

1.1.Биологические основы функционирования нервных клеток
ИНС представляют собой попытку использования процессов, происходящих в нервных системах живых существ для создания новых информационных технологий. Основным элементом нервной системы является нервная клетка, сокращенно называемая нейроном. Как и у любой другой клетки, у нейрона имеется тело, называемое сомой, внутри которого располагается ядро, а наружу выходят многочисленные отростки – тонкие, густо ветвящиеся дендриты, и более толстый, расщепляющийся на конце аксон (рис. 1.1). Входные сигналы поступают в клетку через синапсы, выходной сигнал передается аксоном через его нервные окончания (колатералы) к синапсам других нейронов, которые могут находиться как на дендритах, так и непосредственно на теле клетки.
Передача сигналов внутри нервной системы – очень сложный электрохимический процесс, основанный на выделении особых химических веществ – нейромедиаторов, которые образуются под действием поступающих от синапсов раздражителей и воздействуют на клеточную мембрану, вызывая изменение ее энергетического потенциала пропорционально количеству попавшего нейромедиатора. Поскольку синапсы отличаются размерами и концентрацией выделяемого нейромедиатора, то импульсы одинаковой величины, поступающие по различным синапсам, могут возбуждать нервную клетку в разной степени. Мерой возбуждения клетки считается уровень поляризации ее мембраны, зависящий от суммарного количества нейромедиатора по всем синапсам, причем синапсы могут оказывать как возбуждающее, так и тормозящее действие. Если баланс возбуждений и торможений невелик или отрицателен, т.е. ниже порога срабатывания клетки, то выходной сигнал не образуется, в противном случае значение выходного сигнала лавинообразно нарастает, принимая характерный вид нервного импульса (рис. 1.2), передаваемого аксоном на подключенные к нему нейроны. Амплитуда этого импульса не зависит от степени превышения порога, т.е. клетка действует по принципу «всё или ничего». После выполнения своей функции нейромедиатор удаляется путем либо всасывания или разложения клеткой, либо выбросом за пределы синапса.
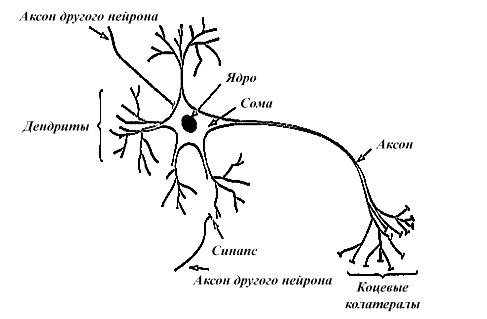
Рис. 1.1. Упрощенная структура биологической нервной клетки
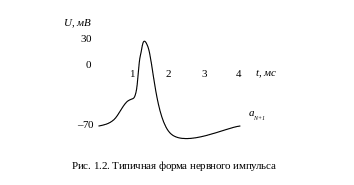
После генерации нервного импульса в клетке наступает период абсолютной рефрактерности tp, когда нейрон теряет способность вырабатывать очередной сигнал даже при сильном возбуждении. По окончании tp, наступает период относительной рефракции t0, за который порог срабатывания возвращается к равновесному значению. В это время клетку можно активировать, но только прикладывая более сильные возбуждения. В естественных процессах, как правило, выполняется t0>>tp.
Количество взаимодействующих друг с другом нервных клеток чрезвычайно велико. Считается, что человеческий мозг содержит порядка 1011 нейронов, соединенных между собой примерно 1015 связями (до 104 на один нейрон). Каждый нейрон имеет свои веса связей и свое пороговое значение, определяемые его расположением и выполняемой функцией. Громадное число нейронов и межнейронных связей приводит к тому, что ошибки в срабатывании отдельных нейронов (до 10% от общего числа) слабо влияют на конечный результат, несколько ухудшая, может быть, его точность. Вторая важная особенность нервных систем – высокая скорость их функционирования, несмотря на невысокое быстродействие (~10–3 с) отдельных клеток, что достигается благодаря параллельной обработке информации лавинообразно нарастающим количеством элементарных вычислительных ячеек – нейронов. Если бы удалось создать вычислительную систему с аналогичной степенью параллельности независимых операций при существующих в СБИС скоростях их выполнения (~10–9 с на такт), ее потенциальные возможности трудно даже представить.