

2. Задачі на дигібридне та полігібридне схрещювання не зцеплених генів.
При дигібридному схрещюванні просліджуеться схрещювання по двум незалежним ознакам, при полігібридному схрещюванні_по декільком_не зцепленим ознакам.
Задача № 7. У людини деякі форми короткозорості домінують над нормальним зіром , а колір каріх очей над блакитними. Гени обох пар знаходяться в різних хромосомах. Яке потомство слід очикувати від браку гетерозіготних по обом ознакам батьків?
Рішення:
З умови обидва батьки гетерозіготні з обох ознак тобто «АаВв»
А - алель яка визначає короткозорість;
а - алель яка визначає норм. зір;
В - алель яка визначає карі очі ;
в - алель яка визначає блакиткі очі.
АаВв
АВ ; Ав ; аВ ; ав; Короткзор. кароокі.
Р АаВв Х
Гамети: АВ ; Ав ; аВ ; ав;
Фенотипи: Короткзор.кароокі
Жен Муж |
АВ |
Ав |
аВ |
ав |
АВ |
ААВВ Корот. Кароок |
ААВв Корот. Кароок |
АаВВ Корот. Кароок |
АаВв Корот. Кароок |
Ав |
ААВв Корот. Кароок |
ААвв Корот. голуб |
АаВв Корот. Кароок |
Аавв Корот. блакит . |
аВ |
АаВВ Корот. Кароок |
АаВв Корот. Кароок |
ааВВ норма Кароок |
ааВв норма Кароок |
ав
|
АаВв Корот. Кароок |
Аавв Корот. блакит |
ааВв норма Кароок |
аавв Норма блакит |
Відповідь : Можливі варіанти -9:3:3:1; короткозорих карооких - 9 частин (ААВВ; ААВв; АаВВ; АаВв;) короткозорих блакитнооких 3 частини (ААвв; Аавв;), Нормальний зір з карими очима - 3 частини ( ааВВ; ааВв;), Нормальний зір з блакитними очима -1 частина (аавв).
Аналіз: Тут має прояв ІІІ закон Г. Менделя
Задача № 8. У собак чорний окрас хутра визначаеться геном - В, коришневий -в , сплошний окрас - С , пегій (строкатий) — с. Коришневий самец и чорно-пега самиця мають 5 цуценят: 1 чорний, 1 коришневий , 1 чорнопегій, 2 коришневопегі . Які генотипи батьків?
Рішення : з умови видно , що батьки з рецесивної ознаки є гомозіготними. З 2 ознаки вони можут бути як гомо- так і гетерозіготними. Їх генотипи будуть такими : самець - (ввС?) самиця - (В?сс ). Щоб визначити , необхідно розрахувати у потомків генотипи . Особо звертають увагу на гомозіготність алелів. У цуценят ( В?С? - чорний, ввС?- коришневий пыдходить для визначення тому як алель (вв) в равный мыры йде выд обох батькыв, виходить що одне (в) от самця, а друге (в) від самиці, тобто у самиці замість ? можно поставити (в) — Ввсс ! Далі : В?сс — чорно-пегий - теж саме можно розрахувати і по (сс) виходить у самця замість ? — (с) та отримуем генотип - (ввСс )) .Тепер перепровіряем :
Р самець ввСс х самиця Ввсс
Гамети: вС; вс; Вс; вс; (тількі такі види)
Генотипи: ВвСс : ввСс : Ввсс : ввсс .
Фенотипи: Чорний коришневий чорнопегій коришневопегій.
Що зпівпадає з умовою !
Відповідь : Генотипи у коришневого самця - (ввСс) у чорнопегої самиці - (Ввсс) .
Аналіз : Тут має прояв ІІІ закон Г. Менделя
Розв’яжіть задачі : Задача № 9. В родині де батьки гарно чули та мали один рівне волосся , а другий хвилясте, народилась дитина глуха з рівним волоссям. Їх друга дитина гарно чула та мала хвилясте волосся. Яка верогідність народження глухих дітей з хвилястим волоссям? Відомо, що ген хвилястості волосся домінує над рівним, а глухота рецесивна ознака та обидві пари генів знаходяться в різних хромосомах.
Задача №10.У людини деякі форми короткозорості домінують над нормальним зором, а колір
карих очей над блакитними. Гени обох пар вільні. Якого потомства можна
очикувати від браку гетерозіготного чоловіка з жінкою яка має блакитні очі та
нормальний зір ?
Задача № 11.Жінка має вільну мочку вуха (домінантна ознака) та рівне підборіддя , а чоловік прирощину мочку та треугольну ямку на підборідді (домінантна ознака). Син має вільну мочку вуха та треугольну ямку на підборідді, а дочка має тіж ознаки, що у матері. Напишіть можливі генотипи батьків та їх дітей.
Задача № 12 ( на тригібрідное скрещивание - полігібрідне) . Полідактилія, короткозорість та відсутність малих кутніх зубів спадкуються як домінантні аутосомні ознаки. Гени всіх 3 ознак знаходяться в різних парах хромосом. Яка верогідність народження дітей без аномалій в родині де обидва батьки страждають всіма з цих 3 недоліків, та є ще й гетерозіготами з усіх 3 пар генів ?
Література
Основна : с. 153 – 160.
додаткова: Т.1 с.234 – 254.
Медична біологія
Лекція № 4. Хромосомна теорія спадковості. Генетика статі.
1. Хромосомний рівень організації спадковості.
2. Аутосоми та гетерохромосоми.
3. Хромосоми людини. Класифікація. Кариотип людини.
4. Генетика статі .
5. Механізми генетичного визначення статі .
Самостійна робота № 5. Розв’язування задач на зчеплене успадкування та взаємодію
генів.
Самостійна робота № 6. Генетичні карти. Методи картування хромосом людини. Сучасний стан
дослідження генома людини. Генетична небезпека забруднення середовища.
Поняття про антимутаген та комутаген. Генна інженерія. Біотехнології.
Поняття про генну терапію.
1. Хромосомний рівень організації спадковості.
Х ромосома
- це ниткоподібні щільні тільця, видимі
у світловий мікроскоп тільки впродовж
поділу клітини. Вони утворюються
в результаті ущільнення і спіралізації
хроматину.
Довжина хромосом залежить від кількості
ДНК і білків, а також від ступеня
скручування хроматину. На різних
ділянках однієї і тієї ж хромосоми
спіралізація, компактність її основних
елементів неоднакові,
з цим пов'язана різна інтенсивність
забарвлення окремих ділянок хромосоми.
Ділянки,
що інтенсивно сприймають барвники,
одержали
назву гетерохроматичних
(утворених
із гетерохроматину),
вони навіть у період між поділом клітин
залишаються компактними, видимими у
світловий
мікроскоп. Ділянки, що слабко забарвлюються,
деконденсуються між поділами клітин,
одержали
назву еухроматичних
(утворених
із еухроматину).
Встановлено, що еухроматин містить у
собі
активні гени, а гетерохроматин виконує
переважно
структурну функцію. Він знаходиться в
інтенсивно спіралізованому стані.
Гетерохроматин займає однакові
ділянки в гомологічних хромосомах:
утворює
ділянки, що прилягають до центромери і
такі, що знаходяться
на кінцях хромосом. Втрата ділянок
ге-терохроматину
або мутація може не відбиватися на
життєдіяльності
клітини. Розрізняють також факультативний
гетерохроматин. Він
виникає при спіралізації
та інактивації однієї з двох гомологічних
Х-хромосом.
Таким чином утворюється тільце Барра
(статевий
хроматин). Його утворює одна з двох
Х-хромосом
у жіночих особин ссавців і людини.
ромосома
- це ниткоподібні щільні тільця, видимі
у світловий мікроскоп тільки впродовж
поділу клітини. Вони утворюються
в результаті ущільнення і спіралізації
хроматину.
Довжина хромосом залежить від кількості
ДНК і білків, а також від ступеня
скручування хроматину. На різних
ділянках однієї і тієї ж хромосоми
спіралізація, компактність її основних
елементів неоднакові,
з цим пов'язана різна інтенсивність
забарвлення окремих ділянок хромосоми.
Ділянки,
що інтенсивно сприймають барвники,
одержали
назву гетерохроматичних
(утворених
із гетерохроматину),
вони навіть у період між поділом клітин
залишаються компактними, видимими у
світловий
мікроскоп. Ділянки, що слабко забарвлюються,
деконденсуються між поділами клітин,
одержали
назву еухроматичних
(утворених
із еухроматину).
Встановлено, що еухроматин містить у
собі
активні гени, а гетерохроматин виконує
переважно
структурну функцію. Він знаходиться в
інтенсивно спіралізованому стані.
Гетерохроматин займає однакові
ділянки в гомологічних хромосомах:
утворює
ділянки, що прилягають до центромери і
такі, що знаходяться
на кінцях хромосом. Втрата ділянок
ге-терохроматину
або мутація може не відбиватися на
життєдіяльності
клітини. Розрізняють також факультативний
гетерохроматин. Він
виникає при спіралізації
та інактивації однієї з двох гомологічних
Х-хромосом.
Таким чином утворюється тільце Барра
(статевий
хроматин). Його утворює одна з двох
Х-хромосом
у жіночих особин ссавців і людини.
Кожна молекула ДНК, що утворює хромосому, крім кодуючих послідовностей, повинна містити центромеру, два теломери і точки початку реплікації. Теломерні послідовності запобігають укороченню хромосом, що без них відбувалося б при кожному циклі реплікації ДНК. Центромери служать для вибудовування молекул ДНК на мітотичному веретені під час мітозу. Точки початку реплікації потрібні для формування реплікаційних вилок у 5-фазі.
С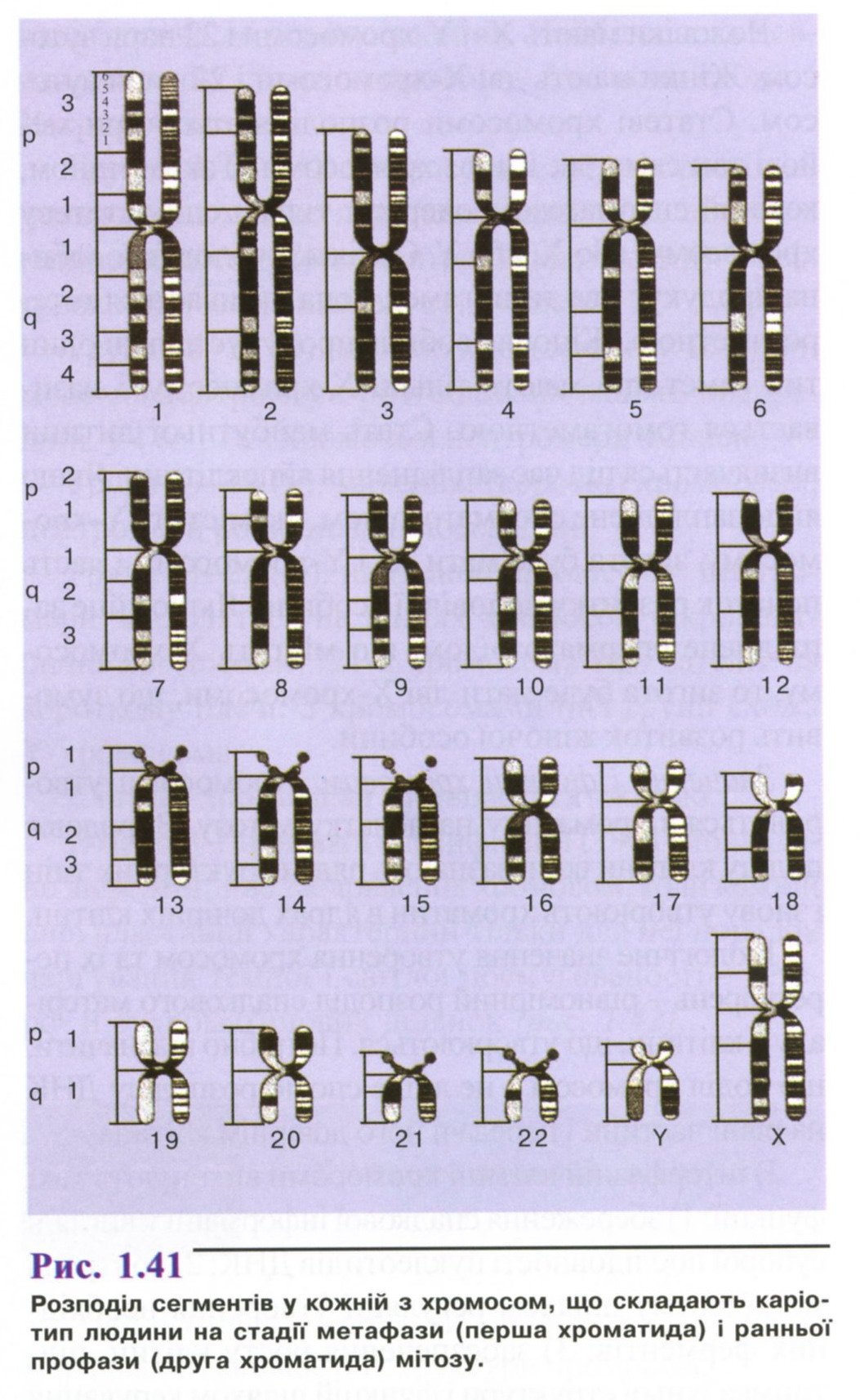 труктура
метафазної хромосоми.
Всі хромосоми
під час метафази складаються із двох
хроматид, що утворені з максимально
спіралізованого хроматину
(рис.
1.40). Кожна
хроматида - це зв'язана
з гістонами одинарна двониткова
суперспіралізована
ДНК. Дві дочірні молекули ДНК, що
знаходяться
у двох хроматидах, утримуються разом у
ділянці
центромери за допомогою нереплікованого
сегмента ДНК. Будова хромосоми на різних
ділянках неоднакова. У хромосомах
розрізняють первинну
перетяжку, що
поділяє хромосому на два плеча.
Первинна перетяжка (центромера) - найбільш
спіралізована
частина хромосоми. Центромера являє
собою загальну, нерепліковану ділянку
ДНК. На
ній розташовуються спеціальні білки,
що утворюють
кінетохори,
до яких при розподілі генетичного
матеріалу
прикріплюються нитки веретена. Це сприяє
поділу
дочірніх хроматид під час анафази. Місце
розташування
первинної перетяжки в кожній парі
хромосом
індивідуальне і стале, що зумовлює,
головним
чином, її форму.
труктура
метафазної хромосоми.
Всі хромосоми
під час метафази складаються із двох
хроматид, що утворені з максимально
спіралізованого хроматину
(рис.
