

Этап вдоха.
Поступление первого импульса к дыхательным мышцам.
Синаптическая передача нервных импульсов.
Увеличение объема грудной полости.
Увеличение объема легких.
Снижение давления воздуха в легких.
Всасывание воздуха из атмосферы в легкие.
Этап выдоха.
Расслабление дыхательных мышц вслед за сокращением их при вдохе.
Уменьшение объема грудной полости.
Уменьшение объема легких.
Повышение давления в легких.
Выдавливание воздуха из легких в атмосферу.
Энергия, затрачиваемая мускулатурой для осуществления свободного дыхания равна примерно 2 – 2,5 Дж, при этом 70% расходуется на преодоление эластичного сопротивления грудной клетки и легочной ткани, а остальные 30% идут на совершение работы по перемещению воздуха против сил трения. Силы трения пропорциональны скорости перемещения воздушной массы, следовательно второй компонент энергозатрат возрастает при учащении дыхания. Энергозатраты на преодоление эластичности органов дыхания практически не зависят от частоты дыхания. В то время как энергозатраты на эластичное сопротивление в основном определяются объемом воздуха, поступающего в легкие при вдохе, то есть глубиной дыхания. Неодинаковая зависимость каждого из компонентов работы дыхательного аппарата от частоты и глубины дыхания приводит к установлению оптимального соотношения между ними. При этом соотношении работа должна поддерживаться на минимальном уровне, при данном объеме вентиляции. Под объемом вентиляции понимают минутный объем дыхания. В условиях физиологического покоя глубина дыхания обычно составляет примерно 0,5 л, а частота 12 – 16 вдохов в минуту.
ФИЗИЧЕСКИЕ ОСНОВЫ ГАЗООБМЕНА.
В
легких совершается обмен
![]() между организмом и окружающей средой.
Благодаря работе дыхательных мышц
легочные альвеолы обмениваются этими
газами с атмосферой. Это обеспечивает
относительное постоянство
и
в альвеолярной газовой смеси. Газообмен
между альвеолярной газовой смесью и
кровью легочных капилляров происходит
на альвеолярно-капиллярной мембране
(АКМ). Схематично АКМ представляется:
между организмом и окружающей средой.
Благодаря работе дыхательных мышц
легочные альвеолы обмениваются этими
газами с атмосферой. Это обеспечивает
относительное постоянство
и
в альвеолярной газовой смеси. Газообмен
между альвеолярной газовой смесью и
кровью легочных капилляров происходит
на альвеолярно-капиллярной мембране
(АКМ). Схематично АКМ представляется:

Схема альвеолярно-капиллярной мембраны и диффузии газов в легких
1– альвеолярный эпителий, 2 – межклеточная среда, 3 – эндотелий капилляра, 4 – плазма крови, 5 – эритроцитарная мембрана
Газообмен в легких осуществляется под действием только одной силы – концентрационного градиента каждого из газов на АКМ. Массоперенос газов подчиняется уравнению Фика:
![]()
Используя уравнение Фика для количественной оценки газопереноса через АКМ целесообразно заменить градиент концентраций на градиент парциальных давлений и напряжений газов.
Парциальное давление газа – такое давление компонента газовой смеси, которое он оказал бы на заключающую его оболочку, если бы один занимал весь объем, предоставленный этой смеси.
Содержание газа в жидкости принято характеризовать напряжением. Под напряжением газа в жидкости понимают такое парциальное давление данного газа, которое нужно создать для поддержания равновесия в его содержании между газовой смесью и жидкостью. То есть для прекращения всякого газообмена между ними. Размерность парциального давления и напряжения – единица давления (Па – мм.рт.ст.).
![]() -парциальное
давление
со стороны альвеол.
-парциальное
давление
со стороны альвеол.
![]() -напряжение
в капиллярах (в крови).
-напряжение
в капиллярах (в крови).
Рассмотрим
величины парциальных давлений
и
![]() в альвеолах газовой смеси. С одной
стороны и объем в крови легочных
капилляров с другой. Это необходимо
для оценки сил под действием которых
совершается обмен через АКМ. В газовой
смеси легочных альвеол
в альвеолах газовой смеси. С одной
стороны и объем в крови легочных
капилляров с другой. Это необходимо
для оценки сил под действием которых
совершается обмен через АКМ. В газовой
смеси легочных альвеол
![]() мм.рт.ст.
или 13,7 кПа.
мм.рт.ст.
или 13,7 кПа.
![]() мм.рт.ст. или 5,3 кПа. Расчет парциальных
давлений в газовой смеси альвеол обычно
производят по формуле:
мм.рт.ст. или 5,3 кПа. Расчет парциальных
давлений в газовой смеси альвеол обычно
производят по формуле:
![]()
![]() -парциальное
давление газа.
-парциальное
давление газа.
![]() -давление
насыщенного газа паров воды в альвеолах
при t=36,7
-давление
насыщенного газа паров воды в альвеолах
при t=36,7
![]() ,
,
![]() =47
мм.рт.ст. или 6,3 кПа.
=47
мм.рт.ст. или 6,3 кПа.
![]() -атмосферное
давление.
-атмосферное
давление.
![]() -концентрация
данного газа в газовой смеси в %.
-концентрация
данного газа в газовой смеси в %.
Парциальному
давлению
и
в альвеолах газовой смеси противодействует
на АКМ напряжение этих газов в венозной
крови легочных капилляров как
![]() мм.рт.ст.
или 4,9 кПа,
мм.рт.ст.
или 4,9 кПа,
![]() мм.рт.ст. или 6,1 кПа . Из сопоставления
мм.рт.ст. или 6,1 кПа . Из сопоставления
![]() и
и
![]() каждого из газов следует, что
и
преодолевают АКМ по концентрационным
градиентам, но в противоположных
направлениях:
из альвеолярной газовой смеси в кровь,
а
наоборот.
каждого из газов следует, что
и
преодолевают АКМ по концентрационным
градиентам, но в противоположных
направлениях:
из альвеолярной газовой смеси в кровь,
а
наоборот.
РАСЧЕТ ГАЗООБМЕНА
Производится расчет не массы газа, переносимого через АКМ, а его объем. Тогда уравнение:
![]()
![]()
![]()
![]()
-длина, толщина АКМ.
Для полной оценки grad на АКМ необходимо знать толщину . АКМ состоит из двух слоев клеток:
эндотелия легочных капилляров,
альвеолярного эпителия
и базилярной мембраны, расположенной между ними. Общая толщина АКМ примерно 0,2 мкм, но в области ядерных клеток примерно 10 мкм.
Кроме перечисленных компонентов эта многомембранная система, через которую диффундирует газ. Легкие также включают пленку субфактата.
Основой субфактата является биомолекулярный липидный слой, подобный фосфолипидному слою биологической мембраны.
