

Открытие кроссинговера. При допущении размещения в одной хромосоме более одного гена встает вопрос о том, могут ли аллели одного гена в гомологичной паре хромосом меняться местами, перемещаясь из одной гомологичной хромосомы в другую. Если бы такой процесс не происходил, то гены комбинировались бы только путем случайного расхождения негомологичных хромосом в мейозе, а гены, находящиеся в одной паре гомологичных хромосом, наследовались бы всегда сцепленно — группой. Исследования Т. Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. Кроссинговер обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, так же как и сцепление, оказалось общим для всех животных, растений и микроорганизмов. Наличие обмена идентичными участками между гомологичными хромосомами обеспечивает рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. Генетический анализ кроссинговера. Какими же генетическими методами можно отличить явление сцепленного наследования от независимого комбинирования генов? О перекресте хромосом можно судить на основе учета частоты возникновения организмов с новым сочетанием признаков. Такие организмы называются рекомбинантами. Явление кроссинговера было открыто на дрозофиле. Рассмотрим один из классических опытов Моргана, позволивший ему доказать, что гены находятся в хромосомах в определенном порядке. Когда гены находятся в разных парах хромосом, то, например, генотип дигетерозиготы записывается так. Если гены находятся в одной паре гомологичных хромосом, формула видоизменяется. При этом аллели одного генa (Аа и ВЬ), находящиеся в гомологичных хромосомах, пишутся строго одна под другой. У дрозофилы рецессивный ген черной окраски тела обозначается, а его доминантная аллель, определяющая дикую серую окраску,— ген рудиментарных крыльев — vg, нормальных vg+. При скрещивании мух, различающихся по двум парам сцепленных признаков,— серых с рудиментарными крыльями и черных с нормальными крыльями гибриды по фенотипу серые с нормальными крыльями. b vgc На рисунке 55 представлены два анализирующих скрещивания: в одном дигетерозиготой является самец, в другом — самка. Если гибридные самцы скрещиваются с самками, гомозиготными по обоим рецессивным генам. то в потомстве получается расщепление в отношении 1 серотелая с рудиментарными крыльями: 1 чернотелая с нормальными крыльями. Такое расщепление показывает, что данная дигетерозигота образует только два сорта гамет b+vg и bvg+ вместо четырех, причем сочетание генов в гаметах самца соответствует тому, которое было у его родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмена участками гомологичных хромосом. В дальнейшем выяснилось, что у самцов дрозофилы действительно как в аутосомах, так и в половых хромосомах в норме не происходит кроссинговер. Поэтому при анализирующем скрещивании в потомстве появляются только две исходные родительские комбинации признаков в равных количествах, что указывает на полное сцепление генов, находящихся в одной паре гомологичных хромосом.
Может
возникнуть предположение, что серая
окраска тела и рудиментарные крылья, а
также черное тело и нормальные крылья
— это пары признаков, наследующихся
вместе вследствие плейотропного действия
одного гена. Однако если взять для
анализа гетерозиготных самок, а не
самцов, то в Fb наблюдается иное расщепление.
Кроме родительских комбинаций признаков
появляются новые типы — мухи с черным
телом и рудиментарными крыльями, а также
мухи с серым телом и нормальными крыльями.
В этом скрещивании сцепление тех
же
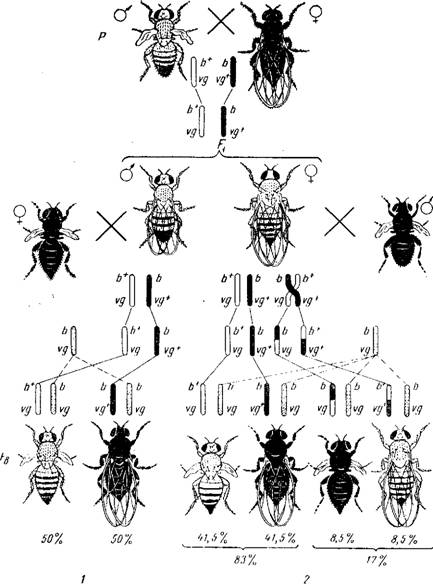 55.