1.40). Кожна
хроматида - це зв'язана
з гістонами одинарна двониткова
суперспіралізована
ДНК. Дві дочірні молекули ДНК, що
знаходяться
у двох хроматидах, утримуються разом у
ділянці
центромери за допомогою нереплікованого
сегмента ДНК. Будова хромосоми на різних
ділянках неоднакова. У хромосомах
розрізняють первинну
перетяжку, що
поділяє хромосому на два плеча.
Первинна перетяжка (центромера) - найбільш
спіралізована
частина хромосоми. Центромера являє
собою загальну, нерепліковану ділянку
ДНК. На
ній розташовуються спеціальні білки,
що утворюють
кінетохори,
до яких при розподілі генетичного
матеріалу
прикріплюються нитки веретена. Це сприяє
поділу
дочірніх хроматид під час анафази. Місце
розташування
первинної перетяжки в кожній парі
хромосом
індивідуальне і стале, що зумовлює,
головним
чином, її форму.
Кінці плечей хромосом одержали назву теломерів. Вони містять тисячі повторюваних послідовностей, наприклад, ТТАТТГ. Це генетичне неактивні спіралізовані ділянки, що перешкоджають з'єднанню хромосом між собою або з їх фрагментами. Позбавлена теломери хромосома виявляється "липкою" і легко з'єднується з такими ж ділянками інших хромосом. Отже, теломери зберігають хромосому як індивідуальну дискретну одиницю, забезпечують її індивідуальність.
Хромосома на стадії ранньої профази набагато довша і тонша, ніж метафазні хромосоми, тому в них можна розпізнати більше сегментів (рис. 1.41). Показані на рисунку сегменти виявляються шляхом фарбування препаратів барвниками, що зв'язуються переважно з А-Т - насиченими послідовностями ДНК. На рисунку представлено тільки по одній хромосомі з кожної гомологічної пари.
2. Аутосоми та гетерохромосоми.
Аутосоми та гетерохромосоми. Було відмічено, що практично в будь-якому каріотипі існує одна пара хромосом, що істотно відрізняється від інших. У самок ця пара була однакова, але різна в представників протилежної статі (самців). Хромосома, що була присутня двічі в парі в каріотипі самки й один раз у самця, була названа як Х-хромосома. Друга хромосома в самця була названа У-хромосомою. X- і У-хромосоми називаються статевими хромосомами, або гетерохромосомами. Інші хромосоми з даного набору, що є однаковими для обох статей, називаються аутосомами.
Чоловіки мають X- і У-хромосоми і 22 пари аутосом. Жінки мають дві Х-хромосоми і 22 пари аутосом. Статеві хромосоми розподіляються при мейозі так само, як і інші хромосоми. Таким чином, кожний сперматозоїд одержує тільки одну статеву хромосому: або X, або У. Оскільки чоловіча особина продукує два типи гамет, вона називається гете-рогаметною. Жіноча особина продукує тільки один тип гамет, що мають тільки Х-хромосому, і називається гомогаметною. Стать майбутньої дитини визначається під час запліднення яйцеклітини. Якщо яйце запліднене сперматозоїдом, що містить У-хро-мосому, зигота буде мати X- і У-хромосоми і дасть початок розвитку чоловічої особини. Якщо яйце запліднене сперматозоїдом, що містить Х-хромосому, то зигота буде мати дві Х-хромосоми, що зумовить розвиток жіночої особини.
Значення і функції хромосом. Хромосоми утворюються з хроматину на початку мітозу. Впродовж поділу клітини вони зазнають ряду структурних змін і знову утворюють хроматин в ядрах дочірніх клітин.
Біологічне значення утворення хромосом та їх перетворень - рівномірний розподіл спадкового матеріалу в клітини, що утворюються. Потрібно відзначити, що поділ хромосом - це лише спосіб розподілу ДНК на рівні частини і передачі його дочірнім клітинам.
В інтерфазній клітині хромосоми виконують такі функції: 1) збереження спадкової інформації у вигляді суворої послідовності нуклеотидів ДНК; 2) контроль метаболізму шляхом регуляції утворення необхідних ферментів; 3) забезпечення росту клітин, підтримка їхньої структури і функцій шляхом керування синтезом структурних білків; 4) контроль клітинного диференціювання під час розвитку; 5) забезпечення умов подвоєння ДНК.
У клітинах, що діляться, утворюються візуально помітні хромосоми, що необхідні для таких функцій: 1) компактизація (ущільнення) спадкового матеріалу в тисячі разів, при цьому генетичний матеріал "консервується" і втрачає спроможність до експресії; 2) утворення структур, зручних для маніпуляцій при поділі (метафазна хромосома, що складається з двох хроматид); 3) рівномірний розподіл компактизованого неактивного генетичного матеріалу між дочірніми клітинами у вигляді окремих хроматид; 4) декомпактизація (розпушення) спадкового матеріалу й утворення активного інтерфаз-ного хроматину.
У 50-х роках XX ст. стало можливим спостерігати і було описано кожну хромосому людини у вигляді самостійної одиниці. Загальна довжина молекули ДНК у хромосомі людини (середньої за розміром) досягає 4 см, а сумарна довжина цих молекул у клітині з диплоїдним (подвійним) набором -близько 180 см. У 1956 р. Дж. Тийо, А. Леван установили, що у людини хромосомний набір складається з 46 хромосом. Через три роки були відкриті хромосомні хвороби.
В основу ідентифікації хромосом покладено такі ознаки: загальна довжина хромосоми, розміщення центромери, вторинна перетяжка та ін. Але за цими ознаками не завжди можна розрізнити індивідуальні особливості хромосоми. Останнім часом проводять диференційне забарвлення хромосом, яке дозволяє чітко розрізнити кожну з хромосом.
Сукупність хромосом клітини, яка характеризується їх числом, розмірами і формою, називається каріотипом. Ідіограма- це систематизований каріотип, коли хромосоми розташовуються в порядку зменшення їх довжини.
3. Хромосоми людини. Класифікація. Кариотип людини.
За Денверською класифікацією (Денвер, США, 1960 р.) всі аутосоми людини поділяються на 7 груп залежно від довжини хромосом і розміщення центромери. Кожна група позначається латинськими літерами від А до О. Крім того, всі аутосоми в порядку зменшення нумеруються (від 1 до 22). Визначені також статеві хромосоми X і У.
Група 1-3 (А): великі хромосоми, які чітко відрізняються одна від одної; центромери розташовані посередині.
Група 4-5 (В): великі хромосоми, які мало відрізняються одна від одної; центромери зміщені до одного з кінців хромосоми.
Група 6-12 (С): хромосоми середніх розмірів, мало різняться між собою; центромери розташовані ближче до одного з кінців. Найбільша за довжиною з цієї групи хромосом - 6-а, вона схожа з Х-хромосомою.
Група 13-15 (О): хромосоми середніх розмірів; центромери майже повністю зміщені до одного з кінців хромосоми (акроцентричні хромосоми). У всіх трьох хромосом виявлені супутники.
Група 16-18 (Е): короткі хромосоми; у 16-ї хромосоми центромера розташована майже посередині, у 17-ї і 18-ї хромосом центромери зміщені.
Група 19-20 (Р): маленькі (короткі) хромосоми; центромери розташовані посередині.
Група 21-22 (О): найменші хромосоми; центромери знаходяться на кінцях хромосом (акроцентричні хромосоми). 21-а хромосома має сателіт на короткому плечі. З хромосомами цієї групи схожа У-хромосома.
Статеві хромосоми виділяються окремо.
В основу Паризької класифікації (1971) покладено диференційне забарвлення хромосом, коли кожній парі властивий характерний тільки для неї порядок чергування темної і світлої посмугованості - гетеро- й еухроматинових ділянок (рис. 1.41).
Хромосомний аналіз - це дослідження й аналіз морфології, кількості і структури хромосом.
Розміри і форма хромосом. Розміри хромосом варіюють від одного біологічного виду до іншого. Хромосоми різних організмів на стадії метафази мають довжину від 0,1 до 33,0 мкм і товщину від 0,2 до 2,0 мкм. Хромосоми рослин мають більші розміри, ніж хромосоми тварин. Хромосоми різних пар однієї і тієї ж клітини різняться за розміром. Розмір хромосом людини: в середньому 1,5 мкм у товщину і 10,0 мкм у довжину.
Форма хромосом визначається за відносним положенням центромери (первинної перетяжки). На підставі цього розрізняють такі форми хромосом : 1) метацентрична - хромосома має Х-подібну форму, при якій центромера знаходиться всередині так, що плечі є рівними за довжиною; 2) субметацентрична - хромосома має Х-форму з центромерою, віддаленою від середньої точки так,
що її плечі є нерівними за довжиною; 3) акроцентрична - центромера розташована дуже близько до одного з кінців хромосоми, тобто вона має плечі, що суттєво відрізняються за розмірами, маленькі плечі часто мають супутники.
Каріотип людини має всі різновиди хромосом.
Правила хромосом. 1. Специфічність набору хромосом для кожного виду. Рослини і тварини мають сталий набір хромосом у кожній соматичній клітині. Диплоїдний набір хромосом (2п)
для людини - 46, для дрозофіли - 8, для коня - 66, шимпанзе - 48, собаки - 78 і т. д. Гаплоїдний набір (п) для людини - 23, дрозофіли - 4 і т. д. Гамети містять тільки одинарний набір хромосом. Число хромосом часто використовується для ідентифікації виду.
Число хромосом не залежить від рівня організації виду і не завжди вказує на філогенетичну спорідненість. Однакова кількість хромосом може зустрічатися в далеких один від одного видів і дуже відрізнятися у близьких. Однак істотні відмінності й специфічність виявляються в тому, що кожний вид має у своєму наборі хромосоми певної форми і розмірів. А головне - усі хромосоми мають свій унікальний набір генів, що визначають розвиток особин тільки певного виду.
Хромосомний комплекс виду з усіма його особливостями - числом хромосом, формою, наявністю видимих у світловий мікроскоп деталей будови окремих хромосом - називається каріотипом.
2. Парність хромосом. Кожна хромосома соматичних клітин має аналогічну собі хромосому. Хромосоми з такої пари мають однаковий розмір, форму і склад генів. Пари хромосом, що мають однакові гени або їх алелі, та контролюють альтернативні ознаки, називаються гомологічними. Гомологічні хромосоми однакові за розміром і формою. У них збігаються розміщення центромер, порядок розташування хро-момер і міжхромомерних ділянок та інші елементи будови. Негомологічні хромосоми мають зовсім інші характеристики.
Одна гомологічна хромосома надається від одного з батьків (батька), а інша від другого (матері). Генетична інформація, необхідна для розвитку організму, міститься тільки в повному комплекті всіх негомологічних хромосом (тобто в повному диплоїдному наборі хромосом).
Індивідуальність окремих пар хромосом. Кожна пара гомологічних хромосом індивідуума відрізняється від іншої пари за розміром, формою і генетичним складом. Наприклад, перша пара хромосом людини (метацентричні, найбільші, мають індивідуальну посмугованість) дуже відрізняється від 22-ї пари (акроцентричні, найменші, мають вторинну перетяжку і супутник) . Вони містять різні гени, що визначають розвиток різних ознак.
Безперервність хромосом. Це означає, що кожна дочірня хромосома походить від материнської хромосоми. В інтерфазі (3-періоді) відбувається
подвоєння ДНК і утворюються дві ідентичні дочірні молекули, що формують хромосому. Така хромосома складається з двох хроматид, що потім потрапляють у різні клітини внаслідок мітозу. У кожному наступному поділі цей цикл повторюється. Це забезпечує стабільність каріотипу організмів впродовж тисячоліть.
Таким чином, у послідовних генераціях клітин зберігається постійне число хромосом та їх індивідуальність внаслідок здатності хромосом до точної репродукції при поділі клітини. Отже, не тільки "кожна клітина від клітини", але і "кожна хромосома від хромосоми".
4. Хромосомна теорія спадковості (Т. X. Морган та ін.)
Доведено, що кількість спадкових ознак організму значно перевищує число хромосом гаплоїдного набору. Так, у гаплоїдному наборі класичного об'єкта генетичних досліджень - мухи-дрозофіли - є лише чотири хромосоми, але число спадкових ознак і відповідно генів, які їх визначають, безсумнівно, значно більше. Це означає, що в кожній хромосомі розміщено багато генів. Тож разом із ознаками, які успадковуються незалежно, повинні існувати і такі, що успадковуються зчеплено одна з одною, бо вони визначаються генами, розташованими в одній хромосомі. Такі гени утворюють групу зчеплення. Кількість груп зчеплення в організмів певного виду дорівнює кількості хромосом у гаплоїдному наборі (наприклад, у дрозофіли 1п=4, у людини 1п=23).
Експериментальні дослідження явища зчепленого успадкування провів видатний американський генетик Т. X. Морган зі своїми співробітниками, їхні результати обґрунтували запропоновану ними хромосомну теорію спадковості.
Слід зазначити, що Т. X. Морган, як і свого часу Г. Мендель, вдало обрав для досліджень муху-дрозофілу, яка згодом стала класичним об'єктом для генетичних експериментів. Дрозофіл легко утримувати в лабораторіях, вони мають значну плодючість, швидку зміну поколінь (за оптимальних умов утримання нове
покоління виникає кожні півтора-два тижні), невелике число хромосом, що спрощує спостереження.
Явище зчепленого успадкування Т. X. Морган встановив у такому досліді. Самців дрозофіли, гомозиготних за домінантними алелями забарвлення тіла (сіре) та формою крил (нормальні), схрестили із самками, гомозиготними за відповідними рецесивними алелями (чорне тіло - недорозвинені крила). Генотипи цих особин позначили відповідно ЕЕVV та ееvv. Усі гібриди першого покоління мали сіре тіло і нормальні крила, тобто були гетерозиготними за обома парами алелів (генотип - ЕеVV) .
Потім гібридів схрестили з особинами, гомозиготними за відповідними рецесивними алелями (аналізуюче схрещування).
Теоретично можна було очікувати два варіанти розщеплення. Якби гени, які зумовлюють забарвлення тіла та форму крил, містилися в негомологіч-них хромосомах, тобто успадковувалися незалежно, розщеплення мало бути таким: 25 % особин із сірим тілом і нормальними крилами, 25 % - із сірим тілом і недорозвиненими крилами, ще 25 % - з чорним тілом і нормальними крилами та 25 % - з чорним тілом і недорозвиненими крилами (тобто у співвідношенні -1:1:1:1). Якщо б ці гени розміщувалися в одній хромосомі і успадковувалися зчеплено, то було б отримано 50 % особин з сірим тілом і нормальними крилами та 50 % - з чорним тілом і недорозвиненими крилами (тобто 1:1).