Наслоение субфактата на эпителиальную поверхность альвеол создает дополнительную к АКМ диффузионную среду, которую газы преодолевают при их массопереносе. За счет субфактата удлиняется диффузионный путь, что в свою очередь приводит к некоторому снижению концентрационных градиентов на АКМ. Однако без субфактата дыхание вообще было бы невозможным, так как стенки альвеол слиплись бы под действием значительного поверхностного натяжения, которое присуще альвеолярному эпителию. Субфактат снижает поверхностное натяжение альвеолярных стенок и предотвращает слипание альвеол.
Концентрационные градиенты и на АКМ резко падают при патологии. Они могут уменьшатся как при изменении разности концентраций в альвеолах или легочных капилляров, так и вследствие утончения АКМ. Первая причина характерна для изменения газовой среды, в которой находится организм (при пребывании человека в разряженной атмосфере), при нарушении легочного дыхания (при многих болезнях легких, при недостаточной работе дыхательных мышц). Вторая патология – отек легких. При этом жидкость, выходящая из капилляров скапливается в межклеточной среде (базилярная мембрана). Это приводит к утоньшению АКМ и резко снижает градиент концентрации газов на АКМ даже при нормальной разности их концентрации.
В
нормальных условиях диффузионный путь
составляет примерно 5 мкм. На этом
расстоянии перепад концентраций
и
неодинаков. Он различается более чем
на порядок и выражается величинами
парциального давления, составляющие
соответственно: для
примерно 70 мм. рт. ст. или 9,3 кПа, для
примерно
6 мм. рт. ст. или 0,8 кПа. Но при столь
значительных перепадах парциальных
давлений
и
массоперенос этих газов одинаков
![]() .
.
Причина
того заключается в том, что
и
имеют разную проницаемость через АКМ.
Проникающая способность каждого из
них оценивается коэффициентом диффузии.
Экспериментально установлено, что в
воде
![]() .
В воздухе на 4-5 порядков выше (
.
В воздухе на 4-5 порядков выше (![]() ).
В липиде
).
В липиде
![]() на
несколько порядков ниже по сравнению
с водой. Проникающая способность через
АКМ характеризуется
на
несколько порядков ниже по сравнению
с водой. Проникающая способность через
АКМ характеризуется
![]() . Это позволило заключить, что
проходит через АКМ сквозь поры,
заполненные водой. Также проходит через
АКМ и
. Это позволило заключить, что
проходит через АКМ сквозь поры,
заполненные водой. Также проходит через
АКМ и
![]() ,
но он делает это в 20-25 раз быстрее, чем
,
так как
лучше растворяется в воде. Высокая
степень растворимости
в воде обуславливается его малым
(близким к нулю) дипольным моментом.
Молекулы
не поляризованы благодаря своей
симметричной структуре. Молекула
не взаимодействует с заряженными
группами компонентов биологической
мембраны и легко проходит сквозь нее.
В этом отношении с высочайшей проникающей
способностью
связано его участие во многих процессах
которые обеспечивают регуляцию дыхания,
кровообращения, обмена веществ и других
функций организма.
,
но он делает это в 20-25 раз быстрее, чем
,
так как
лучше растворяется в воде. Высокая
степень растворимости
в воде обуславливается его малым
(близким к нулю) дипольным моментом.
Молекулы
не поляризованы благодаря своей
симметричной структуре. Молекула
не взаимодействует с заряженными
группами компонентов биологической
мембраны и легко проходит сквозь нее.
В этом отношении с высочайшей проникающей
способностью
связано его участие во многих процессах
которые обеспечивают регуляцию дыхания,
кровообращения, обмена веществ и других
функций организма.
ДИФФУЗИОННАЯ СПОСОБНОСТЬ ЛЕГКИХ. ТРАНСПОРТ И
Для
оценки газообмена в легких достаточно
трудно измерить коэффициент диффузии,
так как для его расчета нужно знать
величину площади диффузии, которую
невозможно измерить в каждом конкретном
случае, чаще при ее оценке ориентируются
на площадь альвеолярной поверхности.
В легких находиться приблизительно …
альвеол, каждая из которых имеет диаметр
от 75 до 300 мкм. При спокойном дыхании
объем альвеол в среднем изменяется на
2-3 %. При этих условиях полная альвеолярная
поверхность достигает 70-80![]() .
Но площадь через которую газы диффундируют
определяется не всей площадью дыхательной
поверхности альвеол, а только той
частью, где есть контакт с капиллярами,
наполненными движущейся кровью. Кровь
текущая по легочным капиллярам создает
тонкую пленку толщиной 10 мкм. Необходимо
рассчитать поверхность так называемой
кровяной плоскости, под которой понимают
суммарную поверхность эритроцитов,
контактирующими со всеми функционирующими
альвеолами в течение одной минуты.
Ежеминутно в состоянии покоя через
легкие протекает примерно пять литров
крови. В них содержится
.
Но площадь через которую газы диффундируют
определяется не всей площадью дыхательной
поверхности альвеол, а только той
частью, где есть контакт с капиллярами,
наполненными движущейся кровью. Кровь
текущая по легочным капиллярам создает
тонкую пленку толщиной 10 мкм. Необходимо
рассчитать поверхность так называемой
кровяной плоскости, под которой понимают
суммарную поверхность эритроцитов,
контактирующими со всеми функционирующими
альвеолами в течение одной минуты.
Ежеминутно в состоянии покоя через
легкие протекает примерно пять литров
крови. В них содержится
![]() эритроцитов.
Поверхность каждого из них
эритроцитов.
Поверхность каждого из них![]() .
Следовательно в состоянии покоя площадь
минутного газообмена в легких человека
составляет примерно 3750
.
При физических нагрузках площадь
увеличивается до 12000-15000
.
.
Следовательно в состоянии покоя площадь
минутного газообмена в легких человека
составляет примерно 3750
.
При физических нагрузках площадь
увеличивается до 12000-15000
.
Благодаря значительной поверхности газообмена, полный массоперенос и в легких человека осуществляется всего за 0,1 с. Вместе с тем, известно, что в условиях покоя каждый из эритроцитов проходит легочный капилляр от начала до конца за примерно 2с. При физической нагрузке скорость кровотока возрастает, что приводит, с одной стороны к увеличению поверхности газообмена, приходящемуся на единицу времени контакта крови и альвеолярной газовой смеси, а с другой стороны, к уменьшению продолжительности этого контакта. Согласно закону Фика, сокращение времени газообмена должно уменьшать объем газов диффундирующих через АКМ. Однако, это утверждение справедливо только для очень кратковременного (короче 0,1 с) газообмена. Когда же время кровотока по легочным капиллярам сокращается от 2 с. (в состоянии покоя) до 0,1 с. (в состоянии нагрузки), массоперенос газов в легких не уменьшается, а только увеличивается.
