребер соединяются с помощью суставов. Хрящи VIII - Х ребер между собой и с хрящами вышележащих ребер соединяются с помощью межхрящевых суставов и образуют реберную дугу. Благодаря таким соединениям составляющих ее костей грудная клетка обладает подвижностью. При вдохе и выдохе происходит вращение задних концов ребер в реберно-позвоночных суставах, одновременно смещаются и ребра, и грудина. При вдохе передние концы ребер и грудина поднимаются, межреберные промежутки расширяются, размеры грудной полости увеличиваются. При выдохе происходит опускание ребер и грудины, уменьшение межреберных промежутков и объема грудной полости.
Поднимание ребер осуществляют мышцы: наружные межреберные, поднимающие ребра, задние верхние зубчатые, лестничные. Опускание ребер - мышцы: внутренние межреберные, подреберные, поперечная мышца груди, нижние задние зубчатые.
Соединения ребер с позвонками (рис. 23) представлены суставами головок ребер, реберно-поперечными суставами и связками. Сустав головки ребра образован двумя реберными ямками (полуямками) двух смежных грудных позвонков и головкой ребра. Головки I, II и ХII ребер соединяются с полной ямкой на теле соответствующего позвонка. Суставы подкреплены внутрисуставными связками головок ребер (у I, II и ХII ребер их нет) и лучистой связкой головки ребра, расположенной снаружи.
Сустав бугорка ребра, или реберно-nоnеречный сустав, образован реберной ямкой поперечного отростка и суставной поверхностью бугорка ребра (у ХI и ХII ребер этого сустава нет). Сустав укреплен прочными связками. Пространство между шейкой ребра и поперечным отростком заполняет реберно-поперечная связка, а латерально - латеральная реберно-поперечная связка.
Сустав головки ребра и реберно-поперечный сустав анатомически разобщены, но движения в них происходят одновременно, поэтому функционально они представляют единый комбинированный сустав. По форме он цилиндрический. Вращение ребра осуществляется вокруг одной оси, проходящей вдоль его шейки. При этих вращательных движениях передние концы ребер вместе с грудиной совершают экскурсии вверх и вниз.
С грудиной соединяются только 7 верхних пар истинных ребер. Контактируют передние концы хрящевых частей ребер и реберные вырезки грудины. Хрящ I ребра соединяется с грудиной по типу непрерывного, постоянного хрящевого соединения. Хрящи VI и VII ребер чаще образуют с грудиной суставы, но могут присоединяться и по типу непрерывного хрящевого соединения. Хрящи II, III, IV, V ребер образуют с грудиной суставы. При этом сустав хряща IIребра разделяется внутрисуставной связкой на две полости: одна служит для сочленения с рукояткой грудины, другая - с ее телом. Суставные капсулы у этих четырех ребер не имеют той мощности и послойной структуры, которая характерна для обычных суставов. Она больше напоминает структуру надхрящницы, переходящей с хряща на кость.
Соединения ребер с грудиной укрепляются nередними и задними лучцстымц связками (ligg. sternocostalia radiata), пучки которых веером расходятся от хрящевых частей ребер на переднюю и заднюю поверхности грудины и образуют здесь мембраны (membranae sterni anterior et posterior).
Хрящи VIII, IX и Х ребер присоединяются к хрящу вышележащего ребра, образуя межхрящевые суставы (artt. interchondrales). Ребра ХI и ХIIсвободно расположены в толще стенки туловища.
Грудная клетка имеет четыре стенки. Передняя стенка образована грудиной и реберными хрящами, боковые - ребрами, задняя - грудными позвонками и задними концами ребер.
Грудная клетка человека по форме напоминает неправильной формы бочку, она расширена в поперечном направлении и уплощена в переднезаднем. Через верхнюю апертуру грудной клетки, которая ограничена первым грудным позвонком сзади, первой парой ребер и верхним краем грудины спереди, проходят трахея, пищевод, крупные кровеносные и лимфатические сосуды, нервы. Нижняя апертура грудной клетки, ограниченная ХII грудным позвонком, нижними ребрами, реберными хрящами и нижним концом грудины, закрыта диафрагмой. Грудная клетка спереди несколько короче, чем сзади, а грудина короче грудного отдела позвоночника. Последний находится на границе, разделяющей грудную полость на правую и левую половины, в которых помещаются легкие. С обеих сторон от позвоночника расположены вертикально ориентированные углубления - легочные борозды, в которых находятся задние края легких. Промежутки между двумя смежными ребрами на всем протяжении между позвоночником и грудиной называются межреберными пространствами (промежутками), в них залегают межреберные мышцы и связки, сосуды и нервы.
У человеческого плода грудная клетка сжата с боков, переднезадний размер больше поперечного, у новорожденного ребенка она по форме напоминает колокол. На первом году жизни поперечный размер несколько увеличивается. До 7-летнего возраста грудная клетка удлиненная. К 15 годам ее поперечный размер резко увеличивается, и она медленно растет, достигая окончательной формы к 17 - 20 годам. У стариков она уплощена в переднезаднем направлении, удлинена; у женщин короче, чем у мужчин.
Грудная клетка взрослого человека может изменяться под влиянием условий труда, физкультуры и спорта.
Соединения костей верхней конечности
Кости пояса верхней конечности связаны между собой и с грудиной с помощью суставов. Суставы образуются при соединении костей свободной верхней конечности между собой, а также с лопаткой. Кроме того, диафизы костей предплечья соединяются между собой межкостной перепонкой предплечья. Над ее верхним краем проходит фиброзный пучок - косая хорда.
Грудино-ключичный сустав (art. sternoclavicularis; рис. 20Б). Функциональное биомеханическое значение этого сустава велико: он является единственным суставом, соединяющим скелет верхней конечности со скелетом туловища. Сочленяются ключичная вырезка на рукоятке грудины и грудной конец ключицы. Внутри сустава имеется хрящевой диск (discus articularis), который разделяет полость сустава и устраняет инконгруэнтность (несоответствие по форме) суставных поверхностей ключицы и грудины. Довольно мощная суставная капсула прикрепляется по краю суставных поверхностей и укреплена несколькими связками. Сверху, перекидываясь над яремной вырезкой рукоятки грудины, идет поперечно от одной ключицы к другой межключичная связка (lig. inferсlaviculare). Хорошо укрепляют сустав передняя и задняя грудино-ключичные связки (ligg. sternoclaviculare anterius et posterius). Каждая из них идет от ключицы веером на переднюю или заднюю поверхность рукоятки грудины. Очень мощная связка, удерживающая ключицу в этом суставе, проходит в пространстве между I ребром и грудинным, концом ключицы - ре6ерно-ключuчная связка (lig. costoclaviculare). Форма суставных поверхностей на грудине и ключице приближается к седловидной.
Кроме того, суставной диск несколько увеличивает объем движений. В целом в грудино-ключичном суставе движения происходят вокруг трех осей: сагиттальной - поднимание и опускание плечевого пояса; вертикальной - движение плечевого пояса вперед-назад; фронтальной - вращение ключицы вокруг своей оси, при этом также смещаются весь плечевой пояс и верхняя конечность.
Aкромиально-ключичный сустав (art. acromioclavicularis). Этот сустав образован суставными поверхностями на акромиальном конце ключицы и акромиальном отростке лопатки. Суставные поверхности на обеих костях скошены таким образом, что акромиальный конец ключицы лежит на акромиальном отростке лопатки. По размерам суставные поверхности небольшие, ровные. По форме сустав плоский, объем движений незначителен, Сам сустав механически не прочен, поэтому он укрепляется тремя связками. Непосредственное отношение к суставу имеет акромиально-ключичная связка (lig. acromioclaviculare), соединяющая обе эти кости и имеющая верхние и нижние пучки. Особо прочная клювовидно-ключичная связка (lig. coracoclaviculare), которая имеет две части. Латеральная часть - трапециевидная связка (lig. trapezoideum) идет от трапециевидной линии ключицы к клювовидному отростку, а медиальная - коническая связка (lig. conoideum) соединяет клювовидный отросток с конусовидным бугорком ключицы.
Лопатка имеет свои собственные связки. Между акромионом и клювовидным отростком широкой лентой натянута клювовидно-акромиальная связка (lig. coracoacromiale). Она защищает плечевой сустав сверху. Через вырезку лопатки перекидывается короткая верхняя поперечная связка лопатки, превращающая вырезку в отверстие.
СОЕДИНЕНИЯ СВОБОДНОЙ ВЕРХНЕЙ КОНЕЧНОСТИ
Плечевой сустав (art. humeri; рис. 24). Особенностью верхней конечности человека является ее большая подвижность, которая необходима в трудовой деятельности. Это разнообразие движений обеспечивается несколькими факторами: единственными очень подвижным суставом, соединяющим всю верхнюю конечность с туловищем, - грудино-ключичным, а также весьма подвижным плечевым суставом, соединяющим свободную верхнюю конечность с ее поясом. Этот сустав имеет несколько особенностей. Прежде всего, это суставные поверхности. На лопатке суставная поверхность овальная, мелкая по глубине и маленькая, по отношению к сочленяющейся с ней головкой плечевой кости. Это несоответствие несколько устраняется тем, что по краю суставной впадины лопатки идет треугольной формы хрящевая лента - суставная гу6а (labrum glenоidаlе). Она увеличивает суставную поверхность ямки, создает большее сцепление плечевой кости с лопаткой и вместе с тем не уменьшает объема движений плечевой кости. Суставная поверхность плечевой кости обширна, шаровидная. Она расположена на головке; которая у человека наклонена медиально по отношению к оси плечевой кости, что создает дополнительные возможности для увеличения экскурсий свободного отдела верхней конечности.
Рис. 24 Правый плечевой сустав (вид спереди)
Капсула сустава (capsula articularis) прикрепляется на лопатке кнаружи от суставной губы, а на плечевой кости - по анатомической шейке.
Синовиальный слой ее образует длинное цилиндрическое выпячивание, сопровождающее сухожилие длинной головки двуглавой мышцы плеча.
Единственная, слабая связка плечевого сустава (lig. coracohumerale) идет от основания клювовидного отростка лопатки и вплетается в капсулу в направлении большого бугорка плечевой кости. Эта связка не ограничивает свободы движения плечевой кости и только подстраховывает ее от чрезмерного разгибания.
Сустав по форме типичный шаровидный. Имеет 3 оси движения. Вокруг фронтальной оси происходит сгибание - рука свободно поднимается до горизонтального положения. Обратное движение - разгибание - имеет меньшую амплитуду. Вокруг вертикальной оси верхняя конечность вращается внутрь и кнаружи. Вокруг сагиттальной оси осуществляются отведение и приведение. Только за счет плечевого сустава отведение возможно до горизонтального положения руки. Это определяется кинематическим соотношением суставной поверхности лопатки и бугорков плечевой кости. Движение верхней конечности выше горизонтали сопровождается поворотом всего плечевого пояса вокруг сагиттальной оси (рис. 25).
Рис. 25 Отведение плеча
Высокая подвижность сустава, его свободная капсула, малая площадь сочленения с лопаткой и слабая связка являются причинами возможных вывихов плечевого сустава, особенно при падениях. Несколько защищает сустав от вывиха кверху клювовидно-акромиальная связка. Она образует над суставом свод.
Поднимание лопатки и ключицы выполняют мышцы: поднимающая лопатку, ромбовидная, грудино-ключично-сосцевидная, трапециевидная (верхние пучки).
Опускание лопатки и ключицы - мышцы: трапециевидная (нижние пучки), передняя зубчатая (нижние пучки), малая грудная, подключичная.
Движение вперед и в латеральную сторону - мышцы: передняя зубчатая, малая грудная.
Движения лопатки кзади и в медиальную сторону (к позвоночнику) - мышцы: трапециевидная, ромбовидная.
Вращение лопатки вокруг сагиттальной оси: поворот нижним углом кнаружи - мышцы: передняя зубчатая (нижние зубцы), трапециевидная (верхние пучки).
Поворот лопатки нижним углом медиально (к позвоночнику) - мышцы: ромбовидная, малая грудная.
Сгибание плеча осуществляют мышцы: дельтовидная (часть, начинающаяся от ключицы), большая грудная (ключичная часть), клювовидно-плечевая, двуглавая м. плеча.
Разгибание плеча (размах сгибание - разгибание - 120°) - мышцы: дельтовидная (часть, начинающаяся от ости лопатки), широчайшая м. спины, большая круглая.
Отведение плеча - мышцы: дельтовидная (акромиальная часть), надостная, двуглавая м. плеча (длинная головка), дельтовидная (ключичная и часть, начинающаяся от ости лопатки при отведении более 60°).
Приведение плеча (отведение-приведение 100°) - мышцы: большая грудная, клювовидно-плечевая, широчайшая м. спины, большая и малая круглые, дельтовидная (ключичная и часть, начинающаяся от ости лопатки, если рука отведена менее чем на 60°).
Вращение вовнутрь - мышцы: дельтовидная (ключичная часть), большая грудная, подлопаточная, широчайшая м. спины, большая круглая.
Вращение кнаружи (поворот вокруг продольной оси - 135°), мышцы: дельтовидная (часть, начинающаяся от ости лопатки), надостная, подостная, малая круглая.
В круговых движениях, принимают участие все указанные выше мышцы.
Локтевой сустав (art. cubiti; рис. 26). Это сложное сочленение между плечевой костью и проксимальными концами лучевой и локтевой костей, объединенное общей капсулой и состоящее из трех отдельных суставов: плечелоктевого, плечелучевого и проксимального лучелоктевого.