Насправді 41,5 % особин мали сіре тіло і нормальні крила, 41,5 % - чорне тіло і недорозвинені крила, 8,5 % - сіре тіло і недорозвинені крила і 8,5 % -
чорне тіло і нормальні крила, тобто розщеплення наближувалося до співвідношення фенотипів 1:1 (як у разі зчепленого успадкування), але разом з тим проявилися всі чотири варіанти фенотипу (як у випадку незалежного успадкування).
На підставі цих даних Т. X. Моргай припустив, що гени, які визначають забарвлення тіла і форму крил, розташовані в одній хромосомі, але в процесі мейозу під час утворення гамет гомологічні хромосоми можуть обмінюватися ділянками, тобто має місце явище, яке дістало назву перехрест хромосом, або кросинговер.
Кросинговер - обмін ділянками гомологічних хромосом у процесі клітинного поділу, переважно в профазі першого мейотичного поділу, іноді в мітозі.
Дослідами Т. Моргана, К. Бріджеса і А. Стертеванта було показано, що немає абсолютно повного зчеплення генів, за якого гени передавались би завжди разом. Імовірність того, що два гени, локалізовані в одній хромосомі, не розійдуться в процесі мейозу, коливається в межах 1-0,5. У природі переважає неповне зчеплення, зумовлене перехрестом гомологічних хромосом і рекомбінацією генів .
Цитологічна картина кросинговеру була вперше описана датським вченим Ф. Янсенсом.
Кросинговер проявляється тільки тоді, коли гени знаходяться в гетерозиготному стані (АВ/аЬ). Якщо гени в гомозиготному стані (АВ/АВ або аЬ/аЬ), обмін ідентичними ділянками не дає нових комбінацій генів у гаметах і в поколінні.
Частота (відсоток) перехресту між генами залежить від відстані між ними: чим далі вони розташовані один від одного, тим частіше відбувається кросинговер. Т. Морган запропонував відстань між генами вимірювати кросинговером у відсотках, за формулою:
n 1
-------- . 100 = % крoсинговеру
n
де п - загальне число особин в F;
п1 - сумарне число кросоверних особин.
Відрізок хромосоми, на якому здійснюється 1 % кросинговеру, дорівнює одній морганіді (умовна міра відстані між генами). Частоту кросинговеру використовують для того, щоб визначити взаємне розміщення генів і відстань між ними. Для побудови генетичної карти людини користуються новими технологіями. Побудовані цитогенетичні карти хромосом.
Кросинговер призводить до нового поєднання генів, що викликає зміну фенотипу. Крім того, він поряд з мутаціями є важливим фактором еволюції організмів.
Зазначені докази були покладені в основу хромосомної теорії спадковості:
1. Гени розміщаються у хромосомах по довжині в лінійному порядку; різні хромосоми містять неоднакове число генів; набір генів кожної з негомологічних хромосом - унікальний.
Алельні гени займають певні й ідентичні локуси (місця) гомологічних хромосом.
Гени, розташовані в одній хромосомі, утворюють групу зчеплення, завдяки чому має місце зчеплення деяких ознак, які разом (зчеплено) передаються нащадкам. Кількість груп зчеплення дорівнює гаплоїдному набору хромосом. Зчеплення не абсолютне.
Під час мейозу, який відбувається тільки при утворенні гамет, диплоїдне число хромосом зменшується вдвічі (гаплоїдне число). Це відповідає закону розщеплення, за яким генетичний матеріалобох батьків по-різному комбінується в гаметах.
Згідно з законом незалежного розподілу батьківські й материнські набори незчеплених генів розщеплюються незалежно один від одного. Якщо незчеплені гени розташовані в різних хромосомах, то під час мейозу материнські й батьківські хромосоми повинні розподілитися між гаметами випадково.
Між генами гомологічних батьківських і материнських груп зчеплення можуть відбуватися, завдяки кросинговеру, реципрокній рекомбінації. Цьому відповідає утворення хіазм під час кон'югації гомоло гічних хромосом у мейозі (генетичний кросинговер).
Сила зчеплення між генами обернено пропорційна відстані між ними. Чим ближче розташовані гени в одній хромосомі, тим сильніше їх зчеплення, тим менше виникатиме рекомбінацій між ними, і навпаки. Відстань між генами вимірюється у відсотках кросинговеру. Один відсоток кросинговеру відповідає одній морганіді.
Кожний біологічний вид характеризується специфічним набором хромосом - каріотипом.
5. Генетика статі . 6. Механізми генетичного визначення статі .
Ще наприкінці XIX сторіччя вчені звернули увагу на відмінність за однією із пар гомологічних хромосом у хромосомних наборах самців і самок. У ди-плоїдних соматичних клітинах людини хромосоми кожної з двадцяти двох пар (нестатеві хромосоми -аутосоми) подібні між собою, а хромосоми двадцять третьої пари - різні (статеві хромосоми - гетерохромосоми). Одна з гетерохромосом - паличкоподібної форми (так звана Х-хромосома), інша -гачкоподібна (У-хромосома). Отже, каріотип жінки можна позначити як 44А + XX, а чоловіка - 44А + XУ (символом "А " позначають аутосоми).
Оскільки під час мейозу гомологічні хромосоми розходяться до різних гамет, то у однієї статі виникає лише один тип гамет (гомогаметна стать), у іншої - два, які різняться будовою статевих хромосом (гетерогаметна стать).
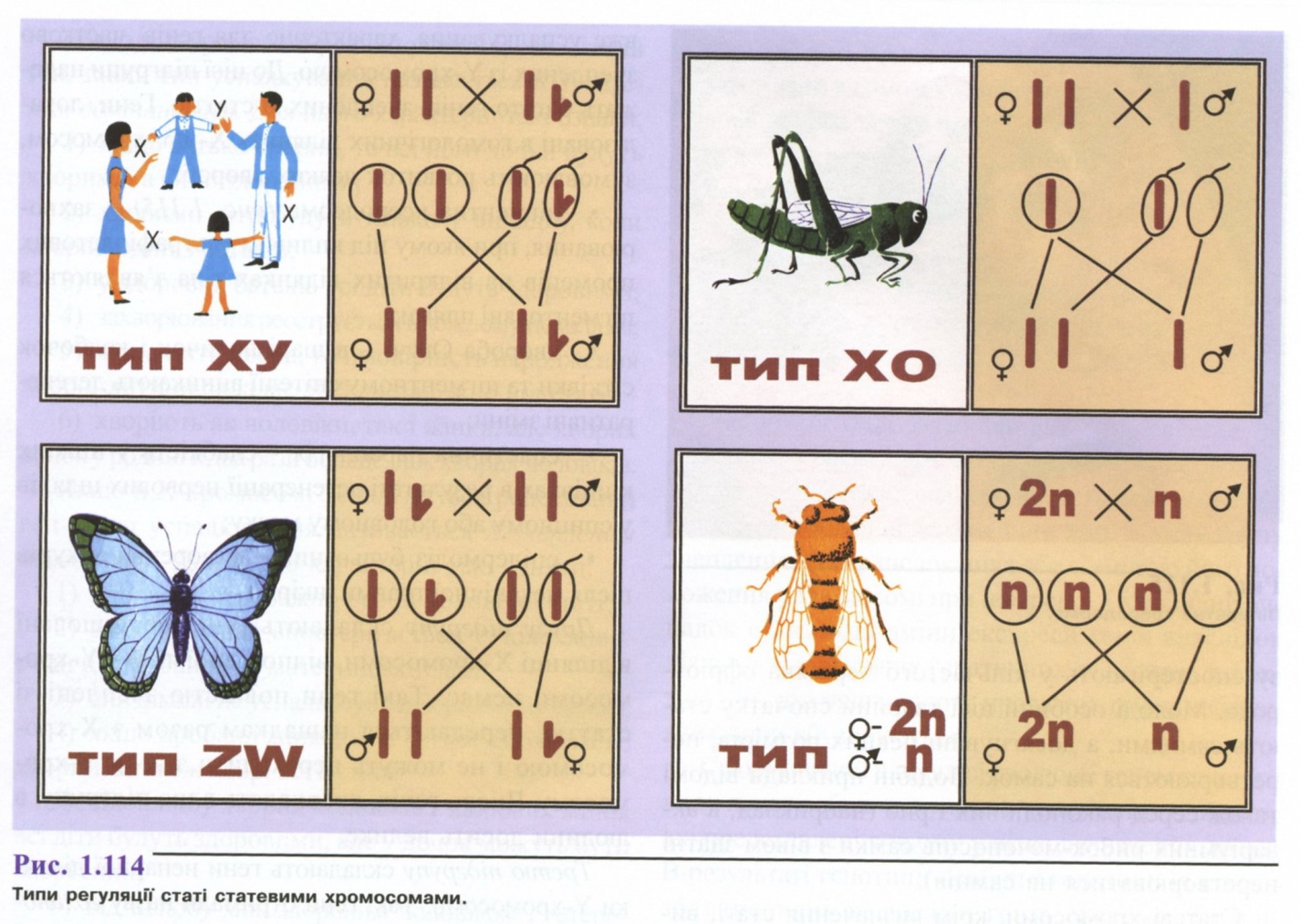
Існують чотири основних типи регуляції статі статевими хромосомами (рис. 1.114):
ХУ-тип, за якого жіноча стать має дві XX статеві хромосоми і є гомозиготною (утворює гамети одного типу - X), а чоловіча стать має чоловічу статеву хромосому - У, яка відрізняється від Х-хромосоми величиною і формою. Чоловіча стать гетерогаметна (утворює гамети двох типів - X і У). Цей тип властивий ссавцям, комахам і більшості дводольних покритонасінних рослин.
ХО-тип - жіноча стать має дві Х-хромосоми, а чоловіча тільки одну Х-хромосому. ХО-тип зустрічається серед комах і ссавців.
ZW-тип. Жіноча стать має одну жіночу статеву хромосому W і другу відмінну від неї за формою і величиною статеву хромосому Z. Тут жіноча стать є гетерогаметною, а чоловіча стать має дві однакових статевих хромосоми Z і є гомологічною.
ZW-тип притаманний деяким рибам, метеликам, птахам і дуже рідко зустрічається серед рослин.
4. Z0-тип - жіноча стать має тільки одну Z-хромосому і гетерогаметна, а чоловіча - дві Z-хромосоми і гомогаметна. Цей тип відомий тільки в одного з видів ящірки.
Статеві хромосоми, крім визначення статі, виконують і інші функції, оскільки містять гени, які впливають на різні системи органів. Успадкування ознак і властивостей, зумовлених генами, локалізованими в статевих хромосомах, називається успадкуванням, зчепленим зі статтю. Це явище було відкрито Т. Морганом. Характер зчепленого зі статтю успадкування істотно залежить від того, в яких статевих хромосомах і в яких їх ділянках розміщені ті чи інші гени.
У статевих хромосомах міститься одна з найбільших груп зчеплення. До її складу входить близько 90 генів. Ці гени поділяються на три підгрупи залежно від того, в яких ділянках X- і У-хромосом вони розташовані.
Самостійна робота № 5. Розв’язування задач на зчеплене успадкування
та взаємодію генів.
Розв’язування задач на зчеплене успадкування та взаємодію генів.
Завдання № 1. Зчеплене успадкування, кросинговер.
Допоміжна інформація : Гени , локалізовані в одній хромосомі, називають групою зчепленя, а процес обміну генами, або ділянками гомологічних хромосом називають кросинговером або перехрестом хромосом. В результаті кросинговеру з’являються рекомбіновані гамети. Але їх буде не більше ніж некомбінованих. Дигетерозіготна особина А В дає некомбіновани гамет
а в
АВ та ав більше 50% , а рекомбінованих Ав та аВ меньше 50%. При гомозіготності
особини А В або а в перехреста хромосом виявити неможливо, томущо обмін
А В а в
між ідентичними ділянками не дає нових комбінацій. Кількість кросинговеру вимірюеться : кількість кросоверів до загальної кількості особин в поколінні в % або в морганідах. Чим далі розміщені гени на хромосомі тим більше перехресників ( кросоверів).
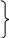
АВ Ав
80% перехресники : 20%
ав аВ
а + в
% кросинговеру можна розрахувати по формулі : х = ------------- 100%
n
а - кількість кросоверів одного класу
в - кількість кросоверів другого класу
n - загальна кількість особин.
Задача № 1. У людини вроджене захворювання очей - катаракта (С) та одна з форм анемії – еліптоцитоз (форма ерітроцита) -(Е) успадковуються зчеплено як аутосомні домінантні ознаки. Написати можливі фенотипи та генотипи дітей від браку чоловіка хворого на еліптоцитоз та катаракту , який є гетерозиготним по цим ознакам з жінкою яка здорова.
Розв’язування задачі:
Вводимо позначки: С – катаракта;
с - норма;
Е – елиптоцитоз;
е - норма.
Чоловік СЕ ; жінка се
се се
_______________________________
Р ♂ СЕ х ♀ се
се се
 гамети
:♂ СЕ; се
; Се; сЕ;
♀ се;
гамети
:♂ СЕ; се
; Се; сЕ;
♀ се;
основні перехрещені ↓
F : СЕ се Се сЕ
 се
се
се
се
се
се
се
се
батьківські форми (> 50%) нові форми(<50%)
Відповідь : Можливо проявлення ознак батьківських форм , кожної в рівній мірі та можливе появлення дітей з якою небудь однією аномалією (також в рівній мірі ).
Задача № 2. У людини ген резус-фактору зчеплений з геном який визначає форму ерітроцитів. Та знаходиться від ньго на відстані 3 морганіди. Резус-позитивність та еліптоцитоз визначаються домінантними генами. Один з супругів гетерозіготен з обох ознак. При цьому резус-позитивність успадкована від одного з батьків, а еліптоцитоз від другого. Другий супруг – резус-негативний та має нормальний ерітроцит. Визначте у відсотках вірогідні генотипи та фенотипи дітей в цій родині .
Розв’язування задачі :
Вводимо позначки : Д – резус-позитивний
д- резус-негативний
А – еліптоцитоз
а – норма .
генотип чоловіка : Да; тому що (А) успадкував від одного з батьків, (Д) від другого.
дА;
генотип жінки : да;
да ; ____________________________________
Відомо, що у чоловіка утворюеться 3% гамет – перехресників, тобто 1,5% - ДА та
1,5% - да , другі без перехреста : 97% ( 48,5% - Да та 48,5% дА ). У жінки формуеться гамети тільки одного гатунку (сорту) - да .