Приводимые
выше аргументы позволяют понять
насколько трудно оценить влияние на
газообмен человека таких параметров,
как диффузионная площадь S
и время газообмена. В этой связи вводится
своеобразная физическая величина,
которую называют диффузионной
способностью легких -
![]() Под
Под
![]() легких понимают объем данного газа,
переносимого через АКМ всей дышащей
поверхностью в течение 1 мин, при разности
парциальных давлений на мембране в 1
мм рт.ст. и нормальной толщине АКМ
(примерно 5 мкм). Другими словами,
дыхательная способность ДЛ определяется
произведением коэффициента диффузии
на площадь газообмена. ДЛ удается
измерить у человека и животных при
помощи специальных приборов. В медицине
принята внесистемная единица измерения
ДЛ =
легких понимают объем данного газа,
переносимого через АКМ всей дышащей
поверхностью в течение 1 мин, при разности
парциальных давлений на мембране в 1
мм рт.ст. и нормальной толщине АКМ
(примерно 5 мкм). Другими словами,
дыхательная способность ДЛ определяется
произведением коэффициента диффузии
на площадь газообмена. ДЛ удается
измерить у человека и животных при
помощи специальных приборов. В медицине
принята внесистемная единица измерения
ДЛ =![]()
Зная величину ДЛ и grad парциальных давлений для и , можно вычислить газоперенос каждого из газов через АКМ за 1 минуту:
![]()
![]()
![]() -
разность между парциальным давлением
и напряжением
;
-
разность между парциальным давлением
и напряжением
;
![]() -
разность между напряжением
и парциальным давлением.
-
разность между напряжением
и парциальным давлением.
При
изменении площади диффузии S,
значение ДЛ также меняется. Так, например,
в покое
![]() примерно
20-25,
примерно
20-25,
![]() примерно
600 . При тяжелой физической работе
достигает примерно 70, а примерно
1700.
примерно
600 . При тяжелой физической работе
достигает примерно 70, а примерно
1700.
Проникнув в кровь, текущую по легочным капиллярам, переносится ею во все ткани организма. Формы этого транспорта различны. Во-первых, растворяется в крови и, во-вторых, связывается с гемоглобином, который содержится в эритроцитах. Наилучшей растворимостью в крови обладает , а растворяется меньше.
Коэффициент растворимости газов существенно зависит от свойств растворителя. В крови газы растворяются хуже, чем в воде. Ухудшение растворимости объясняется главным образом тем, что кровь содержит соли и белки.
Зная
коэффициент растворимости
в
цельной крови при t
примерно
![]() ,
а также напряжения
в крови, оттекающей от легких (примерно
100 мм рт.ст. или 13,3 кПа), легко можно
рассчитать объем
,
растворяющегося в единице объема
артериальной крови. Подставив в уравнение
Генри соответствующие значения, можно
убедиться, что в 100 мл крови растворяется
0,3
,
а также напряжения
в крови, оттекающей от легких (примерно
100 мм рт.ст. или 13,3 кПа), легко можно
рассчитать объем
,
растворяющегося в единице объема
артериальной крови. Подставив в уравнение
Генри соответствующие значения, можно
убедиться, что в 100 мл крови растворяется
0,3
![]() :
:
![]()
![]() -
концентрация газа;
-
концентрация газа;
![]() -
коэффициент растворимости газа.
-
коэффициент растворимости газа.
Однако,
экспериментально установлено, что в
100 мл артериальной крови примерно
растворяется 20
![]() .
Различие обусловлено присутствием в
эритроцитах гемоглобина, каждый грамм
которого связывает и переносит примерно
1,39
.
.
Различие обусловлено присутствием в
эритроцитах гемоглобина, каждый грамм
которого связывает и переносит примерно
1,39
.
В каждых 100 мл крови обычно содержится 14-16 граммов гемоглобина, чем и определяется кислородная емкость крови, то есть, объем транспортируется децилитром крови (100 мл). При содержании гемоглобина в крови 14 г, кислородная емкость примерно 19 , а при 16 г гемоглобина – примерно 22 .
лучше растворяется в крови, в силу чего его объем растворяемый в 100 мл крови составляет примерно 2,4 . В такой форме транспортируется примерно 5% всего , переносимого кровью. Кроме этого, 15% кровь переносит в виде карбамино-гемоглобина (соединения с гемоглобином), но наибольшая часть (примерно 80%) транспортируется кровью в виде бикарбонатов.
Азот переносится кровью только в растворенном состоянии, причем, в каждых 100 мл содержится всего 1 мг азота. Несмотря на столь ничтожное количество, все ткани организма насыщены азотом, так как молекулы азота не участвуют в метаболических процессах.
ЭЛЕМЕНТЫ БИОФИЗИКИ
ОРГАНОВ ЧУВСТВ
Организм представляет собой сложную самоорганизующуюся систему, которая характеризуется обменом веществ, энергией и информацией с окружающей средой. Целесообразное реагирование организма, при постоянно меняющихся условиях внешней среды, возможно только при наличии непрерывного поступления в организм информации от окружающей среды. Функцию получения и переработки информации об условиях внешней среды в организме выполняют органы чувств. По существу, органы чувств представляют собой измерительные устройства для анализа внешних физических стимулов, а также для оценки эффективности действий, производимых организмом. Таким образом, органы чувств выполняют роль обратной информационной связи в системе «организм-среда». В 1968 г. Мэчи предложил следующую схему, показывающую взаимодействие организма с внешней средой. Схема имеет вид:
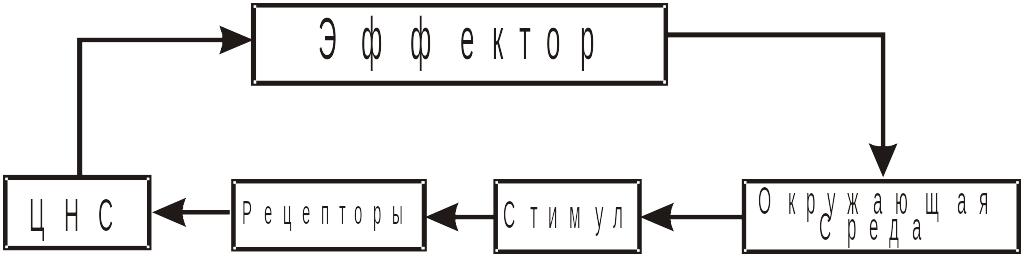
Схема, показывающая взаимодействие организма и внешней среды (по Мэчину)
При исследовании работы органов чувств представляют интерес два аспекта:
кибернетический;
биофизический.
Кибернетический аспект заключается в изучении принципов кодирования и передачи информации в органах чувств.
Биофизический аспект заключается в исследовании конкретных физико-химических процессов взаимодействия факторов внешней среды с органами чувств, что приводит к трансформации энергии внешнего воздействия в специфические сигналы, пригодные для анализа нервной системой.
При действии внешнего стимула на органы чувств у человека, возникает специфическое ощущение. Зависимость между величиной ощущения и величиной действующего стимула сформулирована в законе Вебера-Фехнера. В 1760 г. Буггер установил, что соотношение минимального воспринимаемого изменения освещенности к величине освещенности есть величина постоянная в широких пределах освещенности:
![]()
Такое
же постоянство отношения минимально
воспринимаемого прироста раздражения
(![]() )
к его исходной величине (R)
было установлено Вебером и для других
сенсорных систем, то есть:
)
к его исходной величине (R)
было установлено Вебером и для других
сенсорных систем, то есть:
![]()
Так,
например, прирост в весе груза, чтобы
стать ощутимым, должен превышать ранее
действующий вес на 3%. Фехнер предположил,
что минимальный прирост ощущения (![]() )
относительно исходного уровня ощущения
(S)
также постоянен, тогда:
)
относительно исходного уровня ощущения
(S)
также постоянен, тогда:
![]() ,
где k-
коэффициент пропорциональности.