Плечелоктевой сустав (art. humeroulnaris) образован блоком плечевой и блоковидной вырезками локтевой костей. По форме сустав блоковидный. Борозда на блоке плечевой кости имеет спиральный ход, поэтому при сгибании в локтевом суставе предплечье ложится не на плечо, а на большую грудную мышцу.
Плечелучевой сустав (art. humeroradialis) по форме шаровидный, но как шаровидный он не работает. Лучевая кость соединена с локтевой, а последняя прочно связана с плечевой, поэтому невозможно использовать сагиттальную ось, т. е. отведение и приведение в локтевом суставе невозможны.
В проксимальном лучелоктевом суставе (art. radioulnais proximalis) сочленяются суставная окружность головки лучевой кости и лучевая, вырезка локтевой кости. Этот сустав по форме цилиндрический, вращательный. Общая капсула прикрепляется на плечевой кости впереди выше венечной и локтевой ямок, сзади, почти доходит до верхнего края локтевой ямки, а с боков проходит ниже мыщелков. Фиксация капсулы с боков по краю суставной поверхности оставляетнадмыщелки свободными дляприкрепления мышц-сгибателей и разгибателей локтевого, лучезапястного суставов и суставов кисти. На локтевой кости капсула прикрепляется по краю суставных поверхностей, а на лучевой - по ее шейке. Сустав укреплен лучевой u локтевой коллатеральными связками (ligg. collaterale radiale et ulnаrе), идущими от надмыщелков плечевой кости к локтевой, а также кольцевой связкой лучевой кости, охватывающей шейку лучевой кости и прикрепляющейся к локтевой кости (рис. 27).
Рис. 27 Связки локтевого сустава
Движение в локтевом суставе происходит вокруг фронтальной оси - сгибание и разгибание предплечья. Вокруг вертикальной оси вращается лучевая кость, а вместе с ней и вся кисть. Положение, когда при опущенных руках ладони направлены кпереди, называется супинацией (supinatio); обратное состояние (ладонь направлена кзади) – пронацией (рrоnаtio).
Пространство между диафизами лучевой и локтевой костей занято прочной фиброзной межкостной перепонкой предплечья.
Дистальный лучелоктевой сустав (art. radioulnaris distalis). Образован суставной окружностью головки локтевой кости, локтевой вырезкой лучевой кости и хрящевым диском. Последний имеет форму треугольника, основанием прикрепляется к локтевой вырезке, а вершиной - к шиловидному отростку головки локтевой кости. Локтевая вырезка лучевой кости и суставной диск, вместе образуют суставную ямку для головки локтевой кости. Сустав по форме цилиндрический, вращательный, с одной вертикальной осью, вокруг которой происходят супинация и пронация предплечья.
В локтевом суставе происходит сгибание и разгибание предплечья (170°). При сгибании предплечье несколько отклонено в медиальную сторону, кисть ложится на грудь. При максимальном разгибании локтевой отросток упирается в ямку локтевого отростка плечевой кости и плечо с предплечьем находятся почти на одной прямой.
Сгибание осуществляют мышцы: двуглавая плеча, плечевая, плечелучевая, лучевой сгибатель запястья, длинная ладонная, длинный лучевой разгибатель запястья, короткий лучевой разгибатель запястья.
Разгибание - мышцы: трехглавая м. плеча, локтевая.
В плечелоктевом суставе возможны лишь сгибание и разгибание предплечья.
В плечелучевом - вращение лучевой кости (предплечья) вокруг продольной оси - пронация, супинация и сгибание.
В проксимальном лучелоктевом - вращение лучевой кости (предплечья) вокруг локтевой - пронация и супинация. При этом проксимальный эпифиз лучевой кости поворачивается на месте, так как его головка удерживается кольцевой связкой лучевой кости и у лучевой вырезки, а дистальный скользит по суставной окружности локтевой кости, описывая дугу вокруг неподвижной головки локтевой кости. Максимальный размах движения предплечья в лучелоктевых суставах (супинация и пронация) достигает 180°.
В дистальном лучелоктевом суставе супинацию (при пронированной лучевой кости) выполняют мышцы: супинатор, двуглавая м. плеча, плечелучевая, разгибатель указательного пальца, длинный и короткий разгибатели большого пальца кисти, длинная м., отводящая большой палец кисти (при полусогнутом локтевом суставе), двуглавая и плечелучевая.
Пронация (при супинированной лучевой. кости), мышцы: круглый и квадратный пронаторы, лучевой сгибатель кисти, длинная ладонная, длинный лучевой разгибатель запястья (смещение лучевой кости вокруг лучевой около 150°)
Соединения костей кисти
Лучезапястный сустав (art. radiocarpea) (рис. 28). Этот сустав соединяет кости предплечья с кистью. Суставную ямку в нем образуют запястная суставная поверхность лучевой кости и дистальная поверхность суставного диска. Суставной головкой служат кости проксимального ряда костей запястья. Суставная капсула укреплена лучевой и локтевой коллатеральными связками, а также связками, соединяющими на ладонной и тыльной сторонах отдельные кости запястья. Сустав по форме эллипсовидный. Вокруг фронтальной оси возможны сгибание и разгибание, вокруг сагиттальной оси - приведение и отведение кисти.
Сгибание кисти (около 80°) осуществляют мышцы: лучевой и локтевой сгибатели кисти, длинная ладонная, поверхностный и глубокий сгибатели пальцев (особенно при разогнутых пальцах). Разгибание кисти (около70°) - мышцы: локтевой разгибатель запястья, длинный и короткий лучевые разгибатели запястья, разгибатели пальцев (особенно при согнутых пальцах). Приведение кисти (около 40°) - мышцы: локтевые сгибатель и разгибатель кисти. Отведение кисти (около 15°), мышцы: лучевые сгибатели и разгибатели кисти. Возможно круговое движение кисти - осуществляют последовательно мышцы, сгибающие и разгибающие кисть (около150°)
Среднезапястный сустав (art. mediocarpae). Расположен сустав между проксимальным (за исключением гороховидной) и дистальным рядами костей запястья. Суставная щель идет S-образно. Укреплен теми же связками, что и лучезапястный. Функционально среднезапястный сустав объединяется с лучезапястным суставом, увеличивая объем движений, которые происходят вокруг тех же осей.
3апястно-пястные суставы (artt. carpometaсаrраеае). В суставе кости дистального ряда запястья соединяются с основаниями пяти пястных костей. Линия сустава идет в поперечном направлении. Капсула укрепляется ладонными и тыльными запястно-пястными связками. По форме эти суставы (за исключением сочленения I пальца) плоские, с незначительным объемом движений. 3апястно-пястный сустав I пальца (между костью-трапецией и I пястной) седловидный, очень подвижный. При движении вокруг его осей большой палец может приближаться к указательному или отводиться от него, а также противопоставляться мизинцу.
Рис. 28 Соединения костей кисти, правой; фронтальный распил.
Межпястные суставы (artt. intermetacarpales). Образованы боковыми суставными поверхностями оснований II-V пястных костей. Укрепляются тыльными и ладонными пястными связками. По форме эти суставы плоские, движения в них незначительны.
Пястно-фаланговые суставы (artt. metacarpophalangeaeae). Они находятся между головками пястных костей и основаниями проксимальных фаланг. Укрепляются коллатеральными связками. Эти суставы эллипсовидные; вокруг фронтальной оси в них осуществляются сгибание и разгибание, вокруг сагиттальной оси - отведение и приведение.
Межфаланговые суставы кисти (artt. interphalangeaeae). Суставы образованы головками и основаниями проксимальнее и дистальнее расположенных фаланг. Укрепляются боковыми связками. По форме межфаланговые суставы блоковидные: в них возможны сгибание и разгибание.
Функционально и клинически в суставах кисти выделяют твердую основу кисти. Это комплекс плотно соединенных многочисленными ладонными, тыльными и межкостными связками костей второго ряда запястья и четырех пястных костей. Плоские неровные их поверхности и мощный связочный аппарат не допускают обширных движений.
Соединения костей нижней конечности
Две тазовые кости пояса нижних конечностей, соединяясь между собой с помощью лобкового симфиза и с крестцом сзади, образуют таз (рис. 29).
Рис. 29 Соединения пояса нижней конечности (вид спереди).
Тазовая кость у детей и подростков представлена костными закладками трех составляющих ее костей: подвздошной, лобковой и седалищной, соединенных между собой хрящевыми прослойками. Далее этот временный синхондроз заменяется возрастным синостозом и кости срастаются в одно целое. У человека тазовая кость соединяется с крестцом, поясничным отделом позвоночника, тазовой костью противоположной стороны, имеет свои собственные связки и сочленяется со свободной нижней конечностью.
Крестцово-подвздошный сустав (art. sacroiliaca). Этот сустав образован ушковидными поверхностями крестца и подвздошной кости. Эти поверхности очень шероховаты и занимают почти половину длины боковой части крестца. Капсула прикрепляется по краю суставных поверхностей. Сустав укреплен самыми мощными крестцово-nодвздошными связками (ligg. sacroiliaca interossea), идущими многочисленными пучками между бугристостью крестца и подвздошной бугристостью. На вентральной и дорсальной поверхностях костей имеются соответственно вентральные и дорсальные крестцово-подвздошные связки. Лучшему сцеплению суставных поверхностей способствует и вентродорсальная их скошенность на крестце и подвздошной кости. Последняя почти черепицеобразно покрывает латеральные отделы крестца. Полость сустава имеет вид узкой щели. По форме сустав плоский, объем движений незначительный.
Между крестцом и тазовой костью расположены также крестцово-бугорная и крестцово-остистая связки, идущие от крестца к седалищному бугру и седалищной ости тазовой кости. Эти связки дополняют костные стенки таза в задненижнем его отделе, ограничивая большое и малое седалищные отверстия, через которые проходят мышцы, сосуды и нервы.
Между задним отделом гребня подвздошной кости и поперечным отростком V поясничного позвонка растянута nодвздошно-поясничная связка. Она дополняет сзади костную стенку большого таза, прикрывая собой свободное треугольное пространство между задними отделами крыльев подвздошной кости и нижним поясничным позвонком.
Лобковый симфиз (symphysis рubicа). Симфизиальные поверхности лобковых костей противоположных сторон, соединяясь посредством межлобкового диска, образуют лобковый симфиз. Внутри межлобкового диска, состоящего из волокнистого хряща, у взрослых людей находится узкая синовиальная полость. По верхнему краю лобковый симфиз укреплен верхней лобковой связкой, а по нижнему - дугообразной связкой лобка.
3апирательное отверстие тазовой кости закрывается запирательной мембраной, в верхнемедиальном отделе которой имеется отверстие - запирательный канал. Через него проходят сосуды и нервы. Наконец, тазовая кость сочленяется со свободным отделом, нижней конечности посредством тазобедренного сустава (рис 30).
Рис. 30 Связки тазобедренного сустава.
Тазобедренный сустав (art. сохае). Сочленяются полулунная суставная поверхность вертлужной впадины тазовой кости и головка бедренной кости. Ямка глубокая. Плоскость входа в нее наклонена латерально, поэтому верхний край вертлужной впадины нависает над ней, образуя хорошую опору для головки бедренной кости. Эта особенность связана с функцией нижней конечности как опоры.
По краю вертлужной впадины, дополняя и углубляя ее, проходит хрящевой ободок – вертлужная губа. Капсула сустава на тазовой кости прикрепляется по краю вертлужной впадины, на бедренной кости спереди доходит до межвертельной линии, а сзади фиксируется медиальнее межвертельного гребня, оставляя вертельную ямку вне полости сустава.
Четыре мощные наружные связки укрепляют сустав. Спереди, от передненижней ости подвздошной кости до межвертельной линии, идет наиболее прочная подвздошно-6едренная связка (lig. iliofemorale). Она помогает сохранять вертикальное положение и препятствует чрезмерному разгибанию в тазобедренном суставе. На медиально-нижней стороне сустава расположена лобково-бедренная связка (lig. рubоfemorale). Она проходит от верхней ветви лобковой кости к капсуле сустава и малому вертелу и ограничивает отведение бедра. 3адненижние отделы капсулы укрепляет седалищно-бедренная связка (lig. ischiofemorale), идущая от тела седалищной кости к задним отделам капсулы и ограничивающая вращение бедра внутрь. Под этими тремя связками в глубоких слоях капсулы расположена связка, называемая круговой зоной (zona orbicularis). Она петлей охватывает шейку бедренной кости и прикрепляется под нижней передней подвздошной остью.
Особенностью тазобедренного сустава является наличие двух внутрисуставных связок. Одна из них - поперечная связка вертлужной впадины (lig. transversum acetabuli) - перекидывается над вырезкой вертлужной впадины. Другая – связка головки бедренной кости (lig. capitis femoris) - начинается от краев вырезки вертлужной впадины и предыдущей связки и заканчивается на ямке головки бедренной кости. В ней проходят питающие головку бедренной кости кровеносные сосуды.
По форме тазобедренный сустав относится к шаровидным. Однако вследствие глубины ямки и более прочного соединения ее с головкой бедренной кости сустав считается разновидностью шаровидного - чашеобразным суставом (art. cotylica). Вокруг фронтальной оси возможны сгибание и разгибание, вокруг сагиттальной оси – отведение и приведение, вокруг вертикальной оси -вращение бедра внутрь и наружу. Возможно также сочетанное круговое движение, когда бедро и вся нижняя конечность описывают конус. Вследствие большой глубины вертлужной впадины движения в суставе не так обширны, но сустав более приспособлен к значительным нагрузкам.