Р ♂ Да х ♀ да
дА да
гамети : 48,5 % Да ; 48,5% дА тільки - да 100%
1,5 % ДА; 1,5% да
F 48,5% Да 48,5% дА 1,5% ДА 1,5% да
да да да да
Відповідь:
За фенотипами: 48, 5% резус-позитивний з нормальними ерітроцитами
48,5% резус-негативний з еліптоцитозом
1,5% резус-позитивні з еліптоцитозом
1,5% резус-негативні з нормальними ерітроцитами
Розв’яжіть задачу:
Задача № 3. У людини ген який визначає синдром дефекта нігтів і наколінка (коліна чашечка) та ген визначаючий групу крові (АВО) зчеплені між собою та знаходяться на відстані 10 морганід. Синдром дефекту нігтів та наколінка успадковуеться за домінантним типом. Один з супругів гетерозіготен за синдромом та має 4 групу крові. Відомо, що цей супруг успадкував від свого батька ген 2 групи крові та синдром. Другий супруг гомозиготен за рецесивним геном аномалії та має 1 групу крові. Визначте верогідні фенотипи дітей від цьго браку.
Завдання № 2. Взаємодія алельних генів.
А). Основна форма взаємодії - повне домінування, яке вперше описано Г. Менделем. Суть його полягає в тому, що в гетерозиготного організму прояв одного з алелів домінує над проявом іншого. За повного домінування розщеплення за генотипом 1:2:1 не співпадає з розщепленням за фенотипом - 3:1. У медичній практиці з двох тисяч моно-генних спадкових хвороб у майже половини має місце домінування прояву патологічних генів над нормальними. У гетерозигот патологічний алель проявляється у більшості випадків ознаками захворювання (домінантний фенотип).
( цей тип задач розглядався в СПРС № 4 )
Б). Неповне домінування - така форма взаємодії, коли у гетерозиготного організму (Аа) домінантний ген (А) не повністю пригнічує рецесивний ген (а), внаслідок чого проявляється проміжна між батьківськими ознака. Тут розщеплення за генотипом і фенотипом співпадає і складає 1:2:1.
Розв’яжіть задачу:
Задача № 4.Еліптоцитоз (серповидноклітинна анемія) успадковується як неповністю домінантна аутосомна ознака. Гомозіготні ( за еліптоцитозом) особини помирають не доживши до статевого дозрівання, гетерозіготні – життєздатні та не хворіють малярією, особини з нормальними ерітроцитами хворіють на марярію.
Яка верогідність народження дітей стійких до малярії, в родині де один з батьків гетерозіготний . а другий має нормальні ерітроцити?
Яка верогідність народження дітей , які будуть хворіти малярією, в родині , де обидва батьки стійкі до марярії?
( ця задача розв’язується по типу задач з СПРС № 4)
В). При кодомінуванні в гетерозиготних організмів кожний з алельних генів викликає формування залежного від нього продукту, тобто виявляються продукти обох алелів. Класичним прикладом такого прояву є система груп крові, зокрема система АВО, коли ерітроцити людини несуть на поверхні антигени, що контролюються обома алелями. Така форма прояву носить назву кодомінування. Так, ген JA визначає утворення в еритроцитах антигену А, тоді як ген JB зумовлює утворення антигену В. Люди гетерозиготні за цими генами JAJB, містять в еритроцитах як антиген А, так і антиген В.
Задача № 5. У матері 1 група крові, у батька 4 група. Чи можливо успадкування дітьми груп крові одного з батьків?
Розв’язування:
Р ♀ JО JО х ♂ JA JВ
Гамети JО JA ; JВ
F JA JО ; JВ JО;
2 група 3 група
Відповідь : У дітей неможливо успадкування груп крові одного з батьків.
Розв’яжіть задачі:
Задача № 6. В батьківстві підозрюють 2-х чоловіків з 4-ю та 1-ю групами крові. Хто є батьком дитини, якщо у неї 2 група крові, а у матері 1 ?
Задача №7. В пологовому будинку переплутали 2-х хлопчиків . Батьки одного з них мають 1 та 2 групу крові, батьки другого 2 та 4. Дослідженнями виявили, діти мають один 1 , другий 4 групу. Визначте , де чья дитина.
Г). Наддомінування - коли домінантний ген у гетерозиготному стані виявляється сильніше, ніж у гомозиготному. Так, у дрозофіли при генотипі АА -нормальна тривалість життя; Аа - подовжена тривалість життя; аа - летальний наслідок.
Розв’яжіть задачу:
Задача № 8. Ген - М відповідає за формування нормальної м’язової маси, в гетерозіготному стані формується атлетичний тип статури, ген – м провокує недорозвиток деяких м’язів. Яких очикувати дітей у жінки з атлетичним типуом статури, супруг якої має недорозвиток деяких м’язів?
( ця задача розв’язується по типу задач з СПРС № 4)
Д). Множинні алелі позначаються однією літерою з різними індексами, наприклад Ja, Jb, Jc,(групи крові) або А1, А2, А3, А4... Алельні гени локалізуються в однакових ділянках гомологічних хромосом. Оскільки в каріотипі завжди присутні по дві гомологічних хромосоми, то i при множинних алелях кожний організм може мати одночасно лише по два однакових або різних алелі. У статеву клітину (разом із розходженням гомологічних хромосом) попадає тільки по одному із них.
Для множинних алелів характерний вплив всіх алелів на одну і ту ж ознаку. Відмінність між ними полягає лише в ступені розвитку ознаки.
Одним з прикладів прояву множинних алелів у людини є успадкування групи крові по системі АВО.
( дивись задачі по кодомінування , на групи крові , пункт В, завдання 2).
Завдання № 3. Взаємодія неалельних генів.
А). Комплементарність -такий тип взаємодії неалельних генів, коли один домінантний ген доповнює дію іншого неалельного домінантного гена, і вони разом визначають нову ознаку, яка відсутня у батьків. Причому відповідна ознака розвивається тільки в присутності обох неалельних генів.
Задача № 9. Гени, які визначають високу та помірну форму короткозорісті – домінантні, їх нормальні алелі – рецесивні. Жінка з помірною короткозорістю одружилаласьз чоловіком у якого висока висока форма короткозорісті, при цьому слід враховувати, що наявність гена, визначаючого помірну форму короткозорісті посилює дію гена який контролює високу форму короткозорісті. При наявності 2-х домінантних генів з різних алельних пар проявляється комплементарна їх дія. Як будуть проявлятись ці гени у дітей від цього подружжя?
Розв’язування задачі:
Вводимо позначки: К – помірна короткозорість;
к - норма;
Р - висока короткозорість;
р - норма.
_________________________________________
Р ♀ Ккрр х ♂ ккРр
Гамети( тільки такі різновиди) Кр; кр; кР; кр;
↓
F КкРр ; Ккрр; ккРр ; ккрр;
25% 25% 25% 25%
Висока помірна висока нормальний
зір.
Відповідь: Можливі генотипи батьків : КкРр ; Ккрр; ккРр ; ккрр. З фенотипів дуже високий відсоток хворих дітей 75%, 50% будуть мати високу ступінь короткозорості.
Розв’яжіть задачу:
Задача № 10. у людей одна з форм спадкової глухоти визначається двома рецесивними незчепленими алелями : д та е . Для нормального слуху необхідна наявність 2 домінантних алелів : Д та Е . Визначте генотипи в родині, де обидва батьки глухі. А їх можуть мати нормальних слух.
Б). Епістаз - це така взаємодія неалеьних генів, за якої один ген пригнічує дію іншого неалельного гена. Пригнічення можуть викликати як домінантні, так і рецесивні гени (А>В, а>В, В>А, b>А), і залежно від цього розрізняють епістаз домінантний і рецесивний. Пригнічуючий ген отримав назву інгібіторa, або супресора. Гени-інгібітори в основному не детермінують розвиток певної ознаки, а лише пригнічують дію іншого гена.
Задача №11. Деякі кури мають біле пір’я , другі забарвлені. Колір пір’я визначається 2 генами (не зчепленими та неалельними). Ген С відповідає за забарвлення, а його рецесивна алель – с за відсутність забарвлення (біле пір’я). Ген М пригнічує ген С, а ген – м не пригнічує алель С. При схрещуванні білих кур з кольоровими всі курчата були білі. Коли ці курчата підросли , провели схрещювання між ними, після чого з’явились кольорові курчата. Визначте розщеплення по фенотипу в F2 ?
Розв’язування задачі:
Вводимо позначки: С – кольорове пір’я;
с - незабарвлене (біле);
М – пригнічування( епістаз) дії гена М ;
м - не пригнічує.
Р ССмм х ссММ
F1 СсМм х СсМм
F2 :
-
♂
♀
СМ
См
сМ
см
СМ
ССММ
біле
СС ССМм
біле
СсММ
біле
СсМм
біле
См
ССМм
біле
ССмм
кольор.
СсМм
біле
Ссмм
кольор.
сМ
СсММ
біле
СсМм
біле
ссММ
біле
ссМм
біле
см
СсМм
біле
Ссмм
кольор.
ссМм
біле
Ссмм
біле
Відповідь : Розщеплення по фенотипу 13 : 3 ! ( 13 білих та 3 кольорові).
Розв’яжіть задачу :
Задача № 12. У коней масть може бути чорною, рудою або сірою. Локалізований в автосомі домінантний ген В відповідає за формування чорної масті, рецесивний ген – в за формування рудої масті. Обидва алелі можуть пригнічуватись домінантним алелем – С, унаслідок чого формується сіра масть. Рецесивний алель - с не перешкоджає прояву алелей В та в . При схрещуванні двох особин, що мають генотипи ВвСс . отримали коня чорної масті. Які ймовірні генотипи мін може мати? Яка ймовірність, що після другого схрещування тих самих батьківських особин народиться знову кінь чорної масті ?
В). Більшість кількісних ознак організмів визначається декількома неалельними генами (полігенами). Взаємодія таких генів у процесі формування ознаки називається полімерною .
У цьому випадку два або більше домінантних алелі однаковою мірою впливають на розвиток однієї і тієї ж ознаки. Тому полімерні гени прийнято позначати однією літерою латинського алфавіту з цифровим індексом, наприклад: А1А1 і а1a1, А2А2, а2а2; А3А3 і a3a3.
Задача № 13. У людини різниця кольору шкіри обумовлена як правило 2 парами домінантних генів : А1А1 А2А2 - чорний колір, та а1а1 а2а2 - білий колір шкіри. Які –небудь 3 домінантні гени дають темний колір шкіри, які – небудь 2 – смуглявий колір, один домінантний ген – світлу шкіру. А). Визначте генотипи батьків, коли обидва смугляві та мають одну дитину з чорним кольором шкіри, а другу дитину з світлою шкірою?
Розв’язування задачі:
Розв’язується методом підбору по генотипам дітей:
Генотипи батьків: Р А1а1 А2а2 х А1а1 А2а2
Генотипи дітей: F1 а1а1 а2а2 ; А1А1 А2А2
світла смугла
Відповідь : генотипи батьків А1а1 А2а2 .
Розв’яжіть задачу :
Задача № 14: Умови задачі такіж як у задачі №13, тільки . Б). Визначте генотипи смуглявих батьків які мають смуглявих дітей ?
Г). Плейотропія . Плейотропна дія генів -це залежність кількох ознак від одного гена, тобто множинна дія одного гена. У дрозофіли ген білого кольору очей воднораз впливає на колір тіла, довжину крил, будову статевого апарату, знижує плодючість, зменшує тривалість життя. У людини відома спадкова хвороба - арахнодактилія ("павучі пальці" -дуже тонкі і довгі пальці), або хвороба Марфана.
Задача № 15. Хвороба Марфана успадковується домінантним плейотропним геном. Переважне володіння лівою рукою - рецесивним аутосомним геном. Визначити верогідність прояву у дітей цих ознак, якщо батьки гетерозиготні за цими парами генів?
Розв’язування задачі:
Вводимо позначки: А – алель, яка визначає хворобу Марфана;
а - алель норми;
Е - алель, яка визначає володіння переважно правою рукою;
е - алель, яка визначає володіння переважно лівою рукою.
______________________________________________
Р ♂ Аа Ее х ♀ АаЕе
Розщеплення проходить з класичного ІІІ Закона Г. Менделя , за такими генотипами: 9 А_ Е_ - правші з синдромом Марфана ;
3 А_ ее - ліворукі та з синдромом;
3 аа Е_ - правші та зророві;
1 аа ее - ліворукі та здорові.
Відповідь : Вірогідність народження : правші з синдромом Марфана – 56,25%;
ліворукі та з синдромом - 18,75 %;
правші та зророві - 18,75 %;
ліворукі та здорові - 6,25%.
Розв’яжіть задачу.
Задача № 16: Синдром Ван дер Хеве детермінується домінантним плейотропним аутосомним геном, який проявляється в блакитному кольору склери ока, ламкістью кісток та глухотою. Один з супругів має блакитну склеру, другі ознаки у нього в нормі, другий супруг ,у якого в родині ніколи не було хворих за синдромом Ван дер Хеве, не має аномальних ознак.
Визначити вірогідність народження дітей з ознакою ламкостю кісток, якщо відомо що по лінії першого супруга ознаками синдрома володів лише один з його батьків?
(моногібридне схрещування, неповне домінування).
Самостійна робота № 6. Генетичні карти. Методи картування хромосом людини. Сучасний стан
дослідження генома людини. Генетична небезпека забруднення середовища.
Поняття про антимутаген та комутаген. Генна інженерія. Біотехнології.
Поняття про генну терапію.
Генна інженерія - галузь молекулярної біології і генетики, завдання якої - конструювання генетичних структур за заздалегідь наміченим планом, створення організмів із новою генетичною програмою. Виникнення генної інженерії стало можливим завдяки синтезу ідей і методів молекулярної біології, генетики, біохімії і мікробіології. Основні принципи генної інженерії були розроблені в 60-70-х роках XX сторіччя. Вони включали три основних етапи: а) отримання генетичного матеріалу (штучний синтез або виділення природних генів); б) включення цих генів у генетичну структуру, яка реплікується автономно (векторну молекулу ДНК), тобто створення рекомбінантної молекули ДНК; в) введення векторної молекули (з включеним у неї геном) у клітину-реципієнта, де вона вмонтовується в хромосомний апарат).
Експериментальне перенесення генів в інший геном називається трансгенезом. Він ґрунтується на технології рекомбінантної ДНК. В основі генної інженерії лежать різні методи маніпуляцій із молекулами ДНК.
Отримання генетичного матеріалу. V сучасній генетиці використовуються два способи синтезу генів поза організмом - хімічний і ферментативний. Для хімічного синтезу необхідно мати повністю розшифровану послідовність нуклеотидів ДНК. Вперше штучний ген синтезував індійський вчений Г. Корана (1970) . Це був ген аланіновоїтРНК дріжджів, який складався з 77 нуклеотидів.
Іншим прикладом хімічного синтезу є синтез гена, який кодує фермент розщеплення лактози. Синтезований у пробірці ген був вмонтований у плазміду і введений у бактерію; кишкова паличка набула здатності засвоювати лактозу.