,
где k-
коэффициент пропорциональности.
Интегрируя это уравнение и проведя алгебраические преобразования, Фехнер получил следующее выражение:
S = a lg R + b, где a, b – постоянные.
Таким образом, согласно закону В-Ф, величина ощущения S пропорциональна log силы раздражения. Эта зависимость обусловлена принципом кодирования информации в рецепторном аппарате органов чувств.
Рецепторы представляют собой или специфические окончания афферентных нервных волокон, или нервные окончания в соединении со специализированными клетками, или структурами. Во всех случаях рецептор действует, как преобразователь энергии раздражения в энергию нервного ответа.
При действии внешнего стимула изменяется ионная проницаемость рецепторных мембран, что вызывает их деполяризацию. Это возникает под действием стимула и деполяризация называется генераторным потенциалом (ГП). Величина ГП пропорциональна log интенсивности действующего раздражителя. ГП рецептор клетки генерирует в афферентном нервном волокне ПД, который служит сигналом, передающим информацию в нервной клетке и нервной системе. Между величиной ГП и частотой появления ПД в афферентном волокне, наблюдается линейная зависимость.
Нервное волокно может находиться в двух состояниях:
1 – в возбужденном, когда есть ПД;
2 – в невозбужденном, когда ПД отсутствует.
Таким образом, в нервной системе имеется дискретная двоичная форма кодирования информации, как в цифровых ЭВМ. Как показывает эксперимент, информация в нервной системе кодируется не последовательностью ПД, а их частотой появления, как в аналоговых вычислительных машинах, где различные модулированные показатели, представленные аналогами, изменяются пропорционально величине модулированных факторов.
Поскольку между интенсивностью стимула и величиной ГП существует логарифмическая зависимость, а ГП связан с частотой ПД линейно, интенсивность стимула и частота ПД должны быть связаны логарифмической функцией. Работы Метьюса, Грэма и других, выполненные с помощью микроэлектронной техники, подтверждают, что частота пропорциональна логарифму величины действующего раздражителя.
![]() -
частота ПД.
-
частота ПД.
m,n=const.
Таким образом, по-видимому, данная зависимость преобразования интенсивности действующего стимула (R) в частоту нервных импульсов в афферентных волокнах и обуславливает логарифмическую зависимость величины ощущения от интенсивности раздражителя.
ЭЛЕМЕНТЫ БИОФИЗИКИ СЛУХА
ФУНКЦИИ ВСПОМОГАТЕЛЬНЫХ ЭЛЕМЕНТОВ
НАРУЖНОГО И СРЕДНЕГО УХА
В органе слуха принято выделять три части:
наружное ухо;
среднее ухо;
внутреннее ухо.
Упрощенную схему органа слуха можно представить в виде:
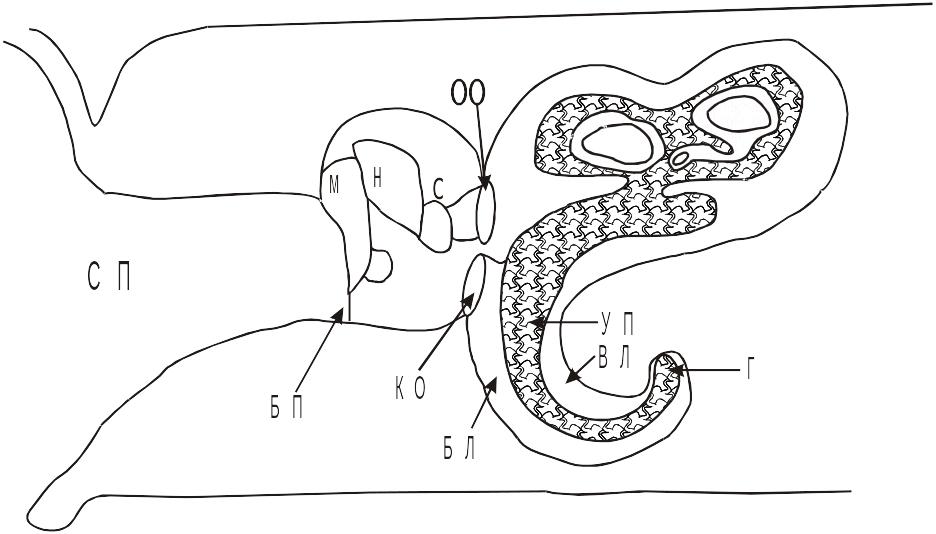
Схема органа слуха.
СП – слуховой проход; БП – барабанная перепонка; М – молоточек; Н – наковальня;
С – стремечко; ОО – овальное окно; ВЛ – вестибулярная лестница;
УП – улитковый проток; БЛ – барабанная лестница; Г – геликотрема;
КО – круглое окно; НУ – наружное ухо; СУ – среднее ухо.
Слуховые рецепторы расположены во внутреннем ухе, тогда, как вспомогательные элементы, способствующие слуховой рецепции, присутствуют во всех частях органа слуха.
Наружное ухо включает ушную раковину и слуховой проход. С ним связано свойство направленности слухового восприятия, под которым понимают лучшее восприятие звуков, идущих в определенном направлении. Это обеспечивает локализацию источника звука в пространстве (ототолику), кроме этого, слуховой проход служит резонатором. Его резонансная частота у человека лежит в области частот примерно 3 кГц, но резонансная кривая имеет уплощенную форму, поскольку добротность СП, как колебательной системы, невелика. Разница в коэффициенте передачи звуков резонансной и не резонансной частот не более 1 порядка. Границей между наружным и средним ухом служит БП. НУ защищает БП от механических повреждений и, поддерживая определенные микроклиматические условия (температуру, влажность и пр.), стабилизирует ее механические свойства. Таким образом, НУ выполняет следующие функции:
Локализация звука в пространстве.
Усиление звукового сигнала (резонансное).
Механическая защита БП.
Обеспечение микроклимата, что необходимо для нормального функционирования БП.
По
СУ звук проводится цепочкой слуховых
косточек: М, Н, С. рукоятка М зафиксирована
на БП, а его головка образует сустав с
Н, который сочленяется с С. Основание
С погружено в ОО, которое служит входом
во ВУ, поэтому система БП и слуховых
косточек СУ выполняет роль связующего
звена между атмосферой и ВУ, заполненным
жидкостью. Если бы звук переходил из
воздуха в жидкость непосредственно,
то интенсивность звуковой волны падала
бы из-за сильного отражения звука на
границе раздела сред с различным
акустическим импедансом [![]() ].