Сгибание (до 120°, при согнутой в коленном суставе голени - 85-90°) осуществляют мышцы: прямая бедра, подвздошно-поясничная, напрягатель широкой фасции бедра, портняжная, средняя ягодичная (передняя часть), малая ягодичная (передняя часть), гребенчатая.
Разгибание (12-13°) - мышцы: большая ягодичная, большая приводящая, полуперепончатая, полусухожильная, средняя ягодичная (дорсальная часть), двуглавая м. бедра (длинная головка), квадратная м. бедра, малая ягодичная (дорсальная часть).
Отведение (до 40°) - мышцы: средняя и малая ягодичные, напрягатель широкой фасции бедра, большая ягодичная (верхняя часть), грушевидная, портняжная.
Приведение (до 20°) - мышцы: длинная, короткая и большая приводящие, большая ягодичная (нижняя часть), полуперепончатая, полусухожильная, подвздошно-поясничная, двуглавая м. бедра (длинная головка), гребенчатая, наружная запирательная, нежная.
Вращение кнутри (до 35°) - мышцы: напрягатель широкой фасции бедра, средняя и малая ягодичные (передние части), большая приводящая (пучки, прикрепляющиеся к медиальному надмыщелку бедра), подвздошно-поясничная.
Вращение кнаружи (до 150) - мышцы: большая ягодичная, средняя ягодичная (дорсальная часть), внутренняя и наружная запирательные, верхняя и нижняя близнецовые, подвздошно-поясничная, малая ягодичная (дорсальная часть), грушевидная, прямая м. бедра, длинная и короткая приводящие, гребенчатая, двуглавая м. бедра (длинная головка), квадратеная м. бедра, портняжная.
Коленный сустав (art. genus; рис. 31) образован бедренной, большеберцовой костями и надколенником. В связи с тем, что мыщелки бедренной кости выпуклые, а мыщелки большеберцовой кости только слегка вогнутые, возникает неконгруэнтность суставных поверхностей. Это несоответствие устраняется тем, что внутри коленного сустава находятся латеральный и медиальный мениски (meniscus medialis et lateralis). Их верхние поверхности конгруэнтны с мыщелками бедренной кости, а нижние с мыщелками большеберцовой кости.
Мениски серповидные, наружный край свободен. По внешнему виду они разные. Медиальный мениск имеет вид очень узкого серпа, кольцом охватывающего суставную поверхность медиального мыщелка. Латеральный мениск меньше по диаметру, с широким серповидным поясом, прикрывающим большую часть суставной поверхности латерального мыщелка большеберцовой кости. Своими концами мениски прикрепляются к межмыщелковому возвышению большеберцовой кости. Спереди они связаны поперечной связкой колена (lig. transversum genus).
Рис. 31 Левый коленный сустав (вид спереди).
Внутри коленного сустава также находятся передняя и задняя крестообразные связки (ligg. cruciatum anterius et posterius). Они укрепляют сустав, соединяя бедренную и большеберцовую кости.
Капсула сустава на бедренной кости прикрепляется высоко, сзади почти доходит до суставных поверхностей мыщелков, с боков она наиболее близка к суставной поверхности - проходит между мыщелками и надмыщелками. Последние остаются свободными. На большеберцовой кости капсула прикрепляется под суставной поверхностью мыщелков, а на надколеннике линия фиксации капсулы идет непосредственно по краю суставной поверхности.
Сустав укрепляется внешними связками. Большеберцовая коллатеральная связка (lig. collaterale tibiale) идет от медиального надмыщелка бедренной кости к медиальному мыщелку большеберцовой кости. Малоберцовая коллатеральная связка (lig. соllаterale fibulare) расположена между латеральным надмыщелком бедренной кости и головкой малоберцовой кости. Укрепляет сустав и связка надколенника (lig. patellae), прикрепляющаяся к бугристости большеберцовой кости и являющаяся частью сухожилия четырехглавой мышцы бедра. Сзади капсулу поддерживают косая и дугообразная подколенные связки (ligg. poplitea obliquum et arcuatum).
Необходимо отметить, что вокруг коленного сустава имеется много (более 10) синовиальных сумок. Часть их них сообщается с полостью сустава и способствует нормальному функционированию больших по площади суставных поверхностей. Другая часть сумок находится под сухожилиями мышц, уменьшая их трение о кость.
Коленный сустав блоковидно - вращательный. Вокруг фронтальной оси возможны сгибание и разгибание, вокруг вертикальной оси при согнутом коленном суставе - вращение голени внутрь и наружу.
Разгибание (до 180°) осуществляют мышцы: четырехглавая м. бедра, напрягатель широкой фасции бедра.
Сгибание (до 130°) - мышцы: полусухожильная, полуперепончатая, двуглавая м. бедра, нежная (тонкая), портняжная, икроножная, подколенная, подошвенная.
Вращение внутрь (до 10°) (только при согнутом коленном суставе) - мышцы: полуперепончатая, полусухожильная, подколенная, портняжная, икроножная (латеральная головка), нежная.
Вращение кнаружи (до 40°) (только при согнутом коленном суставе) - мышцы: двуглавая м. бедра, икроножная (наружная головка), напрягатель широкой фасции бедра.
Соединения костей голени (рис. 32). Проксимальные концы большеберцовой и малоберцовой костей образуют межберцовый сустав (art. tibiofibularis). Капсула его прикрепляется по краю суставных поверхностей и укрепляется передней и задней связками головки малоберцовой кости. Сустав плоский, возможны небольшие скользящие движения.
Пространство между телами костей заполнено межкостной перепонкой голени - прочной фиброзной пластинкой, прикрепляющейся к межкостным краям большеберцовой и малоберцовой костей. Дистальные концы костей голени соединяются посредством передней и задней межберцовых связок.
Рис. 32 Проксимальный межберцовый сустав (вид спереди).
Голеностопный сустав (аrt. talocruralis, рис. 33). Нижняя суставная поверхность большеберцовой кости и суставные поверхности мыщелков обеих костей голени образуют суставную ямку в виде вилки, охватывающей блок таранной кости. Суставная капсула тонкая, прикрепляется по краю суставных поверхностей, спереди несколько отступая от них. С внутренней стороны сустав укреплен медиальной связкой, идущей от медиальной лодыжки к таранной, пяточной и ладьевидной костям. С наружной стороны проходят передняя и задняя таранно-малоберцовые и пяточно-малоберцовая связки. Сустав блоковидный с одной фронтальной осью, вокруг которой возможны разгибание (поднимание носка стопы кверху) и сгибание (опускание стопы), при согнутой стопе - боковые движения.
Тыльное сгибание в суставе осуществляют мышцы: передняя большеберцовая, длинный разгибатель пальцев стопы, длинный разгибатель большого пальца стопы, третья малоберцовая.
Подошвенное сгибание (общий объем 60-70°) - мышцы: икроножная, камбаловидная, длинный сгибатель большого пальца стопы, задняя большеберцовая, длинный сгибатель пальцев стопы, длинная и короткая малоберцовые.
Семь костей предплюсны соединяются между собой межпредплюсневыми суставами подтаранным, таранно-пяточно-ладьевидным, пяточно-кубовидным и клиноладьевидным.
Подтаранный сустав (art. subtalaris). Сустав образуют задние суставные поверхности таранной и пяточной костей. Капсула проходит по краю суставных поверхностей и укрепляется боковыми таранно-пяточными связками. Сустав цилиндрический, функционирует вместе со следующим суставом.
Таранно-пяточно-ладьевидный сустав (art. talocalcaneonavicularis). Суставную впадину для головки таранной кости образуют передняя таранная суставная поверхность пяточной кости, впадина ладьевидной кости и подошвенная пяточно-ладьевидная связка, приобретающая в области сустава хрящевую структуру. Суставная капсула укреплена с тыльной и подошвенной сторон связками. Имеется также мощная межкостная связка между таранной и пяточной костями. Сустав шаровидный. Он функционирует обычно вместе с подтаранным суставом, поэтому движения происходят вокруг одной оси, идущей косо вперед, вверх и медиально. Приведение стопы сопровождается ее супинацией (поднимается медиальный край стопы), а отведение происходит вместе с пронацией (поднимается латеральный край стопы).
Пяточно-кубовидный сустав (art. calcaneocuboidea). Cycтaв образуют прилежащие суставные поверхности пяточной и кубовидной костей. Капсула сустава прикрепляется по краю суставных поверхностей, укреплена тыльной и подошвенной связками. По форме сустав седловидный, с одной продольной осью, допускающей скользящие вращательные движения.
Анатомически разобщенные, таранно-ладьевидный и пяточно-кубовидный суставы для целей практической медицины объединяются под общим названием «поперечный сустав предплюсны». В дополнение к описанным связкам сустав укрепляется раздвоенной связкой (lig. bifurcatum), состоящей из пяточно-кубовидной и пяточно-ладьевидной связок. Раздвоенная связка считается «ключом» Шопарова сустава: без ее рассечения сустав не раскроется.
Клиноладьеввдный сустав (art. cuneonavicularis) образован передней суставной поверхностью ладьевидной кости и задними суставными поверхностями трех клиновидных костей. Капсула прикрепляется по краю суставных поверхностей и дополняется тыльными, подошвенными и межкостными связками. По форме сустав плоский, со скользящими движениями, дополняющими движения в надтаранном и таранно-пяточно-ладьевидном суставах.
Предплюсне-плюсневые суставы (artt. tarsometatarsaeae). Различают три анатомически разобщенных сустава: 1) между медиальной клиновидной и I плюсневой костями; 2) между промежуточной и латеральной клиновидной с одной стороны и II и III плюсневыми костями - с другой; 3) между кубовидной и IV и V плюсневыми костями. Суставы укреплены тыльными, подошвенными и межкостными связками. По форме плоские, с незначительными скользящими движениями. Для практических целей эти три сустава объединяют в один поперечно идущий сустав.
Он также имеет свой «ключ» - медиальную межкостную предплюсне-плюсневую связку, идущую от медиальной клиновидной кости к основанию II плюсневой кости.
Плюснефаланговые суставы (artt. metatarsophaIangeae) образованы суставными поверхностями головок плюсневых костей и оснований проксимальных фаланг. Укреплены коллатеральными, глубокой поперечной плюсневой и подошвенными связками. В суставах возможны сгибание и разгибание пальцев, а также небольшие отведения и приведения.
Межфаланговые суставы стопы (artt. Interphalangeae pedis). Суставы расположены между фалангами пальцев стопы. Укреплены коллатеральными связками. По форме блоковидные, движения возможны вокруг одной поперечной оси - сгибание и разгибание пальцев стопы.
Стопа - орган опоры и передвижения - несет на себе всю тяжесть человеческого тела. Это накладывает существенный отпечаток на ее строение и характер соединения костей. Соединения костей стопы многочисленны. Наряду с сочленениями костей между собой они соединяются и с костями голени. Суставы костей предплюсны укреплены тыльными и подошвенными связками, межкостными связками, среди которых особую роль играет чрезвычайно прочная короткая внутрисуставная межкостная таранно-пяточная связка. Наибольшей прочностью обладает длинная подошвенная связка, которая перекидывается между нижней поверхностью пяточной кости и основаниями II - V плюсневой кости. У человека сводчатая стопа представлена пятью продольными и одним поперечным сводами (дугами), которые обращены выпуклостью кверху (рис. 34).
Своды образованы сочленяющимися между собой костями предплюсны и плюсны. Каждый продольный свод начинается от одной и той же точки пяточной кости - пяточного бугра - и включает кости предплюсны и соответствующую плюсневую кость. В образовании I свода - медиального - участвует и опора таранной кости. Стопа в целом имеет три точки опоры: пяточный бугор и головки I и V плюсневых костей. Продольные своды имеют неодинаковую высоту. Наиболее высокий II свод (вторая дуга). В результате формируется поперечный свод стопы. В его образовании принимают участие ладьевидная, клиновидные и кубовидная кости. Конструкция стопы в виде сводчатой арки у живого человека поддерживается благодаря форме костей, прочности связок (пассивные «затяжки» стопы) и тонусу мышц (активные «затяжки»). Для укрепления продольных сводов стопы наиболее важны длинная подошвенная связка, подошвенный апоневроз, подошвенная пяточно-ладьевидная связка, для поперечного свода - глубокая поперечная плюсневая и межкостные плюсневые связки.
Рис. 34 Своды стопы.
С помощью специальных упражнений можно добиться изумительной гибкости позвоночника, увеличить объем движений в суставах.
|
|
||||
|
|
|
||||
|
ТИП МЫШЕЧНОЙ ТКАНИ Мышечные ткани (textus, musculares; рис. 35) делятся на гладкую (неисчерченную) и поперечнополосатую (исчерченную). В свою очередь последняя состоит из скелетной и сердечной мышечных тканей. Основное свойство всех мышечных тканей – способность к сокращению, что лежит в основе всех двигательных процессов в организме. Сократительными элементами мышечных тканей являются миофибриллы.