Наследование
сцепленных признаков у дрозофилы: / —
в случае отсутствия кроссинговера
(гетерозиготный самец F,); 2— в случае
наличия кроссинговера (гетерозиготная
самка F,); В Fb изображены только самки,
так как самцы тех же фенотипов; Ь* —
серая; Ь — темная окраска тела; og* —
нормальные; vg —
зачаточные крылья.
генов
нарушается за счет того, что гены в
гомологичных хромосомах поменялись
местами благодаря кроссинговеру.
Гаметы
с хромосомами, претерпевшими кроссинговер,
называют кроссоверными, а не претерпевшие
— некроссоверными. Соответственно
организмы, возникшие от сочетания
кроссоверных гамет гибрида с гаметами
анализатора, называют кроссоверами или
рекомбинантами, а возникшие от сочетания
некроссоверных гамет с гаметами
анализатора — некроссоверными или
нерекомбинантными.
Закон сцепления
Моргана. При анализе расщепления в
случае кроссинговера обращает на себя
внимание определенное количественное
отношение кроссоверных и некроссоверных
классов. Обе исходные родительские
комбинации признаков, образовавшиеся
из некроссоверных гамет, оказываются
в потомстве анализирующего скрещивания
в равном количественном отношении. В
указанном опыте с дрозофилой тех и
других особей было примерно по 41,5%- В
сумме некроссоверные мухи составили
83% от общего числа потомков. Два
кроссоверных класса по числу особей
также одинаковы, и сумма их равна
17%.
Частота кроссинговера не зависит
от аллельного состояния генов, участвующих
в скрещивании. Если в качестве родителей
Ь
Vg
b+ Va +
использовать мухт— и ■ х , то в
анализирующем скрещиovg Ь vg 1
вании
кроссоверные b+vg и b vg+ и некроссоверные
— b vg и b+vg+ особи появятся с той же частотой
(соответственно 17 и 83%), что и в первом
случае.
Результаты этих опытов
показывают, что сцепление генов
действительно существует, и лишь в
известном проценте случаев оно нарушается
вследствие кроссинговера. Отсюда и был
сделан вывод, что между гомологичными
хромосомами может осуществляться
взаимный обмен идентичными участками,
в результате чего гены, находящиеся в
этих участках парных хромосом, перемещаются
из одной гомологичной хромосомы в
другую. Отсутствие перекреста (полное
сцепление) между генами представляет
исключение и известно лишь у гетерогаметного
пола двух видов — дрозофилы и
шелкопряда.
Открытое Морганом
сцепленное наследование признаков
получило название закона сцепления
Моргана.
Наследование при плейотропном
действии гена. Однако совместное
наследование признаков при отсутствии
рекомбинантов может быть обусловлено
плейотропным действием одного гена.
Например, у кроликов доминантный ген R
определяет нормальную длину шерсти и
прямые усы, а рецессивная аллель г —
короткую шерсть и извитые усы. При
анализирующем скрещивании гетерозиготы
Rr любого пола с рецессивной гомозиготой
rr в Fb будут появляться в отношении 1 : 1
нор.иальношерстные кролики с прямыми
усами Rr и короткошерстные с извитыми
усами гг. Таким образом, в данном случае
наблюдается картина полного сцепления
признаков за счет плейотропии, но не
сцепления генов.
55.
Наследование
сцепленных признаков у дрозофилы: / —
в случае отсутствия кроссинговера
(гетерозиготный самец F,); 2— в случае
наличия кроссинговера (гетерозиготная
самка F,); В Fb изображены только самки,
так как самцы тех же фенотипов; Ь* —
серая; Ь — темная окраска тела; og* —
нормальные; vg —
зачаточные крылья.
генов
нарушается за счет того, что гены в
гомологичных хромосомах поменялись
местами благодаря кроссинговеру.
Гаметы
с хромосомами, претерпевшими кроссинговер,
называют кроссоверными, а не претерпевшие
— некроссоверными. Соответственно
организмы, возникшие от сочетания
кроссоверных гамет гибрида с гаметами
анализатора, называют кроссоверами или
рекомбинантами, а возникшие от сочетания
некроссоверных гамет с гаметами
анализатора — некроссоверными или
нерекомбинантными.