Жидкость обладает большей
по сравнению с воздухом. БП (исключает)
и система слуховых косточек обеспечивает
согласование
воздуха и жидкости ВУ, вследствие чего
уменьшаются потери интенсивности
звуковой волны при переходе из атмосферы
во ВУ.
].
Жидкость обладает большей
по сравнению с воздухом. БП (исключает)
и система слуховых косточек обеспечивает
согласование
воздуха и жидкости ВУ, вследствие чего
уменьшаются потери интенсивности
звуковой волны при переходе из атмосферы
во ВУ.
Согласование и усиление звука обеспечивается благодаря двум обстоятельствам:
Поверхность БП примерно в 25 раз больше площади ОО, за счет разницы в площадях на входе и выходе системы механической передачи происходит усиление давления на жидкость ВУ, относительно звукового давления, воздействующего на БП.
Слуховые косточки так расположены в цепочке, что при их движении, они образуют рычаги силы и увеличивают давление на основание С по сравнению с давлением на рукоятку М. Вся система передачи звука в СУ работает наподобие гидравлического пресса с примерно 90 кратным выигрышем в силе.
Максимальный коэффициент усиления давления характерен для звуковых волн с f примерно 1 кГц. Звуковые колебания, как меньших, так и больших f, усиливаются слабее, так как для них слуховые косточки СУ хуже согласовывают , и часть звуковых волн с f примерно 1 кГц, при их средней интенсивности, передается на ВУ практически без потерь. Однако, и для них коэффициент передачи может снижаться и оказывается даже меньше 1. Это имеет место в том случае, когда звук становится слишком интенсивным. Это сопровождается ощущение покалывания, а затем и боли в ушах. Понижение коэффициента передачи происходит при помощи тоненьких мышц, которые прикрепляются к М и С. В ответ на сильные звуки, они рефлекторно сокращаются и уменьшают подвижность системы слуховых косточек.
Итак, систему передачи звука, сосредоточенную в СУ, можно считать механическим преобразователем (усилителем), обладающим переменными регулируемым коэффициентом передачи давления с БП на жидкость ВУ. При разрушении слуховых косточек слух полностью не теряется, но понижается примерно в 1000 раз.
СУ выполняет следующие функции:
Связующую между атмосферой и ВУ.
Согласования атмосферы (воздуха) и жидкости ВУ.
Усиление звукового сигнала.
Защитная, при больших интенсивностях звука.
СТРОЕНИЕ ВНУТРЕННЕГО УХА
Внутреннее ухо (ВУ) человека размещается в пирамидальной части височной кости. Часть этого причудливого лабиринта, содержащая слуховые рецепторы, напоминает улитку в форме конусообразной спирали, образующей 2,5 витка. Схематично костную часть улитки можно представить в виде:
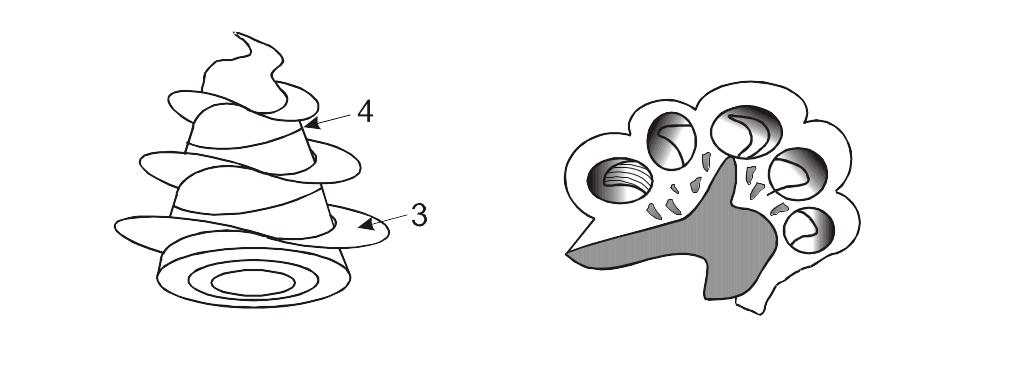
a б
Костная улитка.
а – стержень улитки; б – улитка в разрезе
Внутри улитки имеется просвет с круглым поперечным сечением, повторяющим ее спиралевидное закручивание вокруг костного стержня. От стержня в просвет улитки выступает костная спиральная пластинка (КСП), которая частично разделяет просвет примерно на 1/3-1/2 на всем протяжении спиралевидного хода. Поперечный разрез улитки имеет следующий вид:
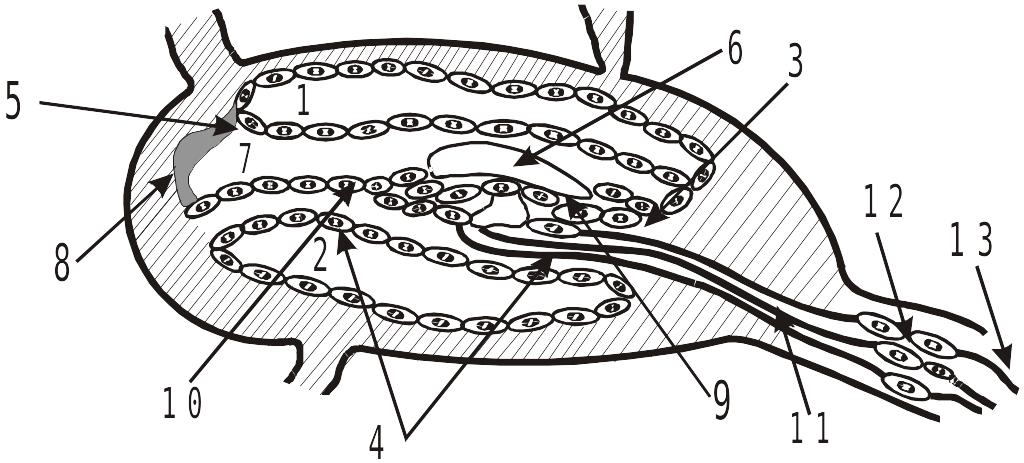
Поперечный разрез улитки
1 – вестибулярная лестница, 2 – барабанная лестница, 3 – костная спиральная пластинка,
4 – базилярная мембрана, 5 – мембрана Рейснера, 6 – покровная пластинка (мембрана),
7 – улитковый проток, 8 – сосудистая полоска, 9 – волосковые клетки внутреннего ряда, 10 – волосковые клетки наружного ряда, 11 – нервные окончания, 12 – биополярные нейроны, 13 – аксоны слухового нерва
Между
выступом костной спиральной пластинки
(3) и противолежащей поверхности
улиткового хода натянута базилярная
мембрана (4), которая образована мягкими
тканями БМ и дополняет костную спиральную
пластинку, чем достигается полное
разделение просвета улитки. Кроме нее,
от костной спиральной пластинки отходит
вестибулярная (преддверная) мембрана,
которая называется мембраной Рейснера
(5). Эта мембрана образует острый угол
с базилярной мембраной, и она также
направлена к наружной стенке улитки.