Гладкая мышечная ткань (textusmuscularis nonstriatus; рис. 35А) входит в состав мышц, расположенных в стенках кровеносных сосудов и полых внутренних органов (желудок, кишечник, матка и т.д.) и сокращается непроизвольно, медленно и ритмично. Она состоит из веретенообразных мышечных клеток, или миоцитов, в центре которых обычно располагаются удлиненные палочковидные ядра. Миофибриллы гладких миоцитов локализуются по продольной оси. Гладкие мышечные клетки объединяются в пучки, последние - в мышечные пласты, которые образуют части стенок полых внутренних органов.
Поперечнополосатая скелетная мышечная ткань(textus muscularis striatus; рис. 35Б) образует мускулатуру скелета, мышц языка, глотки, диафрагмы ит.д. и сокращается произвольно, с высокой скоростью и большой силой. Структурно-функциональной единицей такой ткани является мышечное волокно, представляющее собой удлиненный многоядерный симпласт. Миофибриллы в мышечных волокнах расположены упорядоченно и состоят из регулярно повторяющихся фрагментов (саркомеров) с разными оптическими и физико-химическими свойствами, что обусловливает поперечную исчерченность всего волокна. В цитоплазме мышечных волокон содержится миоглобин, который окрашивает их в красный цвет. В связи с разным содержанием миоглобина в мышечных волокнах различают красные, белые и промежуточные волокна.
Сердечная мышечная ткань(textus muscularis cardiacus; рис. 35В) входит в состав мышечной стенки сердца (миокарда) и отличается тем, что состоит не из мышечных волокон, а из мышечных клеток - кардиомиоцитов. Различают рабочие, атипичные и секреторные кардиомиоциты.
Основной морфофункциональной единицей миокарда является рабочий кардиомиоцит с ядром, расположенным в его центральной части.
Миофибриллы располагаются на периферии кардиомиоцита, окружены многочисленными митохондриями (саркосомами) и могут переходить из одной клетки в другую, образуя сердечный функциональный синцитий. Кардиомиоциты соединяются друг с другом конец в конец при помощи особых соединений - вставочных дисков.
Атипичные проводящие кардиомиоциты входят в состав системы, проводящей возбуждение. Секреторные кардиомиоциты клеток предсердий вырабатывают некоторые гормоны.
МИОЛОГИЯ (учение о мышцах)
В организме человека аппарат движения представлен костями, их соединениями и скелетными поперечнополосатыми мышцами. Только мышцы являются активным звеном в динамической цепи движения, которое, действуя на костные рычаги, изменяет положение тела человека или его частей.
Сокращение скелетных мышц не только удерживает тело человека в вертикальном положении, обеспечивает его перемещение в пространстве, но и способствует улучшению крово- и лимфообращения, оказывает влияние на развитие и форму костей. скелетные мышцы осуществляют дыхательные и глотательные движения, мимику, участвуют в образовании стенок полостей тела.
В организме человека насчитывается 500 - 600 мышц. Масса их у мужчин составляет около 40 - 45% (у культуристов до70%), у женщин - 30% от массы тела.
Мышца (musculus) как орган имеет специфические форму и конструкцию и выполняет присущую только ей функцию. В состав мышц входят мышечная ткань, рыхлая и плотная соединительные ткани, сосуды и нервы. Основным элементом являетсяпоперечнополосатое веретеновидное мышечное волокно (миосимпласт) (Рис. 36).
При изучении мышц, прежде всего, следует обращать внимание на форму, размеры, расположение мышцы и на ее части. Чем длиннее мышца, тем больший размах движений она может обеспечить. Следует помнить, что активная фаза - сокращение мышцы - сопровождается ее укорочением, т.е. она будет сближать те точки на костях, к которым прикрепляется. Поэтому для суждения о функции каждой конкретной мышцы необходимо знать как точкуее начала (origo), так и точку прикрепления (insertio), имеющие вид сухожилия (tendo). Началом мышцы принято условно считать точку, расположенную ближе к срединной оси тела (проксимально), а точкой прикрепления - более удаленную (дистально).
Начальная часть, особенно длинных мышц, называется головкой (caput), средняя - телом, или брюшком (venter), конечная - хвостом (cauda).
Сухожилия мышц построены из плотной оформленной волокнистой соединительной ткани, волокна которой могут увеличиваться до 4-6% от своей длины. Сухожилие значительно тоньше мышцы, однако способно выдерживать большую нагрузку при растяжении: пяточное (ахиллово) сухожилие выдерживает нагрузку до 500 кг, а сухожилие четырехглавой мышцы бедра - около 600 кг. Необходимо обращать внимание на то, с какой стороны мышца прилежит к суставу, на каком расстоянии от сустава прикрепляется к другой кости. В теле человека отдельные мышцы объединяются в группы соответственно их функции (сгибатели, разгибатели, сжиматели и т.д.). При изучении этого раздела следует также обратить внимание на особенности строения мышц отдельных областей. Например, мимические мышцы имеют точку начала на костях, а точку прикрепления - в коже лица. Мышцы нижних конечностей, как правило, сильные (статические), рассчитанные на длительную тяжелую работу, а мышцы верхних конечностей ловкие (динамические), лучше приспособлены к быстрым точным движениям.
С мускулатурой структурно и функционально связаны различные анатомические образования, способствующие мышечным сокращениям и облегчающие их (синовиальные влагалища, слизистые сумки, блоки, сесамовидные кости). Так, в наиболее, подвижных местах конечностей - в области кисти и стопы формируются влагалища сухожилий мышц (vaginae tendinum), определяющие скольжение сухожилий в строго определенных направлениях. По своему строению это фиброзные и костно-фиброзные каналы, или влагалища, внутри которых залегают синовиальные влагалища (vaginae synoviales tendinum). Образующие их наружный (lam. parietalis) и внутренний (lam. visceralis) листки гладкие и смазаны синовией, что способствует скольжению и свободному движению сухожилий.
В местах, где движение мышц или сухожилий достигает значительной степени, располагаются синовиальные сумки (bursae synoviales), представляющие собой щелевидные полости, заполненные жидкостью, которая способствует уменьшению трения. Сумки, залегающие под сухожилиями мышц, называются подсухожильными синовиальными сумками (bursae synoviales subtendineae), а между кожей и выступающей костью - подкожными синовиальными сумками (bursae synoviales subcutaneae). Сумки, расположенные вблизи суставов, часто сообщаются с их полостью.
Блок (trochlea) представляет собой покрытую хрящом выемку на кости там, где через нее перекидывается сухожилие мышцы. Последнее обычно меняет здесь направление, но благодаря блоку не смещается в стороны.
Такую же функцию выполняют и сесамовидные кости (ossa sesamoidea), которые располагаются в толще сухожилий мышц, обеспечивающих движение в некоторых блоковидных суставах (надколенник, косточки у оснований первых фаланг).
К каждой мышце подходит один или несколько кровеносных сосудов и нервов, обеспечивающих жизнедеятельность (трофику) и работоспособность составляющих ее элементов.
Мышечные волокна имеют характерный полосатый вид. Независимо от формы и размера, все мышцы состоят из отдельных волокон, диаметр которых в больших мышцах конечностей взрослого человека составляет приблизительно 50 мкм; У маленького ребенка он значительно меньше. Диаметр волокон может увеличиться почти вдвое вследствие тренировок силовой направленности и занятий физическим трудом.
МЫШЕЧНАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Соединительная ткань мышцы, являясь ее мягким скелетом (прежде всего в ее брюшке), играет такую же важную роль в функционировании мышцы как органа, как и ее мышечные волокна. Соединительная ткань в брюшке мышцы имеет несколько анатомических частей, связанных с особенностями (размерами и направлениями) составляющих ее коллагеновых волокон.
Соединительная ткань мышцы организована на трех уровнях. Любая соединительная ткань в организме человека, состоит из волокон, заключенных в бесформенное основное вещество. Большинство волокон представляют собой коллаген; по меньшей мере пять из них (I-V) являются иммунологически определенными, остальные волокна представляют собой эластин. Соединительная ткань мышцы имеет три анатомические части - эпимизий, перимизий, эндомизий (Рис. 37; Borg и Caufield, 1980; Rowe, 1981).
Рис. 37 Соединительные ткани мышцы
Эпимизий представляет собой особенно плотную оболочку, которая покрывает всю поверхность брюшка мышцы и отделяет ее от других мышц. Он содержит тесно переплетенные пучки коллагеновых волокон диаметром 600-1800 нм, прикрепленные к перимизию.
Перимизий, также достаточно плотный и относительно толстый, разделяет мышечные волокна на пучки и обеспечивает пути прохождения крупных кровеносных сосудов и нервов через брюшко мышцы. Одни большие коллагеновые пучки расположены вдоль внешних мышечных волокон, другие окружают их. Внизу располагается более тонкая сеть коллагеновых фибрилл: идущих во всех направлениях; некоторые из них соединены с эндомизием. В межперимизиальных пространствах могут находиться артериолы и венулы, нередко с внутримышечными нервными ответвлениями. Поперечный разрез пучка показывает, что мышечные волокна имеют скорее многоугольную (полигональную), а не круговую конфигурацию, которая позволяет разместиться в пучке большему числу волокон. Ширина интерстициального пространства между волокнами не превышает 1 мкм.
Эндомизий покрывает каждое мышечное волокно и состоит из плотной сети коллагеновых фибрилл диаметром от 60 до 120 нм, некоторые из них переходят в тонкую фибриллярную сетчатую структуру перимизия. Можно также предположить, что эндомизий имеет соединения с базальной мембраной - гликопротеиновым слоем, покрывающим внешнюю мембрану мышечного волокна.
Соединительная ткань мышцы обладает множеством функций.
1. В процессе развития соединительная ткань выполняет функцию каркаса (мягкого скелета мышцы), на котором фиксируются мышечные волокна. После завершения развития мышц соединительная ткань продолжает удерживать их вместе и во многом определяет структуру мышечного брюшка. 2. Свободная соединительная ткань перимизия определяет строение саркоплазматического ретикулума - сложной системы трубочек и цистерн, имеющих собственную мембранную стенку, и обеспечивает каналы для кровеносных сосудов и нервов, обслуживающих мышечные волокна (см. рис. 38). 3. Соединительная ткань имеет тенденцию противостоять пассивному растяжению мышцы и. обеспечивает такое распределение сил, при котором вероятность повреждения мышечных волокон сводится к минимуму. Кроме того, свойство эластичности, обусловленное эластиновыми фибриллами и коллагеновыми пучками, позволяет брюшку восстановить свою форму после устранения действия пассивных сил. 4. Эндомизий через боковые соединения с мышечным волокном сообщает некоторую часть сократительной силы сухожилию Окончания мышечных волокон "специализируются" в передаче силы сухожилию. Мышечные волокна по мере приближения к сухожилию значительно сужаются, и диаметр их уменьшается почти на 90 % (Loeb, Pratt, Chanaud и Richmond, 1987). Сужение волокон придает брюшку мышцы его типичную веретенообразную форму. На конце каждого волокна имеются значительные складки плазмолеммы. Они обеспечивают распределение сократительной силы на большей площади, тем самым, снижая нагрузку на поверхность волокна. Кроме того, передача силы под углом обусловливает сдвигающую нагрузку на соседние структуры (Tidball, 1983). Под электронным микроскопом можно увидеть, что миофибриллы не простираются на всем пути к плазмолемме; наоборот, тонкие филаменты (актин) непосредственно под плазмолеммой входят в плотный слой вещества, содержащего «прикрепляющие» белки: винкулин, талин, паксиллин и тензин.
На внешней стороне поверхностной мембраны находится хорошо развитая базальная мембрана, состоящая из нескольких слоев. Здесь также есть соединительные белки. Это фибронектин и тенасцин, присоединяющие молекулы интегрина в плазмолемме к коллагеновым волокнам типа I в сухожилии.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА
Скелетные мышцы человека содержат около 300 млн. мышечных волокон и имеют площадь порядка 3 м2. Целая мышца представляет собой отдельный орган, а мышечное волокно – клетку. Мышечное волокнопредставляет собой вытянутую клетку (ее диаметр около 10-100 мкм, а длина 10-12 см). В состав волокна входят его оболочка - сарколемма, жидкое содержимое - саркопазма, ядро, энергетические центры - митохондрии, белковые депо рибосомы, сократительные элементы - мuофuбрuллы, а также замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са2+, - саркоплазматический ретикулум. Поверхностная мембрана клетки через равные промежутки образует поперечные трубочки, входящие внутрь мышечного волокна, по которым внутрь клетки проникает потенциал действия при ее возбуждении (Рис. 38).
Миофибриллы – это тонкие волокна (диаметр их 1-2 мкм; длина 2-2,5 мкм), содержащие 2 основных вида сократительных белков (протофибрилл): тонкие нити актина и вдвое более толстые нити миозuна. Они расположены таким образом, что вокруг миозиновых нитей находится 6 актиновых нитей, в вокруг каждой актиновой - 3 миозиновых.
Миофибриллы разделены Z-мембранами на отдельные участки - саркомерыZ-мембранам по бокам саркомера. Если посмотреть на волокна под микроскопом, то можно увидеть, что каждое волокно состоит из серии светлых и темных полосок. Светлая полоска - это I-диск, а темная, имеющая более высокий рефрактивный индекс, - А-диск. В середине I-диска проходит плотная линия. Это Z-линия, или Z-диск. Центральная часть А-диска, которая в расслабленном волокне имеет более светлую окраску (Н-зона), "рассекается" другой, более темной структурой, - М-участком (рис. 39). Участок волокна между двумя последовательными Z-линиями называется саркомером. Его длина в расслабленном волокне составляет около 2,2 мкм. , в средней части которых расположены преимущественно миозиновые нити, а актиновые нити прикреплены к
В волокне диаметром 50 мкм содержится до 8000 миофибрилл. Равномерное распределение актиновых и миозиновых филаментов вдоль миофибриллы придает ей и мышечному волокну характерный полосатый вид. Два вида филаментов поддерживают друг друга при помощи поперечных мостиков, кроме того, их поддерживают другие структуры, которые соединяются с соединительной тканью эндомизия и мышечным сухожилием.