Ген, отриманий внаслідок ферментативного синтезу, може функціонувати в бактеріальній клітині. На ньому синтезується iРНК, а потім білок. Під керівництвом
В. Енгельгардта був отриманий ген, який визначає синтез ферменту галактозидази. Цей ген вводили у фаг, при розмноженні якого в клітині одержали безліч копій, що забезпечило синтез великої кількості ферменту. Це має не тільки теоретичне, але й практичне значення, тому що галактозидаза застосовується в харчовій промисловості.
Синтезовано гени глобіну людини, кроля, голуба, деякі гени мітохондрій печінки пацюків і багато інших.
Так були синтезовані два гени, відповідальні за синтез ланцюгів інсуліну, їх уводили в геном кишкової палички, яка почала продукувати інсулін. Важливим досягненням генної інженерії є синтез гена соматостатину, який може функціонувати у мікробній клітині. Таким же методом під керівництвом Ю. О. Овчиннікова і М. П. Дубініна здійснений синтез генів, які кодують нейрогормони людини (лейцин-енкефалін і брадикінін).
Ферментативний синтез генів має велике значення, тому що принципово можливо проводити штучний синтез будь-яких індивідуальних генів шляхом транскрибування їх із відповідних матричних РНК. Основною перешкодою є синтез не структурних, а регуляторних частин генів, необхідних для їх нормальної роботи. Це здебільшого обмежує використання штучно синтезованих генів.
Клонування генів - це процес, що включає виділення й ампліфікацію (дублювання великої кількості) окремих генів у реципієнтних про- й еукаріотичних клітинах. Ці клітини, які містять потрібний нам ген, можна використовувати для одержання: а) великої кількості білка, що кодується даним геном, або б) великої кількості самого гена у високоочищеному вигляді.
Рекомбінантна ДНК-технологія має як наукове значення (дозволяє виділити окремий ген складного організму і вивчити його функцію на молекулярному рівні), так і практичне застосування. За допомогою рекомбінантної ДНК-технології можна вироб-
ляти різні білки для медичної практики. Такі ліки більш безпечні, ніж аналогічні білки, отримані безпосередньо з організмів. Першим таким рекомбі-нантним препаратом став інсулін.
Інший важливий напрямок біотехнології- виробництво вакцин. Такі вакцини не можуть викликати хвороб, тому що виготовляються з одного із поверхневих білків. Ген такого білка використовується для біореконструкції бактерії. Так створена вакцина проти гепатиту В. Успішно ведеться робота над вакцинами для гепатитів А, С, хламідіозів, герпесу й інших захворювань.
Ведуться роботи зі створення біоінженерних рослин, що могли б мати наступні властивості: 1) високу пристосованість до умов зовнішнього середовища; 2) містити більшу кількість необхідних для людини поживних речовин; 3) тривалий час зберігатися без псування.
Розробляються трансгенні рослини, здатні продукувати в інтересах людини хімічні речовини й ліки. Реконструйовано картоплю для продукції альбуміну людини. Передбачається, що в майбутньому рослини зможуть утворювати у своїх насіннях такі білки, як гормони людини.
Швидкими темпами розвивається біоінженерія тварин. Яйцеклітину поміщають у спеціальну мішалку разом з чужорідною ДНК і дрібними силікон-карбідними голками. Голки роблять множинні отвори в оболонці, крізь які ДНК попадає в клітину. За допомогою цієї технології бичачий гормон росту був введений у яйцеклітини багатьох видів тварин. Завдяки цій технології отримані великі риби, корови, свині, кролики, вівці. Трансгенні тварини створені для виробництва продуктів медичного значення.
Геном миші. Розшифровка геному людини в лютому 2001 року не дала відповіді на всі питання антропогенетики. Незважаючи на секвенування геному і визначення приблизної кількості генів, що кодують білки, функція більшості з них залишається невідомою. На даний час не ідентифіковано всі гени, що відповідають за розвиток спадкових хвороб і хвороб зі спадковою схильністю.
Розшифровка геному миші, яку використовували в лабораторних дослідженнях з 1900 р., у грудні 2002 року дала нові можливості для вивчення геному людини.
Кількість генів миші складає 27000-30500, що приблизно відповідає кількості генів людини. Близько 99 % цих генів мають нуклеотидну послідовність, властиву людському геному і 96 % із них знаходяться в "синтенних" ділянках хромосом миші і людини.
На даний час можливе виведення трансгенних мишей з точним "вимиканням" (делецією) генів, проводити аналіз змін фенотипу, що виникають внаслідок цього, і згодом шукати подібні нуклеотидні послідовності в геномі людини. Це прискорить ідентифікацію генів, що відповідають за розвиток певних нормальних і патологічних ознак у людини.
Аналіз нуклеотидної послідовності миші дозволить точно "націлювати" гени людини у схожі нуклеотидні послідовності миші і створювати "людські" ознаки у лабораторних трансгенних мишах. Наприклад, включені в геном миші людські гени білків ци-тохромів, що беруть участь у метаболізмі ліків, дозволять точно моделювати дію ліків.
Сучасний стан дослідження генома людини.
Міжнародна програма "Геном людини" була сформульована на початку 90-х років XX ст. Внаслідок копіткої роботи до початку 2003 р. геном людини повністю секвенований, тобто повністю прочитана послідовність трьох мільярдів пар основ, з яких побудована ДНК всіх 23 пар хромосом люди-
ни. Генетична довжина геному людини складає 3000 сМ (сантиморганіда, генетична відстань, яка дорівнює 1 % кросинговеру). Розв'язання програми "Геном людини" сприяло створенню генетичної карти, відтворенню цитогенетичної карти геному .
Генетична карта передбачає встановлення послідовності розміщення генетичних маркерів з відстанню не більше 1 см вздовж усіх хромосом. Така генетична карта дозволяє картувати будь-який ген, встановити відносну відстань між локусами.
Винятковим успіхом програми "Геном людини" було створення інтегральних (фізичних) карт геному.
Картовано близько 40000 кодуючих послідовностей. Загальне число генів, очевидно, складатиме 30500-40000.
На сьогодні весь геном людини клонований у вигляді великих фрагментів, які перекривають один одного. Розташування кожного з цих фрагментів на хромосомі визначено з високою точністю.
Термін клонування означає, що ген картований, виділений, вивчена його структура, знайдена мутація, яка викликає те чи інше захворювання.
Розкриття геному людини сприятиме розвитку нових напрямків медицини, вивченню природи спадкових і злоякісних хвороб, розробці генної і клітинної терапії.
Генетичні карти.
Генетична карта - це відрізок прямої, на якій позначено порядок розташування генів і вказано відстань між ними у відсотках кросинговеру. Вона будується на основі результатів аналізуючого схрещування.
Картування проводиться, щоб дізнатися, в якій парі хромосом і на якій генетичній відстані (коефіцієнт рекомбінації), або в якій частині хромосоми розташовані гени.
П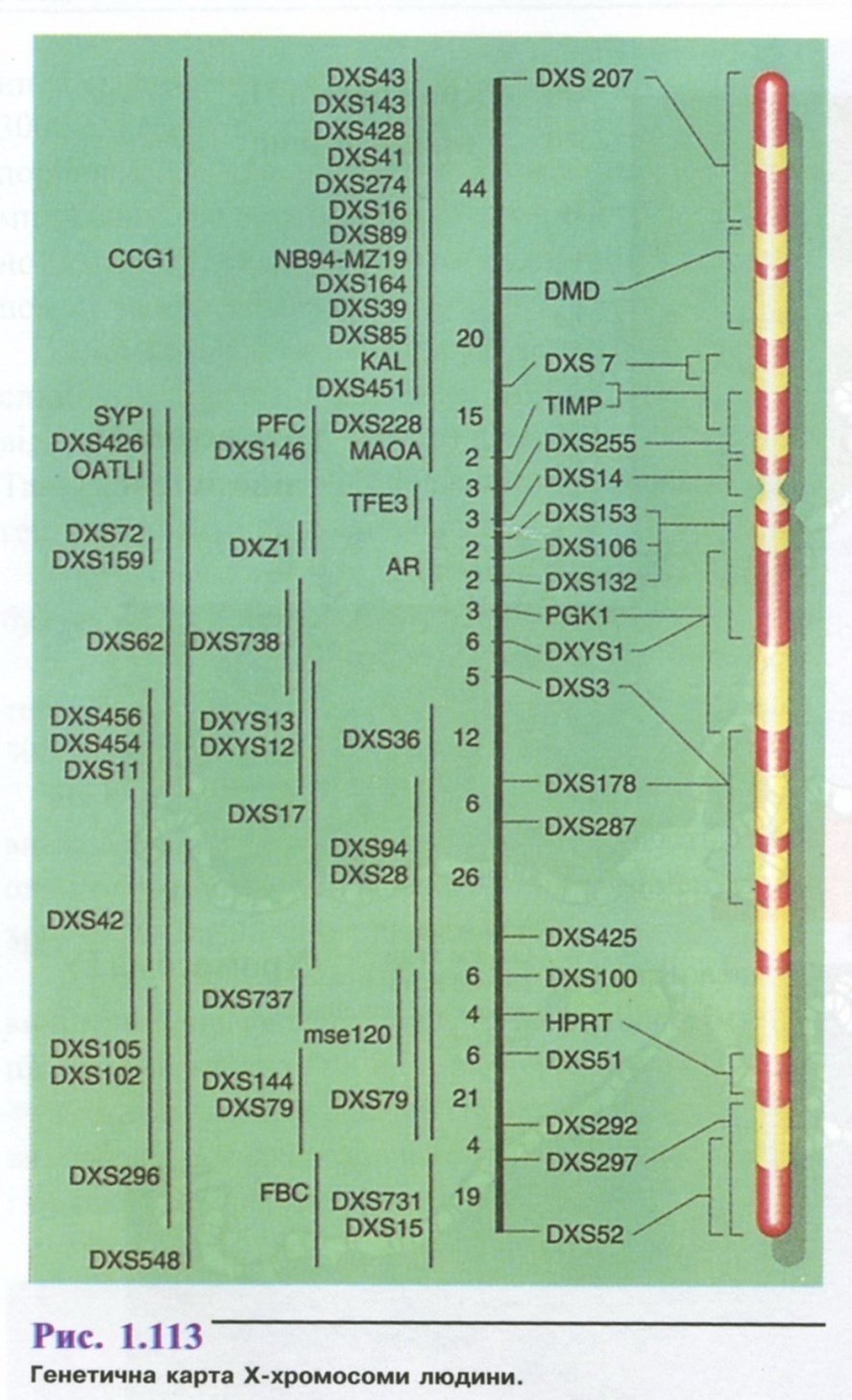 обудовано
генетичні карти для багатьох об'єктів.
Серед покритонасінних рослин краще за
все вивчено генетичні карти кукурудзи,
ячменю, рису, томатів та ін. У тварин
такі карти складено для дрозофіли,
а серед ссавців - для миші. Доведено,
що такий розподіл генів у хромосомах є
загальнобіологічною закономірністю.
обудовано
генетичні карти для багатьох об'єктів.
Серед покритонасінних рослин краще за
все вивчено генетичні карти кукурудзи,
ячменю, рису, томатів та ін. У тварин
такі карти складено для дрозофіли,
а серед ссавців - для миші. Доведено,
що такий розподіл генів у хромосомах є
загальнобіологічною закономірністю.
У людини аналіз зчеплення генів класичними методами, розробленими на дрозофілі, неможливий внаслідок неможливості експериментальних шлюбів. В останні роки розроблено комплекс методів картування генів на хромосомах людини. Так, для вивчення груп зчеплення і складання карт хромосом використовують понад 15 методів.
Картування хромосом у людини проводиться шляхом аналізу родоводів та генетичного аналізу гібридних соматичних клітин.
Тривалий час були відомі тільки три аутосомні групи зчеплення і Х-хромосомна. Новий період розпочався з 1968р.коли вдалося встановити локалізацію гена групи крові Даффі у хромосомі 1. У людини відомі всі 24 групи зчеплення.
З загальної кількості генів геному людини 16000 тепер картовані на скелетній карті геномад, складеній за допомогою 1000 високополі-мерних ДНК-маркерів.
Виявлення груп зчеплення у хромосомах дає можливість побудови хромосомних карт, коли на хромосомі розміщення генів позначають у вигляді цяточок. Найбільш повно побудована карта Х-хро-мосоми. Локуси, зчеплені з Х-хромосомою (а їх понад 200), віднесені до цієї хромосоми спираючись на аналіз родоводів (багато з них підтверджені методами гібридизації соматичних клітин).
Методи побудови карт хромосом ґрунтуються на передбаченні ймовірності кросинговеру по всій довжині хромосоми і випадкових обмінах між різними хроматидами.
Для картування хромосом застосовують гібридизацію соматичних клітин людини з клітинами різних тварин, переважно з клітинами миші. При культивуванні таких гібридних клітин на живильному середовищі відбувається постійна втрата хромосом людини. Коли в гібридній клітині залишається тільки одна хромосома від каріотипу людини, її можна ідентифікувати шляхом диференціального забарвлення. Визначивши потім вміст певного ферменту в даній клітині, можна стверджувати, що локус гена до цього ферменту розташований в цій хромосомі.
Незважаючи на певні труднощі при побудові генетичних карт і на їх відносність, детальна розробка карт - один з провідних методів аналізу поведінки і тонкої будови хромосом.
На рис. 1.113 зображено генетичну карту X-хромосоми. Символами позначено картовані гени з указанням ділянок їх локалізації.
Складання точних карт хромосом є необхідною передумовою подальшого прогресу теоретичної генетики людини. Це має виняткове значення для виявлення та диференціальної діагностики спадкових хвороб плода, для ранньої діагностики цих хвороб, встановлення носіїв генетичних зрушень, які перебувають у безсимптомній фазі, для визначення медико-генетичного прогнозу.
Генетична небезпека забруднення середовища. Моніторинг генетичний - заходи, спрямовані на відслідковування виникнення і поширення спадкової патології. Це комплексне обстеження певних груп населення, спрямоване на реєстрацію подій генетичного характеру, виявлення нових мутацій (анеуплоїдії і тяжких форм домінантних мутацій), успадкованих від попередніх поколінь (генних і хромосомних захворювань), виявлення тимчасових тенденцій або відмінностей у просторовому розподілі хвороб із спадковою схильністю, пошук можливих причин зв'язку між збільшенням частот цих подій з факторами довкілля.