Закон сцепления
Моргана. При анализе расщепления в
случае кроссинговера обращает на себя
внимание определенное количественное
отношение кроссоверных и некроссоверных
классов. Обе исходные родительские
комбинации признаков, образовавшиеся
из некроссоверных гамет, оказываются
в потомстве анализирующего скрещивания
в равном количественном отношении. В
указанном опыте с дрозофилой тех и
других особей было примерно по 41,5%- В
сумме некроссоверные мухи составили
83% от общего числа потомков. Два
кроссоверных класса по числу особей
также одинаковы, и сумма их равна
17%.
Частота кроссинговера не зависит
от аллельного состояния генов, участвующих
в скрещивании. Если в качестве родителей
Ь
Vg
b+ Va +
использовать мухт— и ■ х , то в
анализирующем скрещиovg Ь vg 1
вании
кроссоверные b+vg и b vg+ и некроссоверные
— b vg и b+vg+ особи появятся с той же частотой
(соответственно 17 и 83%), что и в первом
случае.
Результаты этих опытов
показывают, что сцепление генов
действительно существует, и лишь в
известном проценте случаев оно нарушается
вследствие кроссинговера. Отсюда и был
сделан вывод, что между гомологичными
хромосомами может осуществляться
взаимный обмен идентичными участками,
в результате чего гены, находящиеся в
этих участках парных хромосом, перемещаются
из одной гомологичной хромосомы в
другую. Отсутствие перекреста (полное
сцепление) между генами представляет
исключение и известно лишь у гетерогаметного
пола двух видов — дрозофилы и
шелкопряда.
Открытое Морганом
сцепленное наследование признаков
получило название закона сцепления
Моргана.
Наследование при плейотропном
действии гена. Однако совместное
наследование признаков при отсутствии
рекомбинантов может быть обусловлено
плейотропным действием одного гена.
Например, у кроликов доминантный ген R
определяет нормальную длину шерсти и
прямые усы, а рецессивная аллель г —
короткую шерсть и извитые усы. При
анализирующем скрещивании гетерозиготы
Rr любого пола с рецессивной гомозиготой
rr в Fb будут появляться в отношении 1 : 1
нор.иальношерстные кролики с прямыми
усами Rr и короткошерстные с извитыми
усами гг. Таким образом, в данном случае
наблюдается картина полного сцепления
признаков за счет плейотропии, но не
сцепления генов.
Основоположник теории Томас Гент Морган, американский генетик, нобелевский лауреат, выдвинул гипотезу об ограничении законов Менделя.
В экспериментах он использовал плодовую мушку-дрозо-филу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков.
Морган и его ученики установили следующее:
1. Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно.
2. Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей.
3. Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссин-говера возникают гаметы, хромосомы которых содержат новые комбинации генов.
4. Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации.
5. Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности. Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами.
Пример сцепленного наследования:
Vg — нормальные крылья дрозофилы;
vg — зачаточные крылья;
ВВ — серая окраска тела;
bb — темная окраска тела.
Запись в хромосомном выражении:
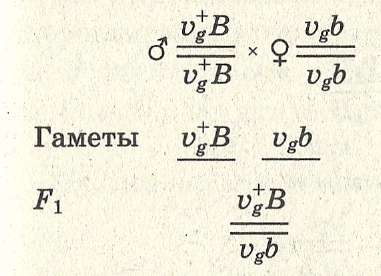
В данном случае правило единообразия гибридов первого поколения соблюдается. В соответствии со вторым и третьим законами Менделя следовало ожидать при последующем анализирующем скрещивании по 25% каждого из возможных фенотипов (серых, длиннокрылых мух, серых короткокрылых, черных длиннокрылых и черных коротко-крылых). Однако опыты Моргана не дали таких результатов. При скрещивании рецессивной по обоим признакам самки VgVgbb с гибридным самцом из F1 образовалось 50% серых мух с короткими крыльями и 50% мух с черным телом и длинными крыльями:
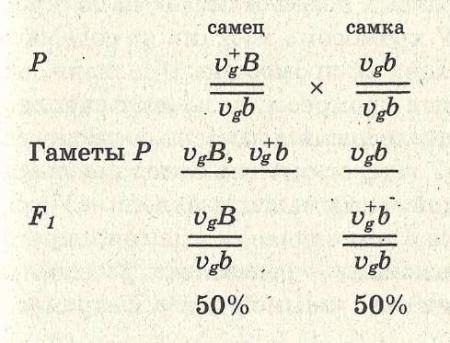
Если же скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в образуется потомство: 41,5% — серых с короткими крыльями, 41,5% — черных с длинными крыльями, 8,5% — серых с длинными крыльями, 8,5% — черных с короткими крыльями.