Фuламенты актина и миозина соответствуют светлым и темным полосам. Темные А-диски соответствуют присутствию миозиновых (толстых) филаментов, тогда как светлые I - диски содержат актиновые (тонкие) филаменты (Рис. 39).
Рис. 39 Строение саркомера
В находящемся в покое мышечном волокне актиновые филаменты в определенной степени перекрывают миозиновые филаменты; участок, где этого перекрытия нет, представляет собой бледную Н-зону в центральной части А-диска. В середине Н-зоны находится темный участок - М-участок, образованный волокнистыми структурами, которые поперечно соединяют филаменты миозина. Имеется и другое расположение тончайших филаментов, состоящих из титина, стабилизирующего филаменты миозина в продольной оси. Во время сокращения филаменты миозина контактируют с филаментами актина при помощи молекулярных поперечных мостиков и стараются сдвинуть противоположные филаменты актина друг к другу, тем самым снижая ширину Н-зоны и I-дисков. Филаменты актина.содержат, кроме актина, два регуляторных белка - тропонин и тропомиозин, а также "укрепляющий" белок небулин.
Фuламенты миозина удерживаются вместе в М-зоне. Уже в первых исследованиях, в которых использовали оптический микроскоп, в центре А-диска наблюдали темную линию. Применение электронного микроскопа позволило увидеть, что в действительности имеется несколько М-линий, поэтому более уместно говорить об М-зоне (участке), чем об М-линии. Позднее в М-зоне выявили тонкие филаменты (М-филаменты) диаметром около 5 нм, расположенные параллельно филаментам миозина и связанные как с последними, так и друг с другом. Именно поперечные соединения между двумя видами филаментов обусловливают наличие множества М-линий в каждом А-диске. В результате исследований в М-зоне, кроме миозина, удалось выявить еще два белка. Один из них М-белок, или миомезин, его молекулярная масса 165 кДа, второй белок – креатинкиназа; очевидно эти молекулы образуют стоики, удерживающие филаменты миозина в решетке (Рис. 40).
Титин помогает стабилизировать филаменты миозина. Если для растворения филаментов актина в мышечном волокне взять такой растворитель, как гелсолин, то мы увидим, что толстые филаменты миозина соединяются с Z-дисками тончайшими полосками. Эти полоски, диаметр которых приблизительно 5 нм, а длина 1 мкм, состоят из исключительно большого белка титина (молекулярная масса 3000 кДа); очевидно, 6 или, возможно, 12 молекул титина образуют каждую полоску (Trinick, 1991). Часть титиновой полоски перекрывает миозиновый филамент и прочно к нему прикрепляется, остальная часть простирается через I-диск и, очевидно, обладает эластичными свойствами. Исходя из структуры и положения титиновых полосок, можно предположить, что каждая их пара выполняет функцию продольного стабилизатора миозинового филамента, удерживая его в центре саркомера во время сокращения и расслабления. Вполне вероятно, что титиновые полоски в значительной степени обусловливают эластичность мышечного волокна при его растяжении, либо пассивном, либо активном. На рис. 41 показаны также полоски небулина, поддерживающие актиновые филаменты.
Филаменты актина привязаны у Z-линиu. Филаментам актина, подобно филаментам миозина, необходима позиционная поддержка, которая частично обеспечивается поперечными мостиками, а частично вследствие их вхождения в Z-линию в центре каждого I-диска. Z-линия состоит главным образом из белков а-актинина, десмина, виментина и синемина. а-Актинин - белковый димер с молекулярной массой 180 кДа, как полагают, прикрепляет концы филаментов актина на одной стороне Z-линии к таким же концам на другой стороне. Еще одной особенностью Z-линии является то, что точка, в которой филамент актина прикрепляется на одной стороне, находится между двумя точками соединения на противоположной стороне линии.
Десмин, виментин и синемин, а также остальные существенные белковые компоненты Z-линии образуют промежуточные филаменты, окутывающие линию и соединяющие вместе соседние линии по продольным и поперечным осям (рис. 42). Именно эти промежуточные филаменты благодаря поперечным соединениям от одной Z-линии к другой обеспечивают "регистрацию" всех миофибрилл в отдельном мышечном волокне. Z-линии при помощи промежуточных филаментов соединяются также с цитоскелетом под плазмолеммой и непосредственно с плазмолеммой. Благодаря соединительным белкам плазмолеммы Z-диски соединены с базальной мембраной и эндомизием. Каждое поперечное расположение промежуточных филаментов ограничивает саркомер.
ТРУБЧАТЫЕ СИСТЕМЫ
В начале прошлого столетия Вератти (1902) обнаружил тончайшую взаимопереплетенную сеть в мышечном волокне. Лишь много лет спустя, при помощи электронного микроскопа удалось установить, что эта структура имеет трубчатую систему, состоящую из двух частей (Franzini-Armstrong и Porter, 1964).
Саркоплазматuческuй ретикулум (СР) окружает мuофuбрuллы. Система каналов проходит по длинной оси мышечного волокна и окружает отдельные миофибриллы. Благодаря ответвлениям продольные каналы вокруг миофибрилл соединяются друг с другом, а также с каналами, окружающими другие миофибриллы. При сокращении мышечного волокна продольные каналы становятся шире и короче. Функция СР заключается в выделении Са2+ в цитозол вокруг миофибрилл, где он сочетается с тропонином-С, обеспечивая сокращение. Когда Са2+ направляется обратно в СР, сокращение прекращается. для осуществления функции "активации" волокна СР имеет мембрану, содержащую как каналы выделения Са2+ (рианодиновые рецепторы), так и насосыСа2+ АТФ.
Поперечная трубчатая система соединяет поверхность волокна свнутренней частью. Поперечная трубчатая система (Т - система) расположена перпендикулярно длинной оси мышечного волокна в виде каналов, окружающих миофибриллы через равные промежутки. В каждом саркомере мышечных волокон млекопитающих, в том числе человека, есть две зоны поперечных трубочек; они находятся на стыке А- и I-дисков. В волокнах сердечной мышцы, а также в волокнах мышц лягушки каждый саркомер имеет только одну Т-систему, которая находится в Z-линии.
Поскольку Т-трубочки окружают миофибриллы, то они "прерывают" продольные каналы СР. В этих точках СР расширяется, образуя цистерны (или латеральные мешочки); соседние мешочки соединяются друг с другом. Каждая относительно узкая Т-трубочка окружена цистерной; три элемента, окружающие миофибриллу, называют триадой. По данным Пичи (1965), приблизительно 80 % поперечной тубулярной системы в мышечном волокне лягушки окружено СР. На поверхности мышечного волокна Т-трубочки образуют небольшие отверстия в месте соединения А- и I-диска, и их мембраны соединяются с плазмолеммой. Исследования показывают, что Т - трубочки содержат внеклеточную жидкость. Главная функция трубочек - проведение возбуждающих импульсов с поверхности мышечного волокна в его центральную зону, что ведет к стимуляции выделения Са2+ из СР.
ЯДРА И МИТОХОНДРИИ
Мышечное волокно содержит множество ядер (миоядра), разбросанных вдоль внутренней поверхности плазмолеммы; каждое ядро привязано к ней двумя мембранами. Задача ядер - подготовить и направить в цитоплазму "инструкции", касающиеся белкового синтеза. Эти "инструкции" поступают из генов, содержащихся в хромосомах.
Помимо ядер, миофиламентов и трубчатых систем, другими важными структурами цитоплазмы являются митохондрии. В процессе эволюции эти органеллы специализировались в образовании АТФ - основного энергообразующего вещества живых организмов. Большинство клеточных ферментов содержится в митохондриях, остальные - в цитозоле. Кроме того, цитоплазма содержит два химических соединения, которые митохондрии могут использовать для производства АТФ. Это гранулы гликогена и липидные капельки.
При помощи оптического микроскопа невозможно отличить миоядра от ядер клеток-сателлитов. Последние составляют менее 1 % ядер мышечного волокна у взрослых людей. Их можно дифференцировать только при помощи электронного микроскопа, который позволяет также обнаружить наличие мембран-близнецов, отделяющих цитоплазму клетки-сателлита от цитоплазмы мышечного волокна. Клетки-сателлиты играют особенно важную роль в регенерации мышцы после болезни или травмы.
Митохондрии обеспечивают мышечное волокно АТФ. Митохондрии (саркосомы) являются яйцевидными структурами с максимальным диаметром 1-2 мкм. Каждая митохондрия имеет двойную мембрану. Внутренняя мембрана периодически собирается в складки, образуя гребешки; эти складки выходят в центральный компартмент митохондриона. Мaтpикс между гребешками является домом ферментных систем, необходимых для осуществления цикла трикарбоновой кислоты (цикл Кребса), при котором расщепляется пируват на двуокись углерода и воду. Большая часть энергии, выделяемой в результаге последовательных реакций, захватывается цепочкой цитохрома, обеспечивающей образование АТФ. Сейчас известно, что цепочка цитохрома находится на внутренней мембране гребешков вместе с АТФ-образующим ферментом (АТФ-синтетаза). Мaтpикс содержит также несколько копий ДНК самого митохондриона. Внешняя мембрана гребешка содержит ферменты, необходимые для липидного синтеза. Митохондрии находятся, в частности, между миофибриллами в участке Z-линии.
Гликогеновые гранулы, иногда достигающие в диаметре 25-40 нм, разбросаны в цитозоле и являются главным источником энергии для мышечного волокна. Липидные капельки нередко можно обнаружить вблизи митохондрий, они содержат жирные кислоты или триглицериды и являются важными дополнительными источниками энергии.
МЫШЕЧНЫЕ РЕЦЕПТОРЫ
Мышцы содержат морфологически определенные рецепторы, а также свободные нервные окончания.Мышечные рецепторы принято разделять на четыре вида:
Мышечные и нервно-сухожильные веретена играют наиболее важную роль в сообщении информации о механических явлениях. Мышечные веретена созданы исключительно для восприятия длины мышечных волокон во время сокращений, тогда как нервно-сухожильные веретена в большей степени воспринимают силу, развиваемую волокном. Тельце формы Пачини имеет нервное окончание, покрытое соединительной тканью. Этот рецептор весьма чувствителен к быстрым и небольшим деформациям и широко распространен в таких тканях, как кожа, надкостница и внутренние органы живота; в мышце этот рецептор, по-видимому, играет не самую важную роль при передаче информации о сокращении и расслаблении. Одна из важнейших функций свободных нервных окончаний - обеспечение информаций о метаболическом статусе интерстициальной жидкости и, следовательно, мышечных волокнах; эти окончания реагируют на изменения, в напряжении кислорода, [Н+], [К+] и, возможно, другие сигналы.
Рассмотрим подробнее мышечное веретено. Его характерными особенностями являются, во-первых, сложность структуры, а во-вторых, способ регулирования его реактивности со стороны ЦНС.
Мышечные веретена - длинные тонкие структуры, окруженные экстрафузальными волокнами. Длина мышечного волокна намного больше, чем его ширина. Диаметр веретена в его наиболее широкой части - капсуле составляет всего 80-250 мкм, тогда как длина может достигать 10 мм. Мышечные веретена окружены экстрафузальными мышечными волокнами, составляющими большую часть брюшка мышцы. Количество мышечных веретен в брюшке мышцы значительно колеблется. Наибольшее их количество отмечается в мышцах тыльной части шеи и в небольших мышцах кисти, наименьшее - в больших мышцах рук и ног. Это различие, очевидно, связано со способностью головы и пальцев быстро и точно выполнять короткие движения.
Мышечное веретено содержит два вида мышечных волокон. Характерная веретенообразная форма обусловлена наличием в средней части веретена капсулы соединительной ткани, содержащей жидкость. Состав и функция последней до сих пор не определены, хотя довольно давно предполагают, что это лимфа. В продольной оси веретена центральный и внешний участок нередко называют экваториальным и полярным соответственно.
Удивительной особенностью мышечного веретена является то, что его чувствительность как механорецептора можно регулировать; это оказывается возможным благодаря прикреплению чувствительных нервных окончаний к интрафузальным (внутри веретена) мышечным волокнам, которые могут сокращаться и расслабляться. Два вида интрафузальных волокон различают на основании количества и распределения в них ядер (рис. 43). Волокна ядерного мешочка имеют экстраординарное количество ядер в своей средней части капсуле; в этом участке мышечное волокно состоит практически из одних ядер. Волокна ядерной цепочки, в свою очередь, характеризуются более равномерным распределением ядер вдоль ее длины; ядра находятся в центре волокон. Первые волокна более толстые и длинные, к тому же они малочисленны. В каждом веретене обычно содержится только 2 (от 1 до 4) волокна, тогда как волокон ядерной цепочки может быть 12. Два вида волокон ядерного мешочка называют мешочек 1 и мешочек 2.
Рис. 43 Интрафузальные мышечные волокна
Интрафузальные мышечные волокна имеют собственную иннервацию.