Можна проводити моніторинг великих популяцій для пошуку нових мутацій. У процесі такого обстеження застосовують скринінг на домінантні мутації, які дають специфічні фенотипи. Виконують скринінг на геномні і хромосомні мутації. Для цього необхідно здійснювати скринінг популяцій цілих країн. Проведення таких заходів забезпечує отримання інформації про мутаційні частоти та їх зміну в залежності від умов середовища та генетичної компоненти.
Мутаційний процес - явище, властиве всій живій природі. Людина не є винятком. Цей процес проходить постійно й інтенсивно на генному, хромосомному і геномному рівнях. Близько 20 % всіх спадкових хвороб у кожному з поколінь після народження - хвороби, зумовлені новими мутаціями (М. П. Бочков, 2001), тобто внесок мутацій у спадкову патологію вагомий. Відомо, що індукований мутагенез, зокрема вплив радіації і хімічних мутагенів завдає тільки шкоди, викликають спадкові дефекти.
До заходів, спрямованих на запобігання розвитку індукованих мутацій, належить тестування на мутагенність фармацевтичних препаратів, які вже використовуються або плануються до впровадження в клініку. Існують мікробні тест-системи з використанням мікросом печінки ссавців.
Дослідженню на мутагенність підлягають пестициди, які застосовуються або плануються до впровадження в сільське господарство.
Відомо понад 500 сполук, які мають антимутагенні властивості. До них належать вітаміни С, Е, А, інтерферон, бемитил, томерзол та ін. Механізми дії цих речовин досить складні і передбачають залучення їх у ферментативнозалежний антиради-кальний ланцюг і здатність до зв'язування супероксидного аніон-радикалу кисню. Вітамінопрофілактика мутагенезу передбачає призначення вітамінних комплексів різного кількісного й якісного складу, що призводить до зниження числа пошкоджених мутагеном клітин, зменшує чутливість лімфоцитів до мутагену.
Встановлено, що рівень окиснювальних пошкоджень ДНК зменшується при споживанні низькокалорійної дієти, багатої на каротиноїди.
За умов наявності мутацій (фенілкетонурія, галактоземія та ін.) з метою зменшення негативного впливу на плід призначають внутрішньоутробне лікування. Так, вагітним з ризиком народження дитини з уродженою гіперплазією кори наднирників до 10-20 тижня вагітності призначають дексаметазон (20 мкг/кг) незалежно від стану плоду. При загрозі розвитку ацидурії вагітним призначають вітамін Вр. З виникненням аритмії у плоду вагітні отримують кардіологічні препарати. Якщо провести лікування жінок протягом 3-6 міс до запліднення та впродовж перших місяців вагітності вітамінами (С, Е, фолієва кислота), то імовірність розвитку в дитини аномалій нервової трубки істотно зменшується (М. П. Бочков, 2001).
Комутагени. Комутагени - це речовини, які підвищують ефекти середовищних мутагенів, хоча самі по собі не здатні до мутагенної дії, не мають власної мутагенної активності. Такий ефект мають сполуки природного і штучного походження, неорганічної і органічної природи. Комутагенез - зростання пошкоджувального ефекту мутагенів під дією немутагенних сполук. Так, аскорбінова кислота (вітамін С) підсилює цитогенетичні ефекти мутагенів, зокрема циклофосаміду, в культурі лімфоцитів людини: збільшує пошкодження ДНК і т. п.
Іншим комутагеном вважається кофеїн, який впливає на спонтанний та індукований мутагенез. Кофеїн збільшує індукцію метотрексатом сестринських хро-матидних обмінів, утворення мікроядер тощо.
Комутагенну дію мають деякі фармакологічні засоби. Так, верапаміл та фендилін підсилюють бластогенну дію блеоміцину, пепломіцину. Таким же ефектом володіють нестероїдні протизапальні препарати (лорноксикам, теноксикам та ін).
Присутність у середовищі комутагенів може підвищувати негативні ефекти фізичних, хімічних, біологічних, лікарських та інших мутагенів, з якими контактує людина.
Антимутагени. Відомо понад 500 сполук, в яких доведено антимутагенний вплив, - здатність їх до захисту геному від дії мутагенів. Речовини з таким спрямуванням підвищують стійкість клітин до негативного впливу мутагенів, знижують кількість пошкоджених мутагеном клітин. Так, прийом (3-каротину (25 мг), вітаміну С (100 мг) і вітаміну Е (280 мг) зменшує спонтанне пошкодження ДНК у лімфоцитах периферичної крові. Поліфенольні антиоксиданти, які містяться у зеленому чаї, значно зменшують частоту сестринських хроматидних обмінів у лімфоцитах периферичної крові людей, що курять. Прийом аскорбінової кислоти з розрахунку 1 таб/добу значно зменшує рівень хромосомних аберацій в лімфоцитах периферичної крові у робітників, які контактують з кам'яновугільними смолами та вугільним пилом. Встановлено, що прийом вітамінів А (33000 МО/добу) і С (500 мг/добу) впродовж 1 міс знижують аномально високий рівень спонтанного мутагенезу в робітників, які працюють в умовах виробництва молібдену.
Доведено, що захисний антимутагенний ефект тих чи інших речовин залежить від часу їх введення відносно мутагену, від стадії клітинного циклу. Цей ефект специфічний для різних хвороб людини і тому не має універсального характеру.
Застосування антимутагенів спрямоване на максимальну стійкість клітин людини та захист геному людини від мутагенних впливів.
Література :
Основна : с. 72 – 80, с. 168 – 169.
додаткова: Т.1с. 118 – 127,с. 143 – 145, с. 224 - 230.
Медична біологія
Лекція № 5 . Мінливість у людини, як властивість життя і
генетичне явище.
1. Властивість життя і генетичне явище.
2. Форми мінливості.
3. Фенотипна мінливість ( неспадкова ).
4. Норма реакції.
5. Генотипна мінливість ( спадкова ).
6. Комбінативна мінливість. Явище гетерозісу у людини.
7. Мутаційна мінливість. Класифікація мутацій.
Самостійна робота № 8. Поняття про мультифакторіальні хвороби.
1. Властивість життя і генетичне явище. 2. Форми мінливості.
Мінливістю називають відмінності між особинами одного виду - предками і нащадками, які виникають внаслідок змін спадкового матеріалу або впливу умов зовнішнього середовища.
Мінливість, як і спадковість, властива всій живій природі. Генетична наука розрізняє спадкову і неспадкову мінливість.
Спадкова - це здатність до зміни самого генетичного матеріалу, а неспадкова - здатність організмів реагувати на умови зовнішнього середовища, змінюватися в межах норми реакції, заданої генотипом.
Спадкова мінливість у свою чергу поділяється на комбінативну і мутаційну.
Комбінативна (рекомбінативна) мінливість виникає при гібридизації внаслідок незалежного перекомбінування генів та хромосом. Тут відбувається перекомбінація певних генетичних угруповань без якісної і кількісної зміни генетичного матеріалу.
Мутаційна мінливість виникає раптово, в результаті взаємодії організму і середовища, без схрещування. Вона зумовлена якісною зміною генетичного матеріалу, виникненням нових варіантів дискретних одиниць генетичного матеріалу, перш за все нових алелів.
Неспадкова (модифікаційна мінливість) - це фенотипна мінливість, яка виникає в процесі індивідуального розвитку організмів і не передається нащадкам.
Мінливість забезпечує різноманітність за будовою і фізіологічними функціями організмів. Вона є результатом різних процесів. Деякі з них відбуваються в спадковому матеріалі (генотипі). Інші обмежуються фенотипом.
Кожний організм у процесі розвитку і життя зберігає притаманні виду певні властивості, які контролюються спадковістю. Вона ніби закріплює рівень розвитку, досягнутий видом під дією природного добору. Кожна ознака формується в процесі онтогенезу за дії не одного, а багатьох генів і є результатом численних і складних процесів. Так, колір волосся людини або тварин залежить від складу пігменту, кількості його у волоссі, характеру розподілу по довжині волосся та ін.
У вищих тварин і людини на розвиток тієї чи іншої ознаки впливає багато генів. За участю значних ділянок молекули ДНК синтезуються відповідні іРНК, а останніх - відповідні ферменти. Отже у вищих форм процес розвитку ознаки характеризується цілим рядом специфічних взаємодій генів, які входять до генотипу організму. Тому залежно від генотипу при одному і тому ж гені ознака може зазнавати значних змін. Наприклад, масть гомозиготного за геном чорного забарвлення шерсті корови може бути суцільно чорною або різних відтінків, сірою, тигровою, кавовою або плямистою в різних ділянках тіла. Це залежить від інших генів і їх взаємодії між собою.
Проте генотип визначає тільки спрямування розвитку ознаки. У залежності від умов середовища характер розвитку ознаки може зазнавати змін. У вищих організмів і людини ембріональний розвиток відбувається за відносно стабільних умов, тому ознаки, які формуються до народження, не зазнають істотного впливу зовнішнього середовища і розви-
ваються в основному під впливом генотипу і після народження майже не змінюються. До таких ознак слід віднести: форму вух і носа, групи крові, типи гемоглобіну та ін.
Проте відомі випадки, коли, здавалося б, стійкі ознаки зазнають змін під впливом зовнішніх факторів. Так, у горностаєвих кроликів забарвлення тулуба біле, а вуха, кінчик мордочки, хвіст і кінці лапок пігментовані. Таке забарвлення зміниться, якщо на різних ділянках тіла шерсть вибрити і утримувати тварин за різної температури.
Висока температура може гальмувати прояв певних ознак у личинок дрозофіли, викликати захворювання на екзему і запалення шкіри голови у ягнят та ін.
Всі ознаки, які формуються в постембріональному періоді, зазнають істотного впливу середовища. Це можна спостерігати в монозиготних близнюків, які перебувають в різних умовах. Незважаючи на ідентичність їх генотипу, навіть маса тіла зазнає змін.
Середовище може впливати і на прояв домінантної ознаки. Отже, ступінь домінування тієї або іншої ознаки не залишається однаковим; і залежить як від спадковості, так і впливу умов середовища. Проте існують і такі ознаки, ступінь домінування яких майже не залежить від умов середовища.
Звідси виняткового значення набуває знання критичних періодів розвитку ембріона і періодів, коли зигота чутлива до дії різних чинників і легко пошкоджується, що призводить до вад розвитку.
Фенотипна мінливість ( неспадкова ).
Фенотипна (модифікаційна мінливість)-це така форма мінливості, яка не викликає змін генотипу. Одним із перших дослідників, що вивчав модифікаційну мінливість, був К. Негелі (1865).
Кожний організм розвивається за участі генотипу і під впливом зовнішнього середовища. Спадкові ознаки і властивості проявляються порізному і залежать від умов, в яких розвиток відбувається. Спадковий матеріал при модифікаційній мінливості змін не зазнає.
Зміни ознак і властивостей організму в межах норми реакції, що виникають внаслідок різних умов існування, називаються модифікаціями.
Модифікації носять адаптивний (пристосувальний) характер і мають значення для виживання організмів, виконуючи певну роль у збереженні виду. Крім корисних (пристосувальних), зміни бувають і індиферентні.
Модифікаційна мінливість-явище масове. Вона торкається не одного, а сукупності об'єктів і тому її ще називають груповою, або визначеною.
На розвиток ознаки впливають регуляторні системи організму, в першу чергу, ендокринна система. Такі ознаки в півня, як забарвлення пір'я, величина гребеня та сережок, характер співу і тембр голосу, зумовлені дією статевого гормону тестостерону. Введення півням жіночих статевих гормонів гальмує ці ознаки. Отже, внутрішнє середовище організму має значний вплив на прояв генів у формі ознак.
Не всі ознаки однаковою мірою зазнають модифікаційної мінливості. Під впливом середовища змінюються маса і розміри тіла, розумові здібності, артеріальний тиск. Проте такі ознаки, як морфологічні, -більш стійкі і менше залежать від екзогенних чинників.
Модифікаційна мінливість характеризується такими властивостями: 1) неуспадкува-нням; 2) груповим характером змін; 3) відповідністю змін щодо дії певного чинника середовища; 4) обумовленими генотипом межами змін (хоча напрямок змін однаковий, ступінь їх прояву в різних організмів неоднаковий).
Модифікаційна мінливість - це закономірне біологічне явище, яке постійно супроводжує розмноження організмів. Процес розвитку кожної ознаки або властивостей організму здійснюється на основі генотипу за різних умов середовища. Тому успадкування ознак або властивостей завжди проявляється у формі різних його модифікацій.
Модифікаційну мінливість вивчають за допомогою методів математичної статистики. Вона характеризується варіаційним рядом. Середня величина варіаційного ряду нагадує точку його рівноваги.
4. Норма реакції.
Спадкові властивості організму не можна охарактеризувати однією формою прояву. Генотип за різних умов має різний фенотип. Генотип характеризує норма реакції - діапазон адаптивної мінливості організму в середовищному градієнті. Норма реакції -це спосіб реагування генотипу на зміни довкілля. Наприклад, врожай озимої пшениці 20 ц з 1 га не дає повної уяви, хороший це сорт чи поганий. Якщо такий врожай отримано на бідному грунті або в посушливий сезон, то це хороший сорт. Якщо ж така урожайність досягнута за умов достатньої вологи і поживних речовин, то це - низькопродуктивний сорт.
Мінливість ознаки під впливом середовища не безмежна. Ступінь прояву ознаки, або межі модифікаційної мінливості, зумовлені генотипом і середовищними чинниками. Розрізняють широку і вузьку норми реакції.
Вузька норма реакції властива таким ознакам, як розміри серця або головного мозку, тоді як кількість жиру в організмі людини зазнає змін у широких межах. У вузьких межах коливається будова квітки у рослин, проте значно змінюються розміри листків.
Норма реакції генотипу виявляється в процесі модифікаційної мінливості організму. Нормою реакції визначають пристосувальні можливості сортів і ареали, які вони можуть зайняти.
5. Генотипна мінливість ( спадкова ).
Генотипна мінливість - основа різноманітності живих організмів і головна умова їх здатності до еволюційного розвитку. Механізми генотипної мінливості різноманітні, внаслідок чого розрізняють дві її основні форми: комбінативну і мутаційну.
Комбінативна мінливість - найважливіше джерело безмежно великого спадкового розмаїття, яке спостерігається в живих організмів. В основі комбінативної мінливості є статеве розмноження. Генотип нащадків являє собою поєднання генів, які були властиві батькам. Число генів у кожного організму дуже велике. При статевому розмноженні комбінації генів призводять до формуваня нового унікального генотипу і фенотипу. Комбінативна мінливість широко розповсюджена у природі. Навіть у мікроорганізмів, які розмножуються нестатевим шляхом, виникли своєрідні механізми (трансформація і трансдукція), що призводять до появи комбінативної мінливості. Все це вказує на велике значення комбінативної мінливості для еволюції та видоутворення.