Благодаря существованию БМ и ВМ,
спиралевидный ход внутри улитки разделен
на три параллельные канала, каждый из
которых свернут в спираль. Один из них
(нижний) расположен между стенкой улитки
и БМ. Он называется БЛ, другой канал –
ВЛ (1) заключен между верхней поверхностью
улиткового хода и мембраной Рейснера.
БЛ и ВЛ сообщаются на верхушке улитки.
Место перехода одной лестницы в другую
называют отверстием улитки или
геликотремой (Г). Третий канал клиновидного
сечения называется улитковым протоком
(7), он расположен между БМ и мембраной
Рейснера. Его третью стенку выстилает,
так называемая, сосудистая полоска
(8), с функционированием которой связано
поддержание определенного химического
состава жидкости – эндолимфы. Эндолимфа
заполняет улитковый проток, и она по
своим свойствам существенно отличается
от перелимфы, находящейся в БЛ и ВЛ.
Перелимфа сходна с межклеточной
жидкостью, тогда как эндолимфа скорее
напоминает цитозоль (по содержанию
![]() ,
вязкости и т.д.).
,
вязкости и т.д.).
В эндолимфу погружены специальные клетки слуховых рецепторов, которые входят в состав Кортиева органа. Их опекальный полюс увенчан волосковоподобными выростами плазмолеммы (цилии). В этой связи эти клетки называются волосковыми клетками.
Наряду со слуховыми рецепторами в Кортиевом органе присутствуют разнообразные опорные клетки, относящиеся к вспомогательным элементам ВУ. Кортиев орган расположен на БМ, причем волосковые клетки ориентированы перпендикулярно этой мембране. В каждой из двух улиток уха человека содержится по 15500 волосковых клеток, из которых 3500 образуют внутренний ряд (9) и 12000- (3) наружных ряда (10). По форме волосковые клетки внутреннего ряда подобны эллипсоиду (1), вертикальная ось которого составляет в среднем примерно 40 мкм, а горизонтальная примерно 10 мкм. Наружная волосковая клетка имеет цилиндрическую форму (2).
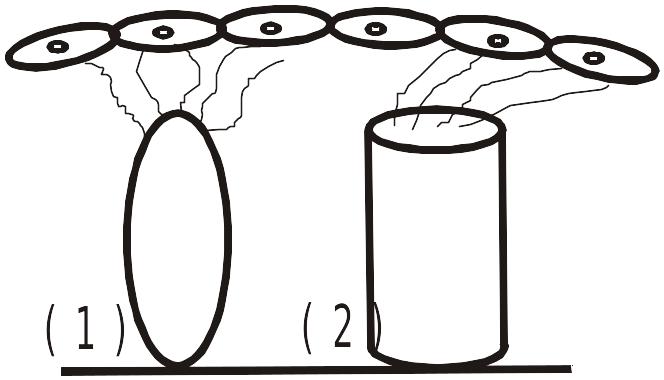
Волосковые клетки
1 – внутреннего ряда, 2 – наружного ряда
На базальном полюсе волосковой клетки сосредоточены синопсы с терминальными веточками афферентных волокон, являющиеся периферическими отростками нейронов, тела которых находятся в спиральном ганглии (12). Центральные отростки этих биополярных нейронов формируют слуховой нерв (13), по которому сигналы от слуховых рецепторов поступают в головной мозг. Каждая волосковая клетка внутреннего ряда образует синопсы примерно с 10 нервными окончаниями, но каждое из окончаний контактирует только с одной клеткой. Афферентные волокна, иннервирующие волосковые клетки наружных рядов, сильно ветвятся, и каждое из них имеет синоптические входы от многих таких клеток. В этой связи, несмотря на обилие волосковых клеток в наружных рядах Кортиева органа, основная часть (примерно 95%) афферентных волокон слухового нерва млекопитающих отводит сигналы от волосковых клеток внутреннего ряда, которые и преобразуют энергию механических колебаний.
РАСПРЕДЕЛЕНИЕ МЕХАНИЧЕСКИХ КОЛЕБАНИЙ
ВО ВНУТРЕННЕМ УХЕ
Под действием звука основание С колеблется, то вдвигаясь внутрь улитки, то выходя из нее. Таким образом энергия механических колебаний передается перелимфе ВЛ. Перелимфа может колебаться, благодаря двум обстоятельствам:
Жидкость, заполняющая ВЛ и БЛ, составляет одно целое, поскольку лестницы сообщаются на верхушке улитки, посредством Г.
Жесткий (костный) каркас, в который заключена гидравлическая система внутреннего уха, имеет окно не только на входе (ОО), но и на дальнем конце в БЛ (КО). КО затянуто мембраной. Оба окна «смотрят» в полость СУ. Поскольку мембрана КО обладает упругостью (эластичностью), начальное смещение перелимфы около ОО создает механическую волну, распределяющуюся вдоль улиткового протока, который заполнен эндолимфой.
Среда, в которой распределяются механические колебания внутри улитки – неоднородна. Она включает перелимфу, эндолимфу, разнообразные мембраны (БМ, ВМ и покровную мембрану), которые сами по себе являются композиционными материалами. Очень сложной биомеханической системой является БМ, с расположенным на ней Кортиевым органом. Инерционность БМ увеличивается от основания улитки к верхушке, тогда, как наиболее упругим ее участком является начальный (около ОО), а по направлению к Г эластичность понижается на два порядка.
Механические колебания внутри уха до настоящего времени не удалось описать математически, однако, при световой микроскопии даже небольшой части Кортиевого органа удалось наблюдать сложные колебания различных групп волосковых клеток. В целом, колебательный процесс, возникающий во внутреннем ухе под действием звуков, имеет характер бегущей волны. Волны такого типа образуются в нелинейной механической системе, являясь результатом интерференции различных колебаний, в которых приводятся различные элементы неоднородной механической среды. Другими словами, одно и то же воздействие вызывает неодинаковые колебания (с различными фазами и амплитудами, которые по-разному направлены) того или иного элемента нелинейной механической системы, в зависимости от его инерционных и упругих свойств. Вместе с тем, такие колебания когерентны, и, накладываясь друг на друга, они интерферируют. В результате их интерференции и образуется бегущая волна. В отличие от волновых процессов в однородной среде, бегущей волне не всегда свойственно максимальное смещение в начальном участке ее распределения и последующее угасание с определенным декрементом. Бегущая волна достигает своей максимальной амплитуды на различном удалении от входа в нелинейную механическую систему, а именно, в той области, где образуется интерфереционный максимум, при суперпозиции различный колебаний, которые возникают в неоднородной среде. Необходимо отметить, что местоположение максимального смещения в композиционной среде, при периферическом воздействии на нее механических сил, зависит от частоты сигнала.
Бегущая волна, возникающая в БМ под действием звуков НЧ, достигает максимального значения (амплитуды) ближе к ОО, тогда как звуки НЧ вызывают максимальное смещение тех участков БМ, которые находятся около Г. Звуковые тоны f = 1,6 кГц максимально смещают середину БМ человека. Таким образом, БМ, как композиционная среда с нелинейными механическими свойствами, функционирует наподобие системы фильтров, в которых расположение максимального отклика зависит от частоты звукового тона.