Первичные и вторичные чувствительные окончания находятся на интрафузальных мышечных волокнах и вокруг них. Каждое веретено имеет одно большое центростремительное нервное волокно - аксон группы Ia, который заканчивается спиралями, окружающими каждое интрафузальное волокно в экваториальном участке; этот тип окончаний называется первичным окончанием (см. рис. 43). В веретенах человека первичные окончания не имеют формы спиралей, а образуют многочисленные пучки, напоминающие букву С, которые частично окружают интрафузальные волокна (Kennedy, 1970).
Кроме того, каждое веретено имеет более тонкое чувствительное нервное волокно - аксон группы II, заканчивающийся на волокнах ядерной цепочки и мешочка2 окончаниями в виде цветка; эта вторичные окончания.
Нервно-сухожильные веретена открыл Камилло Гольджи (1903). Они не столь многочисленны, как мышечные веретена; другое их отличие от мышечных - содержание в мышечно-сухожильном соединении, а не в брюшке мышцы. Кроме того, они характеризуются более простой структурой.
Нервно-сухожильные веретена прикрепляются к экстрафузальным мышечным волокнам. Длина нервно-сухожильных веретен обычно составляет 0,5-1,0 мм, а их диаметр - 0,1-0,2 мм. Отдельный нервный аксон, имеющий относительно большой диаметр, проводит импульсы в спинной мозг и называется аксоном Ib; он выходит из немиелинированных веточек, соприкасающихся с сухожильными пучками (рис. 44, а). Под очень сильным увеличением можно увидеть, что тончайшие нервные веточки прокладывают свой путь между пучками коллагеновых волокон (рис. 44, б). Когда мышечные волокна сокращаются, коллагеновые пучки распрямляются и сжимают нервные веточки, посылая в них импульсы (Swett и Schoиltz, 1975). Через коллагеновые пучки нервно-сухожильное веретено соединяется с 4-25 мышечными волокнами.
Нервно-сухожильные веретена возбуждаются сокращениями мышцы. В отличие от мышечных веретен, нервно-сухожильные веретена не являются столь чувствительными к пассивному растяжению мышцы и ее сухожилия. При сокращении мышцы, однако, происходит разрядка нервно-сухожильных веретен, и их чувствительность проявляется в том, что они могут реагировать на отдельную двигательную единицу через 1-2 мышечных волокна, прикрепляющих ее к нервно-сухожильному веретену. На каждое нервно-сухожильное веретено оказывают влияние от 4 до 15 двигательных единиц (Hoиk и Henneman, 1967). Аксоны Ib, которые выходят из нервно-сухожильных веретен, отличаются от аксонов мышечных веретен тем, что они оказывают ингибиторное воздействие на мотонейроны одной и той же мышцы и возбуждающее - на мотонейроны мышц-антагонистов. Через рефлекторные соединения они обеспечивают постоянную величину силы, произведенной мышцей, если это необходимо.
Рис. 44 Нервно-сухожильные веретена
Итак, нам осталось рассмотреть последний тип мышечных рецепторов, наиболее простой по своей структуре и наиболее многочисленный; это свободное нервное окончание.
Свободные нервные окончания могут реагировать на определенные стимулы или повреждение ткани. Свободные нервные окончания передают информацию в спинной мозг через мельчайшие миелинированные нервные волокна (афференты группы III) и немиелинированные (афференты группы IV). Свободные нервные окончания иннервируют практически все структуры в брюшке мышцы: экстрафузальные волокна, различные виды соединительной ткани, включая эпимизий, крупные кровеносные сосуды и даже мышечные и нервно-сухожильные веретена (Stacey, 1969). Электрофизиологическая регистрация афферентов группы III и IV показывает, что многие свободные нервные окончания чувствительны к механическим стимулам, таким, как те, что связаны с мышечным сокращением, или давлением, или растяжением (Mense иMeyer, 1985). Другие свободные нервные окончания реагируют главным образом на изменения температуры или химической среды; например, увеличение концентрации К+ или молочной кислоты может спровоцировать разряд (Thimm и Ваиm, 1987). Некоторые свободные нервные окончания имеют высокий порог и посылают импульсы только в ответ на стимулы, способные вызвать повреждение ткани (Hense и Stahnke, 1983). Это ноцирецепторные окончания; вполне вероятно, что они синтезируются в результате выделения нейропептидов (например, брадикинина) или других стимуляторов (например, арахидоновой кислоты) в участке повреждения.
Некоторые немиелинированные окончания становятся все более активными в процессе ишемических сокращений. Эти волокна, а также волокна, обусловливающие увеличение концентрации К+ или молочной кислоты, могут отвечать не только за мышечный дискомфорт во время выполнения физической нагрузки, но и за рефлексы сердечно-сосудистой и дыхательной систем (Coote, Hi1ton, Perez.;Gonzalez, 1971).