Не менше значення в еволюції має і мутаційна мінливість. Саме мутаціями зумовлений поліморфізм людських популяцій: різна пігментація шкіри, волосся, колір очей, форма носа, вух, підборіддя тощо. З мутаційною мінливістю пов'язаний процес утворення нових видів, сортів і порід.
6. Комбінативна мінливість. Явище гетерозісу у людини.
Комбінативною називають мінливість, яка залежить від перекомбінування алелів у генотипах нащадків порівняно з генотипами батьків, що призводить до появи нових ознак у дітей. На відміну від перерозподілу спадкового матеріалу нерегулярного типу при хромосомних абераціях така рекомбінація в організмів, які розмножуються статевим шляхом, відбувається закономірно.
Існує три механізми комбінативної мінливості. Два з них пов'язані з мейозом. Завдяки обміну генетичним матеріалом між гомологічними хромосомами при кросинговері в профазі мейозу алельний склад хромосом статевих клітин нащадків зазнає змін порівняно з гаметами батьків. Хромосоми, успадковані від обох батьків, у метафазі мейозу, комбінуються в різних поєднаннях.
Третій механізм комбінативної мінливості пов'язаний із заплідненням. Внаслідок випадкового підбору гамет, які беруть участь у заплідненні, створюються різноманітні поєднання хромосом з різним набором генів, а відповідно, й алельного складу зигот.
Явище комбінативної мінливості пояснює причину поєднання в дітей і онуків ознак родичів по батьківській або материнській лінії.
З врахуванням міграційних процесів серед людей зростає кількість шлюбів між нареченими, які походять з віддалених регіонів. Чим більша відстань, тим більша ймовірність того, що гамети будуть відрізнятися за набором генів.
Комбінативна мінливість сприяє більшій пристосованості до мінливих умов середовища, підвищує виживання виду.
Для розуміння ефекту гетерозису в людини охарактеризуємо гетерозис як біологічне явище.
Поняття про гетерозис як прояв гібридної сили введене в науку американським генетиком Д. Г. Шеллом у 1914 році.
Гетерозис у природі властивий всім організмам: рослинам, тваринам, мікроорганізмам. Він виник одночасно з утворенням диплоїдності і статевого процесу і безпосередньо зумовлений виникненням і вдосконаленням у процесі еволюції перехресного запилення. Розрізняють репродуктивний гетерозис (кращий розвиток органів розмноження, підвищення фертильності, більший урожай плодів і насіння) соматичний гетерозис (більший розвиток вегетативних частин) і пристосувальний, або адаптивний, гетерозис (підвищення життєвої стійкості гібридів).
Для людських популяцій за останні 100-150 років характерний інтенсивний розвиток транспортних комунікацій, зросла міграція населення, що призвело до розпаду ізолятів та розширення інбридних регіонів, поширення кола потенційних партнерів при шлюбах внаслідок подолання релігійних, класових, етнічних, географічних перешкод (бар'єрів) та ін. Це призводить до змішування населення, а тому гени в поколіннях дітей набувають все більшого гетерозиготнего стану. За аддитивної дії різних гетерозиготних генотипів сукупний ефект генів у гетерозигот має бути вищим, ніж в обох гомозигот вихідних популяцій. І дійсно, за останню сотню років у багатьох країнах середній зріст населення підвищився на 5-15 см. Настання статевої зрілості змінилося з юнацького віку (16-18 років) на підлітковий вік (13-14 років).
Проте ефект гетерозису у людському суспільстві вимагає подальшого поглибленого вивчення.
7. Мутаційна мінливість. Класифікація мутацій.
Термін "мутація" запровадив Г. де Фріз (1901) для характеристики випадкових генетичних змін в рослини Ослинник энотера.
Розрізняють спонтанний та індукований мутаційні процеси. Індукований мутаційний процес - це виникнення спадкових змін під впливом направленої дії факторів зовнішнього і внутрішнього середовища. Виникнення мутацій без встановлених причин прийнято називати спонтанним мутаційним процесом.
Мутаційна мінливість зумовлена як впливом на організм чинників зовнішнього середовища, так і його фізіологічним станом. Частота виникнення мутацій залежить від:
генотипу організму;
фази онтогенезу;
статі;
стадії гаметогенезу;
мітотичного і мейотичного циклів хромосом;
хімічної будови окремих ділянок хромосом та ін. Властивості мутацій:
мутації виникають раптово, стрибкоподібно;
мутації успадковуються, тобто передаються від покоління до покоління;
мутації ненаправлені - зазнавати мутації може будь-який локус, викликаючи зміни як незначних, так і життєво важливих ознак;
одні і ті ж мутації можуть виникати повторно;
за проявом мутації можуть бути корисними і шкідливими, домінантними і рецесивними.
Класифікація мутацій:
Мутації можна об'єднати в групи - класифікувати за характером прояву, за місцем або рівнем їх виникнення.
Мутації за характером прояву - бувають домінантними і рецесивними. Більшість з них рецесивні і не проявляються у гетерозигот. Як правило, мутації шкідливі, бо порушують чітко збалансовану систему біохімічних перетворень. Домінантні мутації проявляються зразу у гомо- і гетерозиготного організму, здебільшого такі особини нежиттєздатні і гинуть на ранніх стадіях онтогенезу.
Мутації часто знижують життєстійкість або плодючість. Мутації, які різко впливають на життєздатність, частково або повністю зупиняють розвиток, називаються напівлетальними, а несумісні з життям -летальними. У людини до таких мутацій належить рецесивний ген гемофілії.
Мутації за місцем виникнення. Мутації, що виникають у соматичних тканинах, отримали назву соматичних мутацій. Соматичні клітини складають популяцію, утворену при безстатевому розмноженні (поділ клітин). Соматичні мутації обумовлюють генотипну різноманітність тканин, часто не передаються по спадковості і обмежені тим індивідуумом, у якому вони виникли. Соматичні мутації виникають у диплоїдних клітинах, тому проявляються тільки за домінантними генами або за рецесивними, але в гомозиготному стані. Чим раніше в ембріогенезі людини виникла мутація, тим більша ділянка соматичних клітин відхиляється від норми. І навпаки, чим пізніше в процесі розвитку організм зазнає мутаційного впливу, тим менша та ділянка тканини, яка утворюється із мутаційної клітини. Наприклад, забарвлення райдужної оболонки ока - білий або карий сегменти на блакитній райдужці - зумовлені соматичною мутацією. Вважають, що наслідком соматичних мутацій є ракове переродження. Злоякісний ріст викликається канцерогенами, серед яких найбільш негативні - проникаюча радіація й активні хімічні сполуки (речовини).
І хоча соматичні мутації не успадковуються, вони знижують репродуктивні можливості організму, в якому виникли.
Мутації, що виникають в гаметах або в клітинах, з яких вони утворюються, отримали назву генеративних, або термінальних мутацій. Чим раніше в статевих клітинах виникає мутація, тим більшою буде частка статевих клітин, які нестимуть нову мутацію. Верхня межа частки клітин, які будуть містити індуковану або спонтанну мутацію, складає 50 відсотків.
Існує думка, що найбільша кількість мутацій у статевих клітинах виникає в овоцитах. Оскільки сперматогонії зазнають постійного поділу, то серед них може відбуватися відбір проти мутацій, які зумовлюють шкідливий ефект, і частота мутацій знижується до часу статевої зрілості. Жінка, навпаки, народжується майже з усіма мутантними змінами, в лінії статевих клітин немає паралельного мітотичного відбору. Овоцити не тільки не зазнають мітозу, вони залишаються малоактивними впродовж десятиліть, доки не стануть яйцеклітинами. За цей період овоцити старіють, стають непропорційно чутливими до спонтанної мутації.
На статеві клітини найбільший вплив спричиняють цезій-137, стронцій-90 і вуглець-14.
Генеративні мутації при статевому розмноженні передаються наступним поколінням. Домінантні мутації з являються вже в першому поколінні, а рецесивні -тільки в другому і наступних поколіннях, з переходом у гомозиготний стан. Мутації за характером зміни спадкового матеріалу:
- Зміни, зумовлені заміною одного або декількох нуклеотидів у межах одного гена, називають генними, або точковими мутаціями. Вони обумовлюють зміни як у будові білків, так і функційної активності білкової молекули .
- Зміни структури хромосом називають хромосомними мутаціями або абераціями. Такі мутації можуть виникнути в результаті втрати частини хромосоми (делеція), подвоєння частини хромосоми (дуплікація), відриву і повороту частини хромосоми на 180° (інверсія) (рис. 1.135). Якщо зміна торкається життєво важливих ділянок гена, то така мутація призведе до смерті. Так, втрата невеличкої ділянки 21 -ї хромосоми у людини викликає тяжке захворювання крові - гострий лейкоз. В окремих випадках відірвана ділянка хромосоми може приєднатисядо негомологічної хромосоми (транслокація), що при зведе до нової комбінації генів і зміни їх взаємодії.
- Зміни каріотипу, кратні або некратні гаплоїдному числу хромосом називають геномними мутаціями. Внаслідок порушення розходження пари гомологічних хромосом під час мейозу в одній із утворених гамет міститься на одну хромосому менше, а в іншій на одну хромосому більше, ніж за нормального гаплоїдного набору. Злиття такої аномальної гамети з нормальною гаплоїдною гаметою при заплідненні призводить до утворення зиготи з меншою або більшою кількістю хромосом порівняно з диплоїдним набором, характерним для цього виду.
ю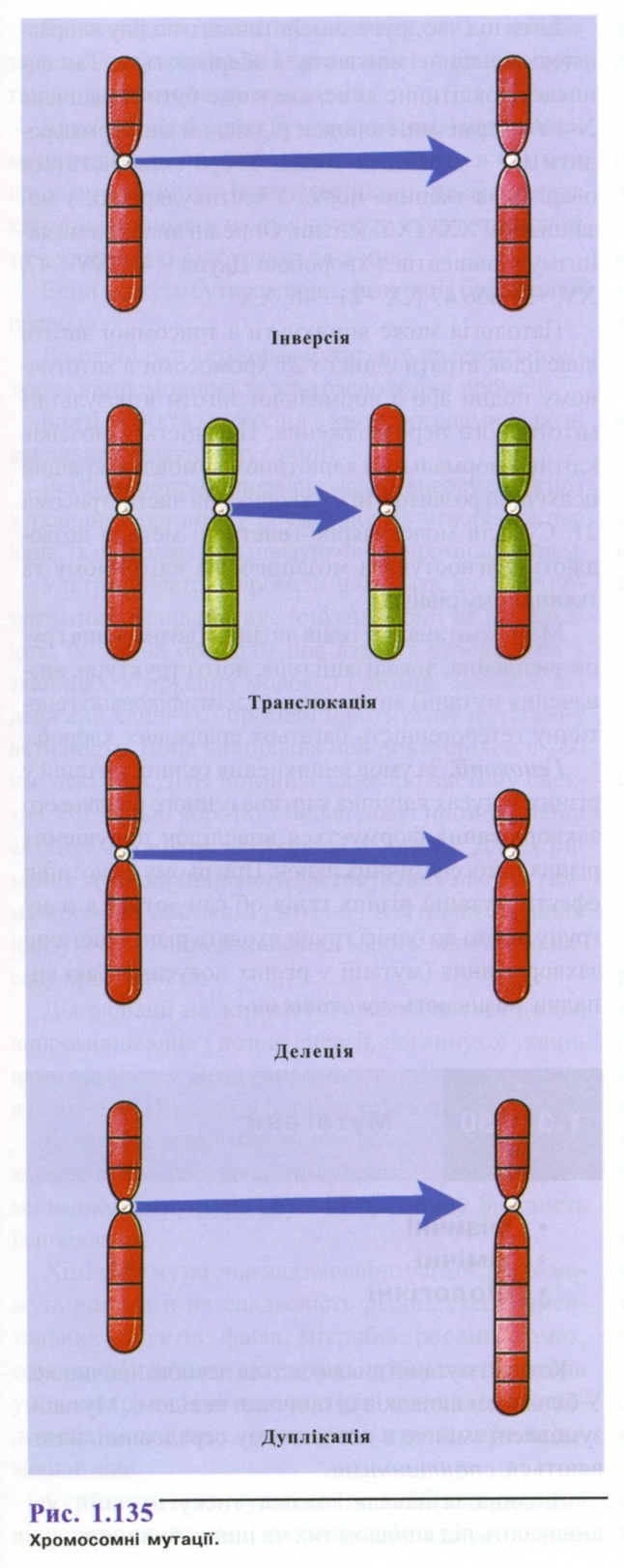 ться
по спадковості, а залишаються, доки живе
організм,
що зазнав мутаційного впливу.
ться
по спадковості, а залишаються, доки живе
організм,
що зазнав мутаційного впливу.
Геномні, хромосомні і генні аберації в соматичних клітинах є наслідком дії мутагенних чинників. У людини це етіологічні фактори спадкових хвороб. Захворювання, зумовлені геномними (зміна числа хромосом) і хромосомними (зміна структури хромосом) мутаціями, називаються хромосомними хворобами. Зміна числа хромосом визначається подвоєнням або зменшенням всього набору хромосом. Це призводить до поліплоїдії або гаплоїдії (відповідно). Лишні або видалення однієї або декількох хромосом призводить до гетероплоїдії або анеуплоїдії. Зміна структури хромосом - це перебудови або аберації. При цьому порушується збалансованість набору генів і нормальний розвиток організму. Як наслідок хромосомного дисбалансу відбувається внутрішньоутробна загибель ембріона або плоду, виникають уроджені вади розвитку. Чим більша кількість хромосомного матеріалу зазнала мутаційного ефекту, тим раніше захворювання з'явиться в онтогенезі і тим вагоміші будуть порушення фізичного і психічного розвитку особини. Характерна риса хромосомного дисбалансу - множинність вад розвитку різних органів і систем.
Хромосомні хвороби складають близько 0,5-1 % всіх спадкових хвороб людини.
Генні, або точкові мутації - це результат молекулярних змін на рівні ДНК. У людини вони викликають генні хвороби. У людини описані наступні види генних мутацій, які призводять до розвитку спадкових хвороб: нісенс, нонсенс, зміщення рамки зчитування, делеції, вставки (інсерції), порушення сплайсингу, збільшення числа (експансії) тринуклеотид-них повторів. Мутації ділянок, що транскрибуються (які визначають амінокислотну послідовність у молекулі білка, що синтезується), призводять до синтезу аномального продукту. Мутації ділянок, що транскрибуються, можуть призвести до зменшення швидкості синтезу білка.
Фенотипно генні мутації виявляються на молекулярному, клітинному, тканинному і органному рівнях. Число генних хвороб складає близько 3500-4500.