Данные результаты свидетельствуют о наличии сцепления генов и кроссинговере между ними. Так как в потомстве от второго скрещивания было получено 17% рекомбинант-ных особей, то расстояние между генами Vg и В равно 17%, или 17 морганидам.
Наследование, сцепленное с полом
Хромосомные наборы разных полов отличаются по строению половых хромосом. У-хромосома мужчин не содержит многих аллелей, имеющихся в Х-хромосоме. Признаки, определяемые генами половых хромосом, называют сцепленными с полом. Характер наследования зависит от распределения хромосом в мейозе. У гетерогаметных полов признаки, сцепленные с Х-хромосомой и не имеющие аллеля в У-хро-мосоме, проявляются даже в том случае, когда ген, определяющий развитие этих признаков, — рецессивен. У человека У-хромосома передается от отца к сыновьям, а Х-хромосо-ма — к дочерям. Вторую хромосому дети получают от матери. Это всегда Х-хромосома. Если мать несет патологический рецессивный ген в одной из Х-хромосом (например, ген дальтонизма или гемофилии), но при этом сама не больна, то она является носительницей. В случае передачи этого гена сыновьям они могут родиться с данным заболеванием, ибо в У-хромосоме нет аллеля, подавляющего патологический ген. Пол организма определяется в момент оплодотворения и зависит от хромосомного набора образовавшейся зиготы. У птиц гетерогаметными являются самки, а гомогаметными — самцы. У пчел половых хромосом вообще нет. Самцы гаплоидны. Самки пчел диплоидны.
Пример наследования, сцепленного с полом

Основные положения хромосомной теории наследственности:
• каждый ген имеет в хромосоме определенный локус (место);
• гены в хромосоме расположены в определенной последовательности;
• гены одной хромосомы сцеплены, поэтому наследуются преимущественно вместе;
• частота кроссинговера между генами равна расстоянию между ними;
• набор хромосом в клетках данного типа (кариотип) является характерной особенностью вида.
Основоположник теории Томас Гент Морган, американский генетик, нобелевский лауреат, выдвинул гипотезу об ограничении законов Менделя.
В экспериментах он использовал плодовую мушку-дрозо-филу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков.
Морган и его ученики установили следующее:
1. Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно.
2. Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей.
3. Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссин-говера возникают гаметы, хромосомы которых содержат новые комбинации генов.
4. Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации.
5. Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности. Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами.
Пример сцепленного наследования:
Vg — нормальные крылья дрозофилы;
vg — зачаточные крылья;
ВВ — серая окраска тела;
bb — темная окраска тела.
Запись в хромосомном выражении:
В данном случае правило единообразия гибридов первого поколения соблюдается. В соответствии со вторым и третьим законами Менделя следовало ожидать при последующем анализирующем скрещивании по 25% каждого из возможных фенотипов (серых, длиннокрылых мух, серых короткокрылых, черных длиннокрылых и черных коротко-крылых). Однако опыты Моргана не дали таких результатов. При скрещивании рецессивной по обоим признакам самки VgVgbb с гибридным самцом из F1 образовалось 50% серых мух с короткими крыльями и 50% мух с черным телом и длинными крыльями:
Если же скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в образуется потомство: 41,5% — серых с короткими крыльями, 41,5% — черных с длинными крыльями, 8,5% — серых с длинными крыльями, 8,5% — черных с короткими крыльями.
Данные результаты свидетельствуют о наличии сцепления генов и кроссинговере между ними. Так как в потомстве от второго скрещивания было получено 17% рекомбинант-ных особей, то расстояние между генами Vg и В равно 17%, или 17 морганидам.