Мышцы снабжены многочисленными вспомогательными аппаратами. К ним относятся фасции, фиброзные и синовиальные влагалища сухожилий, синовиальные сумки, блоки мышц. Фасция (от лат. fascia-полоса) - это оболочка мышцы, которая образует ее соединительнотканный чехол.
Фасции сформированы плотной оформленной соединительной тканью. Пучки коллагеновых волокон образуют сеть, в наибольшей степени противостоящую растяжению. Фасции, отграничивающие мышцы друг от друга, выполняют механическую функцию, создавая опору для брюшка при сокращении, ослабляют трение мышц. Мышцы, как правило, соединены с фасциями с помощью рыхлой неоформленной соединительной ткани. Однако некоторые мышцы начинаются от фасции и прочно с ними сращены (на голени; предплечье). В организме человека различают фасции собственные и поверхностные. Каждая мышца или несколько мышц имеют собственную фасцию. Если фасция лежит поверхностно, отделяет мышцы от подкожной клетчатки – это поверхностная фасция, если расположена между глубоко лежащими мышцами - это глубокая фасция. Поверхностная фасция обычно на всем протяжении окутывает все мышцы какой-либо области (например, груди, конечности). Фасции разделяют группы мышц, выполняющих различную функцию. Фасциальные узлы (утолщения фасций) расположены в участках соединения фасций друг с другом. Они укрепляют фасциальные влагалища сосудов и нервов - фасции, фасциальные узлы прочно сращены с костями, дополняют костный скелет, формируя мягкий остов, или мягкий скелет.
Строение фасций зависит от функции мышцы, от напряжения, которое фасция испытывает при сокращении мышцы. Там, где мышцы развиты хорошо, фасции более плотные (например, широкая фасция бедра, фасция голени), и, наоборот, мышцы выполняющие небольшую нагрузку, окружены рыхлой фасцией. В местах, где фасции проходят над сосудисто-нервными пучками, фасции утолщаются, образуя сухожильные дуги. В области голеностопного, лучезапястного суставов утолщенные фасции прикрепляются к костным выступам, образуя удерживатели сухожилий и мышц. В расположенных под ними пространствах в костно-фиброзных влагалищах проходят сухожилия. В ряде случаев фиброзные влагалища содержат несколько сухожилий, в других – каждое сухожилие имеет свое самостоятельное влагалище. Удерживатели предотвращают боковые смещения сухожилий при сокращении мышц.
Синовиальное влагалище отделяет движущееся сухожилие от неподвижных стенок фиброзного влагалища и костно-фиброзного канала, устраняет трение их друг о друга. Синовиальное влагалище представляет собой замкнутую щелевидную полость, заполненную небольшим количеством жидкости и отграниченную висцеральным и париетальным листками. Висцеральный (внутренний) листок окружает со всех сторон сухожилие и сращен с ним. Париетальный (наружный) листок выстилает стенки фиброзного влагалища. Оба листка переходят друг в друга на концах влагалища и, вдоль сухожилия с его внутренней, обращенной к кости стороны. Удвоенный листок влагалища, соединяющий внутренний и наружный листки, называется брыжейкой сухожилия или мезотендинием. В нем проходят кровеносные сосуды и нервы, снабжающие сухожилие. При движении сухожилия в фиброзном влагалище вместе с ним движется внутренний листок, который благодаря содержащейся в щелевидной полости синовиального влагалища жидкости свободно скользит вдоль наружного листка. Синовиальное влагалище может окружать одно или несколько сухожилий, если они лежат в одном канале. Соседние влагалища могут сообщаться друг с другом. В зонах, где сухожилие или мышца перекидывается через кость или через соседнюю мышцу, имеются синовиальные сумки, которые, подобно описанным синовиальным влагалищам, устраняют трение. Синовиальная сумка представляет собой плоский двухстенный мешочек (вырост синовиальной оболочки), содержащий небольшое количество синовиальной жидкости. Наружная поверхность одной из стенок сумки, сращена с движущимся органом (мышца, сухожилие), другая стенка сращена с надкостницей или сухожилием другой мышцы. Размеры сумок варьируют от нескольких миллиметров до нескольких сантиметров. Иногда синовиальная сумка сообщается с полостью сустава. Синовиальная сумка может располагаться между сухожилием и блоком мышцы, костным выступом, имеющим желобок, покрытый хрящом. Блок, изменяя направление сухожилия, создает для него опору и одновременно увеличивает угол прикрепления сухожилия к кости, тем самым, увеличивая рычаг приложения силы. Такую же функцию выполняют сесамовидные кости, образующиеся в толще некоторых сухожилий или сращенные с ними, например гороховидная кость, или надколенник. |