СЛУХОВАЯ РЕЦЕПЦИЯ
Благодаря
зависимости, местоположение максимальной
амплитуды бегущей волны на БМ от частоты
вызывающей ее звукового тона, происходит
распределение этих частот между
различными участками Кортиевого органа.
Его волосковые клетки возбуждаются,
главным образом, в местах максимального
смещения БМ, следовательно, звуковой
тон каждой частоты возбуждает
соответствующие слуховые рецепторы.
В этой связи в У осуществляется первый
этап частотного анализа звука, который
основывается на пространственном
разграничении участков БМ, колеблющихся
с неодинаковой амплитудой, под действием
звукового тона определенной частоты.
Звуки пороговой интенсивности, вызывающие
бегущие волны, амплитуда которых в
Кортиевом органе не превышает
![]() м. При усилении звуков, амплитуда
колебаний БМ больше этой величины всего
на 1-2 порядка, следовательно, слуховой
рецептор реагирует на ничтожные
механические перемещения. Начало этой
реакции связано с движением волосков
(цилий) на опекальном полюсе волосковой
клетки эти смещения обусловлены
распределением бегущей волны по БМ.
Среди цилий в каждой внутриволосковой
клетке различают одну длинную (киноцилия)
и множество (примерно 100-120) коротких
(стереоцилий). Рецепция звука обусловлена
воздействием механических усилий на
стереоцилии.
м. При усилении звуков, амплитуда
колебаний БМ больше этой величины всего
на 1-2 порядка, следовательно, слуховой
рецептор реагирует на ничтожные
механические перемещения. Начало этой
реакции связано с движением волосков
(цилий) на опекальном полюсе волосковой
клетки эти смещения обусловлены
распределением бегущей волны по БМ.
Среди цилий в каждой внутриволосковой
клетке различают одну длинную (киноцилия)
и множество (примерно 100-120) коротких
(стереоцилий). Рецепция звука обусловлена
воздействием механических усилий на
стереоцилии.
Стереоцилии прикасаются к покровной мембране, которая имеет желеобразное состояние. Один край покровной мембраны закреплен на костной спиральной пластинке, а другой, свободный, нависает над свободными клетками. Модель изгиба стереоцилий под действием звуковых тонов, схематически можно представить в виде:
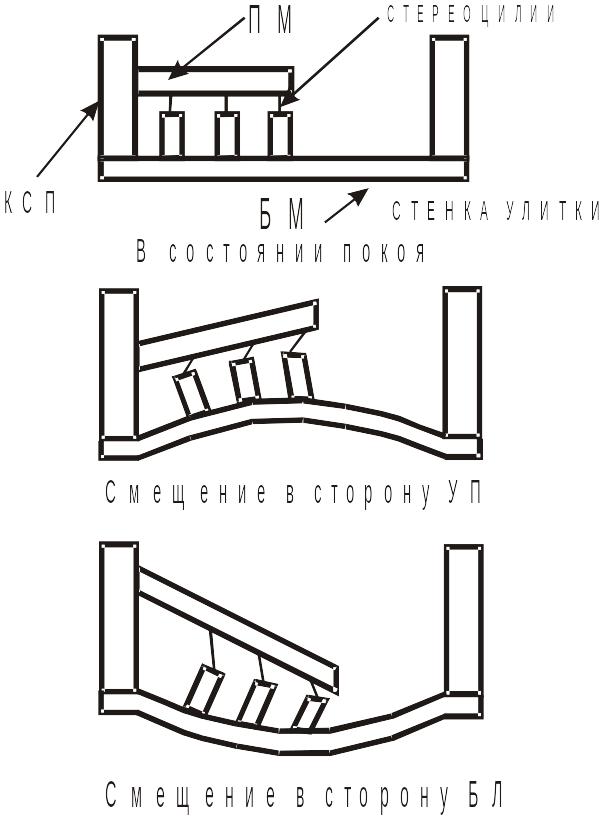
Модель изгиба стереоцилий под действием звука
КСП – костная спиральная пластинка; БМ – базилярная мембрана;
ПМ – покровная мембрана; СУ – стенка улитки, к которой прикрепляется базилярная мембрана; С – стереоцилии волосковой клетки
При совместных колебаниях БМ и ПМ происходит изгиб стереоцилий. Небольшое смещение их свободных концов приводит к значительной конформационной перестройке молекул плазмолеммы волосковой клетки в тех местах, где залегают «корни» изогнувшихся волосков, поскольку стереоцилиям присущи свойства микроочагов.
Конформации мембраных макромолекул обусловливают изменение проницаемостей для ионов и, следовательно, приводят к возникновению ионного тока через мембрану волосковой клетки. Это сопровождается сдвигом разности потенциалов на мембране, когда стереоцилии изгибаются в сторону киноцилий, рецепторная клетка деполяризуется. При их противоположном смещении наблюдается гиперполяризация. Только деполяризация волосковой клетки обеспечивает усиление импульсации в слуховом нерве, поэтому, именно деполяризационный сдвиг мембранного потенциала называется рецепторным потенциалом волосковой клетки – РП.
Уровень ПП волосковой клетки относительно межклеточной среды (перелимфы), находится в пределах от -50 до –70 мВ. Значение рецепторного потенциала зависит от интенсивности звука, и его величина составляет примерно 10 мВ. Максимальная величина РП, зарегистрированного с помощью микроэлектронной техники, составляет примерно 24 мВ. РП электротонически распределяется по плазмолемме волосковой клетки от ее опекального полюса к базальному. Там деполяризация приводит к высвобождению медиатора (определенное химическое вещество), поступающего далее через синоптическую щель на субсиноптическую мембрану, которая принадлежит чувствительному нервному окончанию, контактирующему с данной волосковой клеткой. Под действием медиатора, на
субсиноптической мембране возникает генераторный потенциал – ГП. Этот ГП электротонически распространяется на внесиноптические участки афферентных волокон, где вызывает образование ПД. В таком виде нервная импульсация направляется в ЦНС по слуховому нерву. Следовательно, по механизму преобразования адекватного стимула (звука) в нервные импульсы, слуховой рецептор является типичным представителем вторичночувствующих рецепторов.
Каждая волосковая клетка соединена с нервными окончаниями, при этом, каждое волокно слухового нерва начинается от узкого ограниченного участка Кортиевого органа, в отдельных случаях, от одной волосковой клетки. Поскольку слуховые рецепторы расположены в том или ином месте БМ возбуждается звуками определенных частот, то каждая небольшая группа нервных волокон слухового нерва проводит импульсы в ответ на звуки преимущественно одной частоты. Эта частота называется характерической частотой волокна. Если звук представляет собой сложные колебания, то в слуховом нерве активизируются все волокна, характерические частоты которых соответствуют гармоническому спектру сложного звука. Следовательно, на уровне слуховых рецепторов звуки разлагаются в гармонический спектр. Длительность звукового сигнала кодируется временем активации афферентных волокон, которые входят в состав слухового нерва.