Генні мутації поділяють на односайтові і багатосайтові. Односайтові - це такі, які торкаються змін одного сайту, багатосайтові - охоплюють декілька сайтів генного локусу.
Розрізняють генні мутації прямі й обернені. Прямі мутації - це мутації, які інактивують гени дикого типу і зумовлюють появу мутантного типу. Обернені мутації, або зворотні, - зміни до висхідної форми від мутантної.
Більшість генів стійкі до мутацій, але окремі гени зазанають мутацій досить часто.
Соматичні мутації зумовлюють генотипну різноманітність тканин однієї особини і здебільшого не передаються по спадковості при статевому розмноженні. При безстатевому розмноженні, якщо організм розвивається з однієї клітини або групи клітин, в яких виникла мутація, - такі зміни можуть передаватися нащадкам. Соматичні мутації використовують у організмів, які розмножуються вегетативне. Соматичні мутації складають основу селекції культурних рослин, зокрема цитрусових.
Самостійна робота №8. Поняття про мультифакторіальні хвороби.
Мультифакторіальний принцип формування фенотипу. Значення умов середовища для експресивності й
пенетрантності генів.
У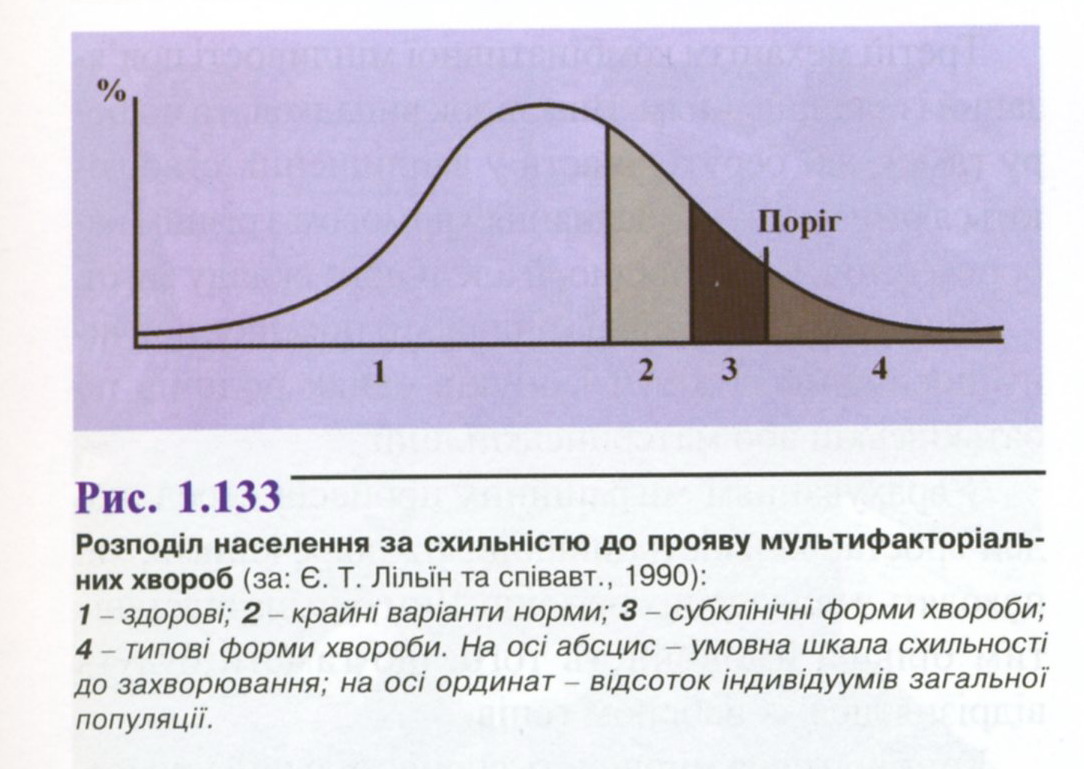 розвитку одних ознак визначальну роль
виконує генотип, а для розвитку інших
істотне значення має зовнішнє середовище.
Проте немає таких ознак, які абсолютно
зумовлювались би тільки спадковістю
або тільки середовищем. Ознаки, прояв
яких залежить від багатьох генів, більшою
мірою зазнають впливу екзогенних
чинників. Такі ознаки є мультифакторіальними
- за цих умов формування фенотипу
відбувається шляхом взаємодії спадковості
і середовища, генетичних і негенетичних
чинників. У людини такими ознаками є
зріст, розумові здібності, артеріальний
тиск, поведінкові реакції та ін. Дія
будь-якого гена залежить від інших генів
та від факторів довкілля. Принцип
взаємодії спадковості і середовища
лежить в основі розвитку
хвороб зі спадковою
схильністю, або мультифакторіальних
хвороб. Це велика, клінічне різноманітна
група патологічних проявів, але зі
схожими генетичними закономірностями
розвитку. Сюди приналежать уроджені
вади розвитку, серцевосудинні, психічні,
злоякісні та інші хвороби.
розвитку одних ознак визначальну роль
виконує генотип, а для розвитку інших
істотне значення має зовнішнє середовище.
Проте немає таких ознак, які абсолютно
зумовлювались би тільки спадковістю
або тільки середовищем. Ознаки, прояв
яких залежить від багатьох генів, більшою
мірою зазнають впливу екзогенних
чинників. Такі ознаки є мультифакторіальними
- за цих умов формування фенотипу
відбувається шляхом взаємодії спадковості
і середовища, генетичних і негенетичних
чинників. У людини такими ознаками є
зріст, розумові здібності, артеріальний
тиск, поведінкові реакції та ін. Дія
будь-якого гена залежить від інших генів
та від факторів довкілля. Принцип
взаємодії спадковості і середовища
лежить в основі розвитку
хвороб зі спадковою
схильністю, або мультифакторіальних
хвороб. Це велика, клінічне різноманітна
група патологічних проявів, але зі
схожими генетичними закономірностями
розвитку. Сюди приналежать уроджені
вади розвитку, серцевосудинні, психічні,
злоякісні та інші хвороби.
На відміну від моногенних ознак (хвороб), мультифакторіальному успадкуванню властива кількісна й якісна неоднорідність (рис. 1.133).
Аналіз такого розподілу показує, що імовірність захворювання серед родичів, яким властива генетична схильність, набагато вища, ніж у загальній популяції. Тобто пенетрантність також буде більшою. Якщо у всіх осіб даного генотипу з'являється очікуваний фенотип, то це вказує на повну пенетрантність. Неповна пенетрантність з'являється у випадку багатьох домінантних алелів і у випадку гомозиготності за рецесивними алелями. Функція пенетрантності виражає імовірність хвороби для кожного генотипу і залежить від таких чинників, як стать, вік, вплив навколишньго середовища й ефекту дії інших генів. Наприклад, ризик захворіти на рак грудної залози у віці 40, 55 і 80 років становить 37,66 і 85 % відповідно.
Сумарна дія генетичних і середовищних чинників зумовлює явище експресивності - ступеня прояву ознаки або хвороби. У нашому випадку - це крайні варіанти норми, субклінічні і типові форми хвороби.
Вирішальним критерієм захворювання є кількісне відхилення у функціях або ознаках, яким властивий широкий розмах мінливості серед населення.
Явище взаємодії генів на рівні фенотипу вивчає новий напрямок генетики, який отримав назву функціональна геноміка або протеоміка.
Фенокопії.
При проведенні медико-генетичного консультування слід пам'ятати про можливість виникнення фенокопій - тобто прояву фенотипу, схожого за характерними проявами мутантного гена, викликаного чинниками зовнішнього середовища.
Фенотипно однорідні аномалії можуть бути наслідком спадкових або середовищних факторів, які діють на ембріон. Так, уроджене помутніння кришталика може бути спадковим рецесивним захворюванням, результатом інфекції (корової краснухи) або впливом іонізуючого випромінювання в ранній період вагітності.
Вживання вагітною синтетичних гормонів, яким властивий маскулінізуючий ефект, може спричинити явище вірилізації плоду жіночої статі (надмірне ово-лосіння), яке нагадує адреногенітальний синдром.
Деякі ліки (талідомід) хімічної природи, які призначалися хворим вагітним, схильні до провокування вад розвитку (атрезія слухового ходу, аплазія променевої кістки, вади розвитку нирок, атрезія дванадцятипалої кишки, вади серця та ін.), які нагадують фенокопії домінантного синдрому Холта - Орама і рецесивної панмієлопатії Фанконі.
Хімічні речовини (етанол) порушують генетичні механізми формування систем органів, кінцівок, голови і мозку на пізніх стадіях ембріонального розвитку, спричиняють появу фенокопій, схожих на патологію спадкового генезу.
Все це докази взаємодії генетичних і негенетичних чинників у виникненні виродливостей та вад розвитку, зокрема фенокопій.
Хвороби із спадковою схильністю. Поняття про
мультифакторіальні хвороби.
Ряд захворювань і ознак зумовлені багатьма генами, тому їх позначають терміном полігенні. На їх розвиток великий вплив мають фактори зовнішнього середовища, тому їх називають мультифакторіальними.
Мультифакторіальні (полігенні) захворювання - це захворювання зі спадковою схильністю, у генезі яких поєднуються взаємодія спадкових і зовнішніх факторів.
Мультифакторіальні захворювання - це такі патологічні стани, для прояву яких необхідні дві умови:
Наявність спадкової схильності.
2. Несприятливі впливи зовнішнього середовища. Мультифакторіальні захворювання зпосідають одне з провідних місць серед хронічних неінфекційних хвороб людини. Вони відрізняються клінічним поліморфізмом. Складна взаємодія генетичних і зовнішньосередовищних факторів призводить до формування клінічної сталості мультифакторіальних захворювань в окремих родинах і популяції в цілому.
До мультифакторіальних захворювань належать: цукровий діабет, гіпертонічна хвороба, виразкова хвороба шлунка, атеросклероз, ішемічна хвороба серця, бронхіальна астма, шизофренія, епілепсія та ін.
Необхідно враховувати, що об'єктом спостереження лікаря будь-якої спеціальності повинен стати не тільки хворий, але й члени його родини. Лікар "загальної практики" і особливо сімейний лікар повинен уміти на підставі методів клініко-генетичного обстеження хворого і його родичів виявляти групи підвищеного ризику, в яких, у першу чергу, потрібно застосовувати відповідні профілактичні заходи.
Мультифакторіальні хвороби чи хвороби зі спадковою схильністю складають у даний час 92 % від загальної патології людини.
Механізм розвитку мультифакторіального захворювання - патологічний фенотип - виявляється тоді, коли сумарна дія генетичних і середовищних факторів досягає чи перевищує деяке граничне значення схильності.
Мультифакторіальні хвороби при всій їх розмаїтості характеризують деякі загальні риси:
Висока частота в популяції.
Існування клінічних форм, які утворюють не перервний ряд, від прихованих субклінічних до різко виражених проявів.
Більш ранній початок і деяке посилення клінічних проявів у наступних поколіннях.
Значні статеві і вікові розходження в попу ляційній частоті нозологічних форм.
Відносно низький рівень за маніфестними проявами хвороби у монозиготних близнюків (60 % і нижче) та істотно перевищуючий відповідний рівень у дизиготних близнюків.
6. Невідповідність закономірностей успадкування простим менделівським моделям. З огляду на різноманітні механізми, що впливають на розвиток і перебіг хвороби, не вдається простежити чітких закономірностей передачі захворювання з покоління до покоління. Аналіз родоводів при мультифакторіальних хворобах грунтується не на законах Менделя, а на емпірично отриманих даних.
7. Залежність ступеня ризику для родичів хворого від частоти хвороби в популяції (вона тим вища, чим рідше зустрічається дане захворювання); ризик зростає з народженням кожного наступного хворого, крім того, він підвищується в міру збільшення ступеня тяжкості хвороби пробанда.
8. Подібність клінічних та інших проявів хвороби в найближчих родичів пробанда, що віддзеркалює коефіцієнт спадковості (для полігенних хворобвін перевищує 50-60 %).
Мультифакторіальним захворюванням властиві певні закономірності успадкування. Так, ризик виникнення захворювання і вираженість клінічної картини залежать від:
1. Кількості уражених родичів і ступеня їх споріднення з пробандом.
2. Віку, в якому відбувається прояв захворювання в родині.
3. Клінічної його тяжкості.
4. Кількості осіб менш ураженої статі.
5. Частоти повторюваності даної патології в популяції.
Отже, при мультифакторіальних захворюваннях має місце зростання відповідних патологічних станів серед близьких родичів пробанда, нерідко з різною частотою ураження особин чоловічої чи жіночої статі. Чим вищий генетичний ризик (ступінь генетичного обтяження), тим менша інтенсивність зовнішньосередовищних впливів необхідна для прояву "граничного ефекту", тобто розвитку хвороби.
Група ризику при мультифакторіальних захворюваннях.
До даної групи відносять осіб з урахуванням величини спадкового обтяження, яка залежить від тяжкості захворювання, ступеня спорідненості з хворим і числа хворих у родині. Виявлення груп ризику за допомогою клініко-генеалогічного методу дозволяє
ефективно провести ранні лікувально-профілактичні заходи для осіб, схильних до різних захворювань.
Так, за наявності гіпертонічної хвороби в одного з батьків необхідно контролювати артеріальний тиск у дитини, рекомендувати щадний режим. У цих родинах потрібно якомога раніше пропагувати здоровий спосіб життя, а членам цих родин треба постійно займатися спортом, дотримуватися режиму праці і відпочинку, обмежувати споживання кухонної солі. Якщо такі звички розвиваються з дитинства, то вони можуть мати профілактичний ефект. Сімейний лікар повинен мати тісний контакт із родиною хворого. При першій фіксації підвищення артеріального тиску особу з групи ризику треба взяти на диспансерний облік. Особлива увага повинна приділятися родинам, члени яких схильні до таких хвороб, як цукровий діабет, виразкова хвороба, епілепсія, шизофренія.
Так, у випадку, якщо один з батьків хворий, то ризик народження дитини з цукровим діабетом дорівнює 10 %. Отже, існує реальна небезпека розвитку даного захворювання. Членів таких родин необхідно поставити на диспансерний облік, періодично проводити профілактичні огляди із застосуванням додаткових методів.
Література :
Основна : с. 192 – 194, с.195 – 198.
додаткова: Т.1 с. 220 – 224, с. 148 – 152.
Медична біологія
Лекція № 6 . Основи генетики людини. Методи вивчення
спадковості.
1. Предмет і завдання генетики людини та медичної генетики.
2. Методи вивчення спадковості людини.
3. Геніалогічний метод.
4. Близнюковий метод.
5. Популяційно-статистичний метод.
6. Цитологічний метод.
Самостійна робота № 7. Методи генетики людини: дерматогліфічний, імунологічний,
гібридизація соматичних клітин.