|||||
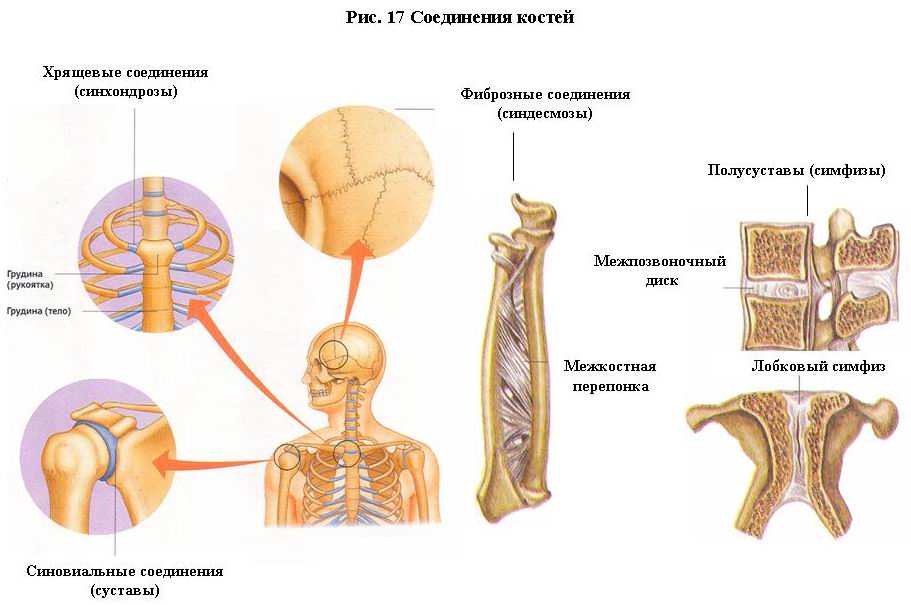

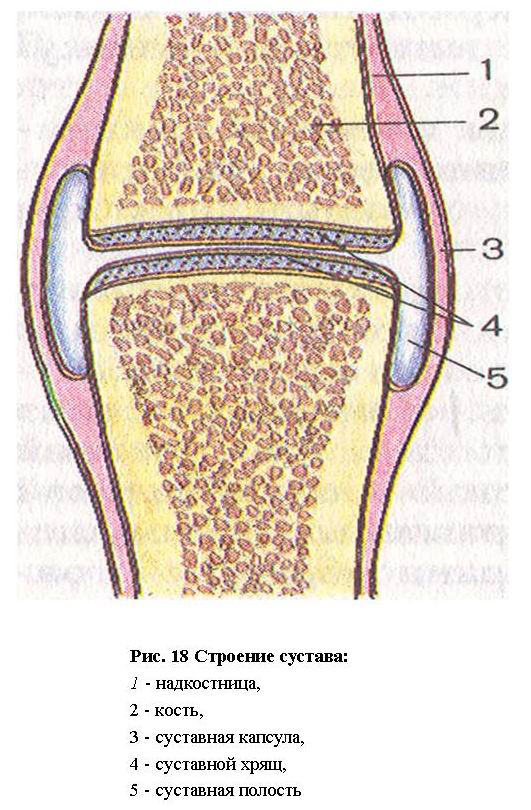

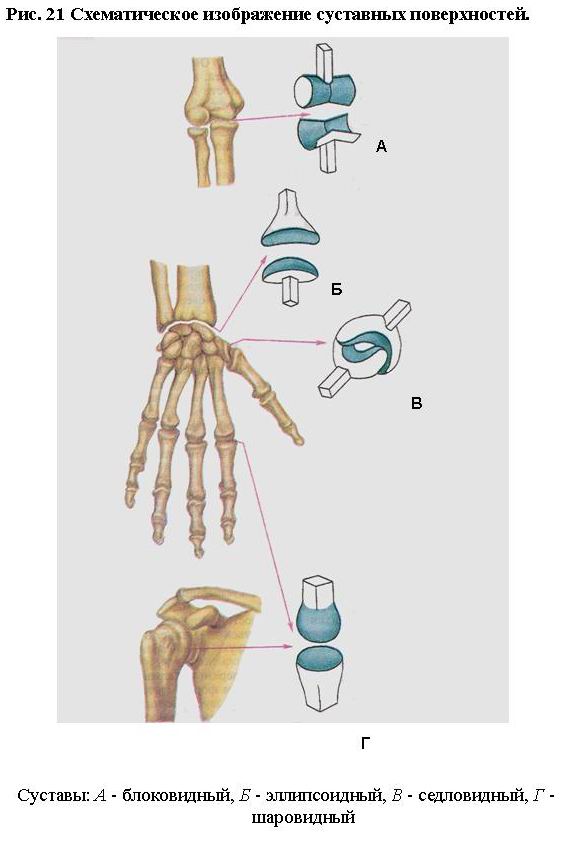
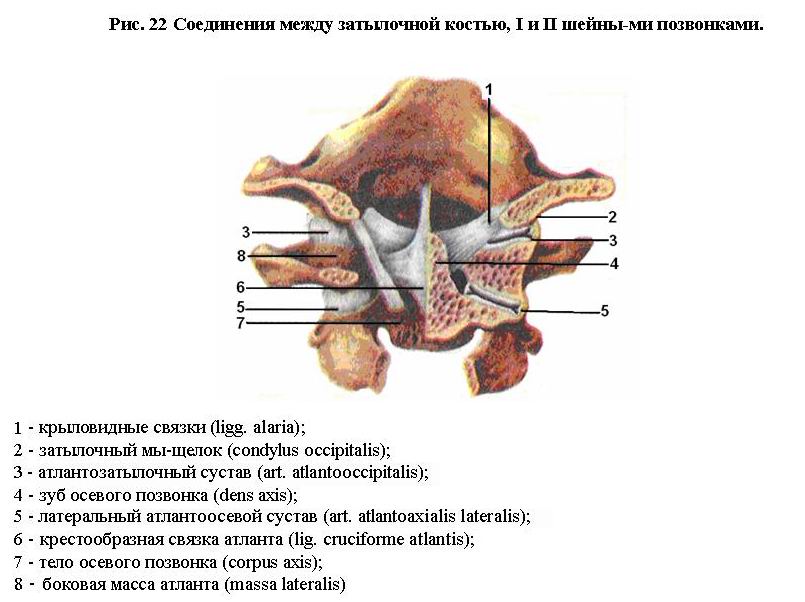
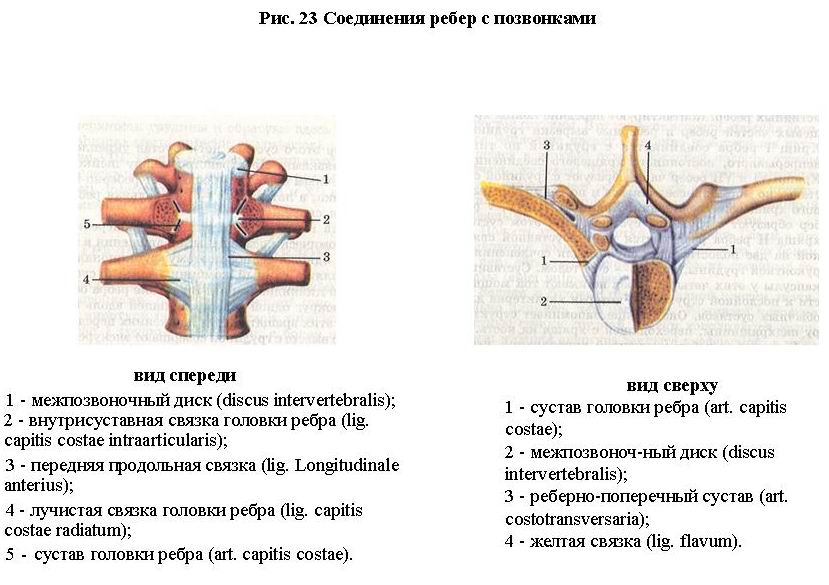
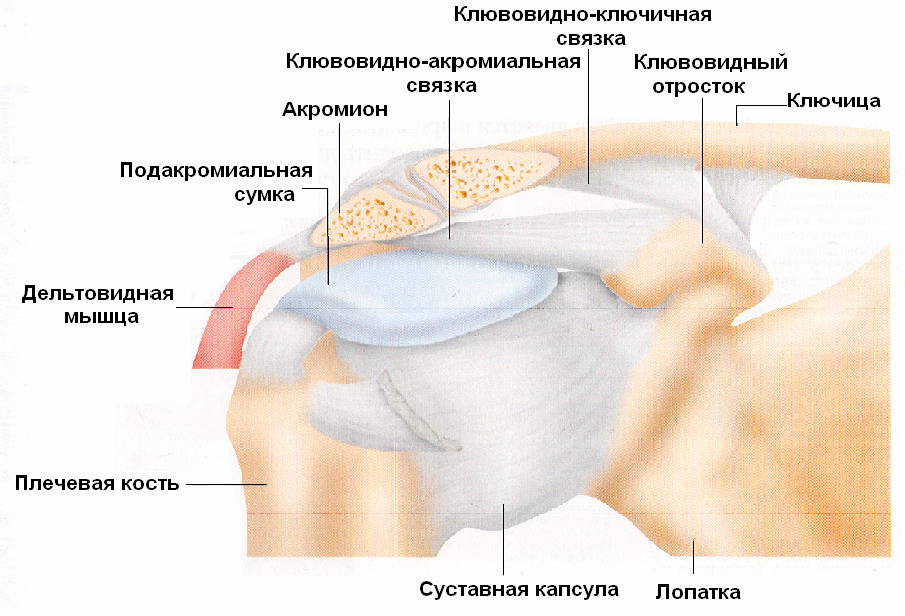
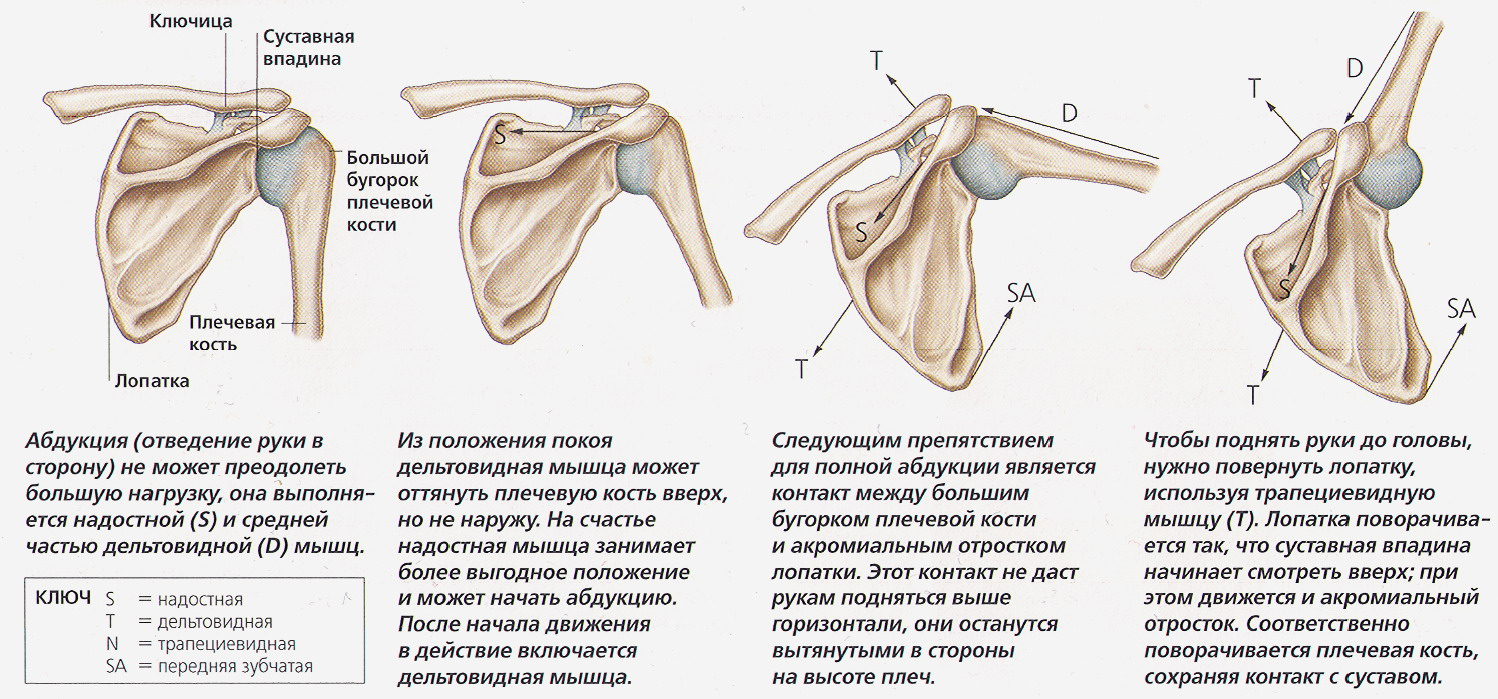
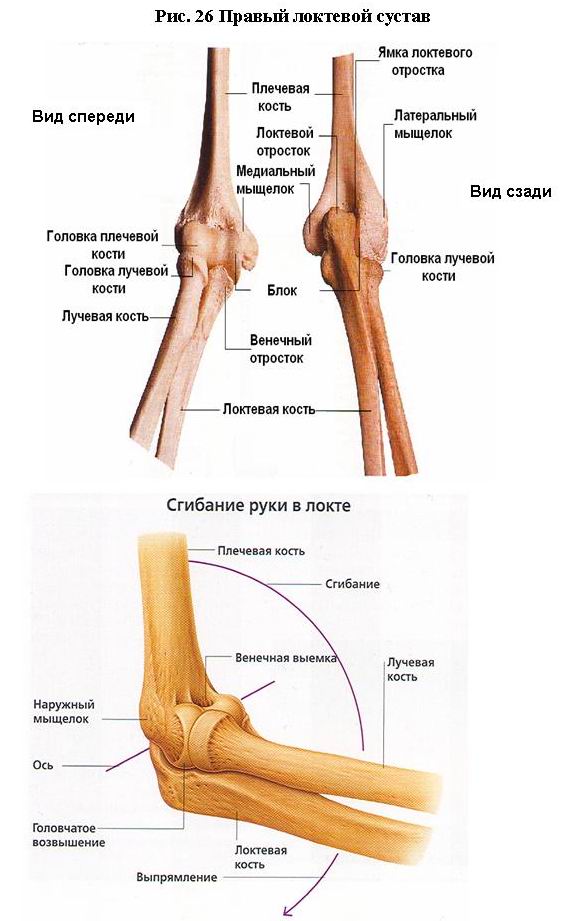
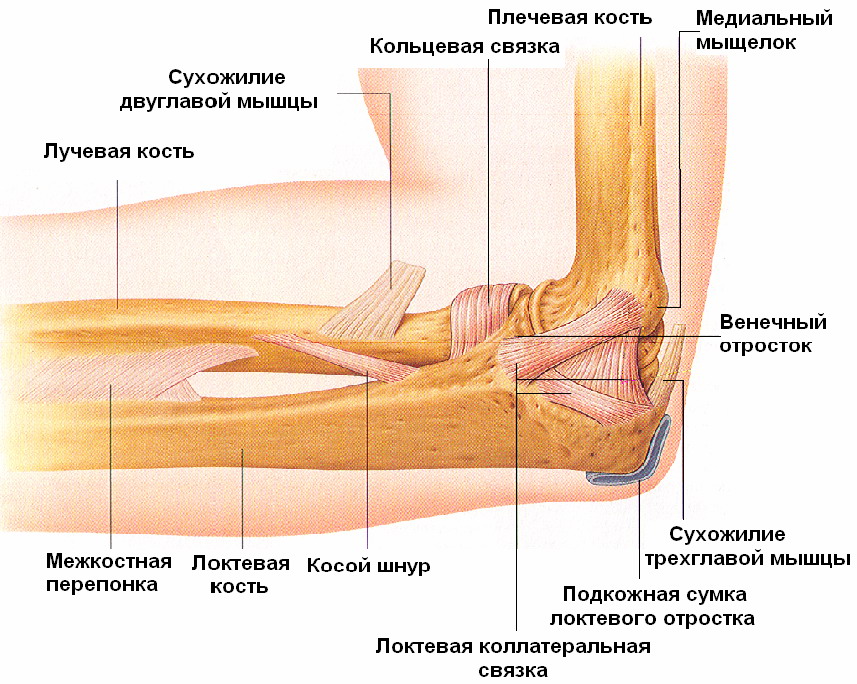
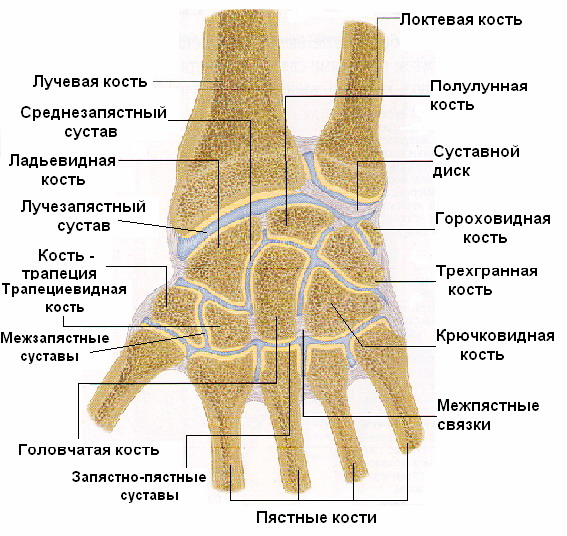
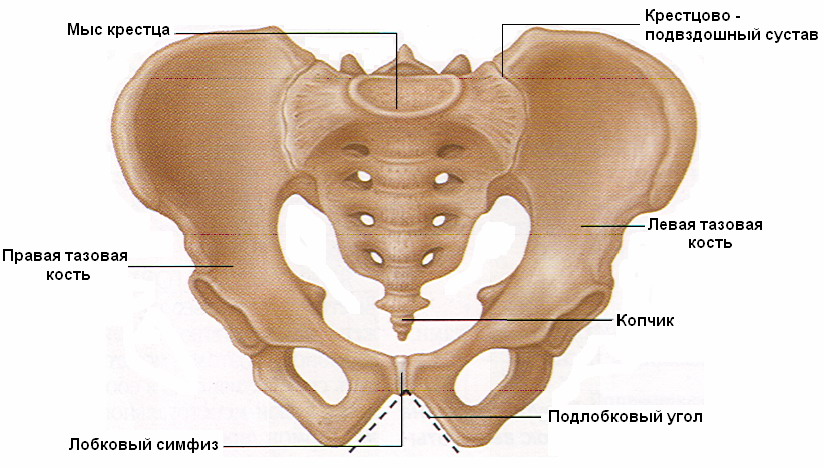
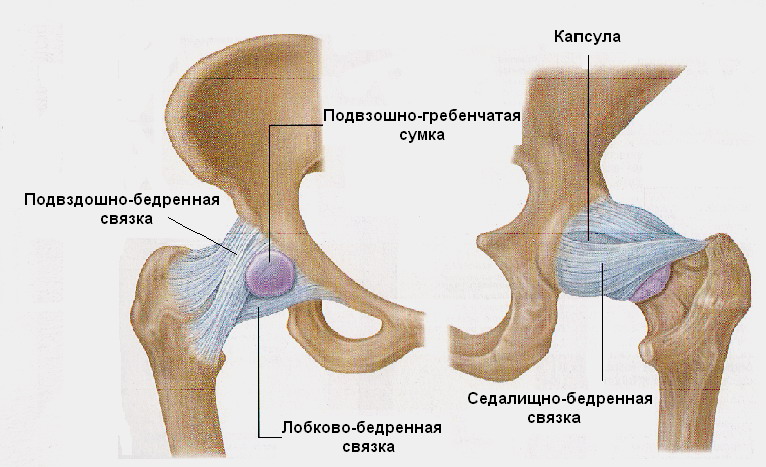
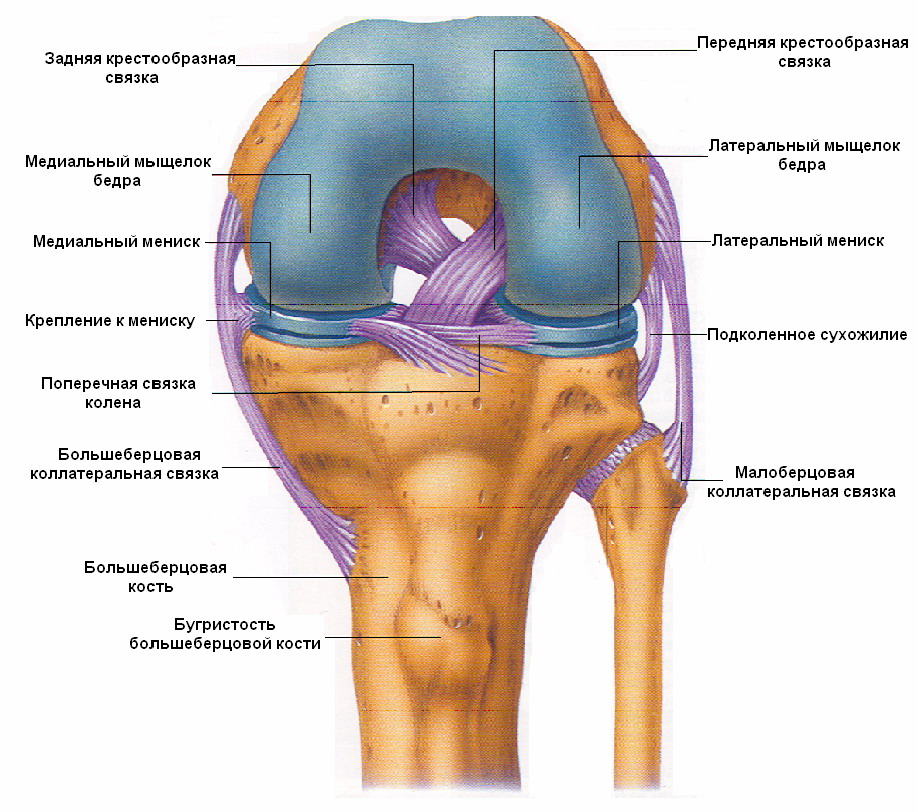
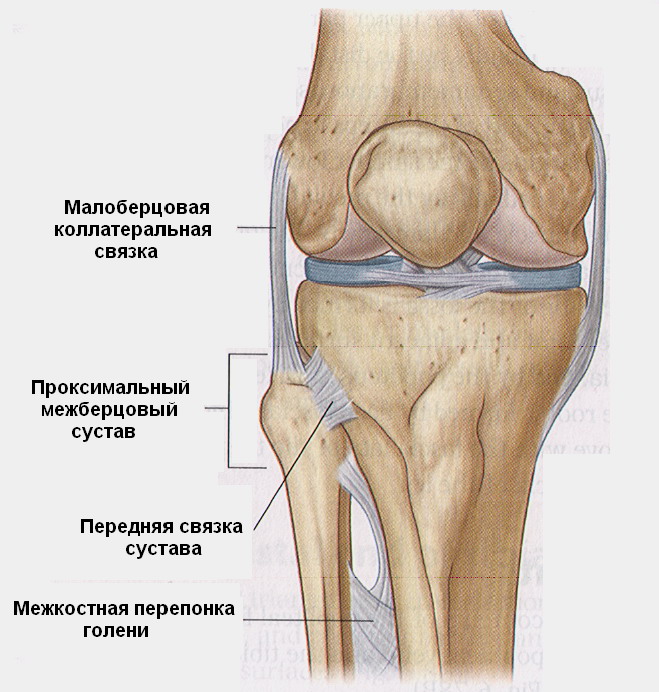
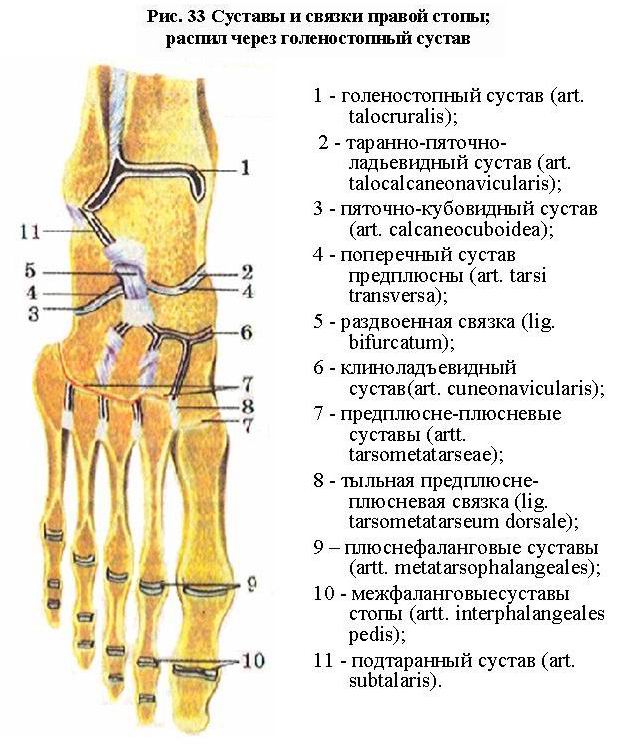

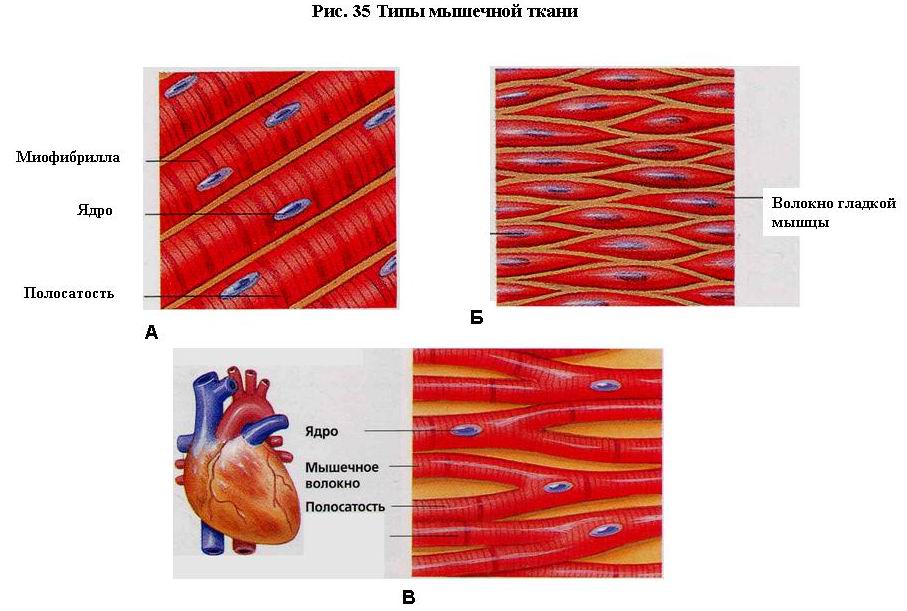







Рис. 37 Соединительные ткани мышцы
Эпимизий представляет собой особенно плотную оболочку, которая покрывает всю поверхность брюшка мышцы и отделяет ее от других мышц. Он содержит тесно переплетенные пучки коллагеновых волокон диаметром 600-1800 нм, прикрепленные к перимизию.
Перимизий, также достаточно плотный и относительно толстый, разделяет мышечные волокна на пучки и обеспечивает пути прохождения крупных кровеносных сосудов и нервов через брюшко мышцы. Одни большие коллагеновые пучки расположены вдоль внешних мышечных волокон, другие окружают их. Внизу располагается более тонкая сеть коллагеновых фибрилл: идущих во всех направлениях; некоторые из них соединены с эндомизием. В межперимизиальных пространствах могут находиться артериолы и венулы, нередко с внутримышечными нервными ответвлениями. Поперечный разрез пучка показывает, что мышечные волокна имеют скорее многоугольную (полигональную), а не круговую конфигурацию, которая позволяет разместиться в пучке большему числу волокон. Ширина интерстициального пространства между волокнами не превышает 1 мкм.
Эндомизий покрывает каждое мышечное волокно и состоит из плотной сети коллагеновых фибрилл диаметром от 60 до 120 нм, некоторые из них переходят в тонкую фибриллярную сетчатую структуру перимизия. Можно также предположить, что эндомизий имеет соединения с базальной мембраной - гликопротеиновым слоем, покрывающим внешнюю мембрану мышечного волокна.
Соединительная ткань мышцы обладает множеством функций.
1. В процессе развития соединительная ткань выполняет функцию каркаса (мягкого скелета мышцы), на котором фиксируются мышечные волокна. После завершения развития мышц соединительная ткань продолжает удерживать их вместе и во многом определяет структуру мышечного брюшка.
2. Свободная соединительная ткань перимизия определяет строение саркоплазматического ретикулума - сложной системы трубочек и цистерн, имеющих собственную мембранную стенку, и обеспечивает каналы для кровеносных сосудов и нервов, обслуживающих мышечные волокна (см. рис. 38).
3. Соединительная ткань имеет тенденцию противостоять пассивному растяжению мышцы и. обеспечивает такое распределение сил, при котором вероятность повреждения мышечных волокон сводится к минимуму. Кроме того, свойство эластичности, обусловленное эластиновыми фибриллами и коллагеновыми пучками, позволяет брюшку восстановить свою форму после устранения действия пассивных сил.
4. Эндомизий через боковые соединения с мышечным волокном сообщает некоторую часть сократительной силы сухожилию
Окончания мышечных волокон "специализируются" в передаче силы сухожилию. Мышечные волокна по мере приближения к сухожилию значительно сужаются, и диаметр их уменьшается почти на 90 % (Loeb, Pratt, Chanaud и Richmond, 1987). Сужение волокон придает брюшку мышцы его типичную веретенообразную форму. На конце каждого волокна имеются значительные складки плазмолеммы. Они обеспечивают распределение сократительной силы на большей площади, тем самым, снижая нагрузку на поверхность волокна. Кроме того, передача силы под углом обусловливает сдвигающую нагрузку на соседние структуры (Tidball, 1983). Под электронным микроскопом можно увидеть, что миофибриллы не простираются на всем пути к плазмолемме; наоборот, тонкие филаменты (актин) непосредственно под плазмолеммой входят в плотный слой вещества, содержащего «прикрепляющие» белки: винкулин, талин, паксиллин и тензин.
На внешней стороне поверхностной мембраны находится хорошо развитая базальная мембрана, состоящая из нескольких слоев. Здесь также есть соединительные белки. Это фибронектин и тенасцин, присоединяющие молекулы интегрина в плазмолемме к коллагеновым волокнам типа I в сухожилии.