РОЛЬ ЭНДОКОХЛИАРНОГО ПОТЕНЦИАЛА (ЭКП)
В СЛУХОВОЙ РЕЦЕПЦИИ
Между эндолимфой и перелимфой в У поддерживается разность потенциалов в пределах от 60 до 80 мВ. Эту разность называют ЭКП. Эндолимфа несет положительный потенциал относительно перелимфы. Источником ЭКП служит сосудистая полоска, выстилающая наружную стенку улиткового протока (8). Падение ЭКП наблюдается при нарушении кислородного снабжения сосудистой полоски, или при нарушении этой полоски (механическое повреждение).
Клетки, входящие в состав сосудистой полоски, содержат очень крупные митохондрии. Эта структурная особенность сосудистой полоски в совокупности с данными о роли ее в клеточном дыхании для поддержания разности потенциалов между эндолимфой и перелимфой, позволяют высказать предположение, что ЭКП по своей природе может быть аналогичен протонному потенциалу на митохондриальной мембране.
Звуки вызывают колебания ЭКП, амплитуда которых не превосходит 1% его постоянного уровня, то есть, эти колебания составляют 0,6-0,8 мВ. Это явление получило название микрофонного эффекта улитки, а сами колебания ЭКП принято называть микрофонными потенциалами. Микрофонный эффект У положен в основу телефонной теории слуха, согласно которой, частотный анализ звуков связан не с механическими колебаниями структур ВУ, а с возбуждением слуховых рецепторов микрофонными потенциалами. Частота этих потенциалов совпадает с гармоническим спектром звуковых тонов. В настоящее время ЭКП рассматривается, как важное условие нормального функционирования слуховых рецепторов, но не ведущий ее механизм.
Благодаря существованию ЭКП, поддерживается очень большая разность потенциалов (до 150 мВ) между эндолимфой и цитоплазмой волосковой клетки. Эта разность образуется в результате суммирования ПП волосковой клетки (разности потенциалов между ее цитоплазмой и перелимфой) и ЭКП.
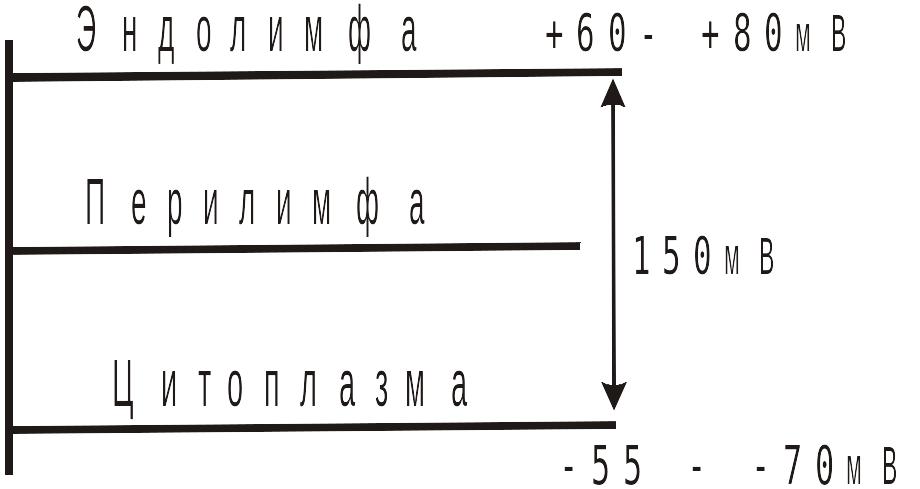
Уровень электрических потенциалов волосковой клетки
По существу, разность потенциалов между эндолимфой и цитоплазмой волосковой клетки выполняет роль ее ПП, относительно которой генерируется рецепторный потенциал. Столь значительный ПП не имеет ни одна другая клетка в организме человека. Это высокая крутизна переднего фронта ЭКП делает плазмолемму волосковой клетки чрезвычайно нестабильной и способной давать резкие и быстрые сдвиги мембранного потенциала, при малых изменениях электрического сопротивления мембраны, которые происходят в ответ на незначительные смещения стереоцилий.
ОСНОВЫ ФИЗИОЛОГИЧЕСКОЙ АКУСТИКИ
Физической акустике принято сопоставлять объективные (физические) и субъективные (психологические) характеристики звука.
Физические характеристики звука |
Психологические характеристики звука |
а) основная гармоника; б) высшие гармоники. |
1. Акустический спектр, характеризуется: а) основной тон (высота); измеряется в [мел] или [барк]; б) обертоны (тембр). |
2.
Интенсивность (плотность потока
звуковой энергии), обозначается
|
2. Громкость, измеряется в [сон]. |
Звук,
при распределении в среде представляет
собой волны сгущения и разряжения ее
частиц. В участках сгущения давление
больше, чем в участках разряжения.
Амплитуда периодических колебаний
давления в среде, происходящих под
действием распространяющего звука,
называют звуковым давлением – р. Оно
имеет размерность [Па] или [![]() ]
Звуковым давлением определяется
интенсивность звука, которая определяется
так:
]
Звуковым давлением определяется
интенсивность звука, которая определяется
так:
![]()
P – давление;
v – скорость распределения звука в данной среде;
– плотность среды [![]() ].
].
![]() – акустический
импеданс среды.
– акустический
импеданс среды.
Кроме того, I, как вектор Умова, можно рассчитать по формуле:
![]() ,
если известна w
– объемная плотность энергии звука,
то есть, энергия, приходящаяся на единицу
объема среды, в которой звук
распространяется. Для установления
взаимосвязи между интенсивностью и
громкостью, вводятся еще две физические
величины, которые характеризуют звук:
,
если известна w
– объемная плотность энергии звука,
то есть, энергия, приходящаяся на единицу
объема среды, в которой звук
распространяется. Для установления
взаимосвязи между интенсивностью и
громкостью, вводятся еще две физические
величины, которые характеризуют звук:
Уровень интенсивности – L.
Уровень громкости – Е.
L, или уровень звукового давления, определяется по формуле:
![]()
![]() -
эталонная интенсивность;
-
эталонная интенсивность;
- любой исследуемый звук.
обычно
выбирают меньше
![]() Вт/
Вт/![]() .
Такая интенсивность чистого тона, f
= 1 кГц, установлена, как средний порог
слуховой чувствительности для человека.
Эта интенсивность соответствует
звуковому давлению, равному
.
Такая интенсивность чистого тона, f
= 1 кГц, установлена, как средний порог
слуховой чувствительности для человека.
Эта интенсивность соответствует
звуковому давлению, равному
![]() Па. При к = 1, единицей уровня интенсивности
L
служит [Б] бел. При к = 10, единицей уровня
интенсивности L
служит [дБ], децибел. В медицине принято
использовать децибельную шкалу. Если
Па. При к = 1, единицей уровня интенсивности
L
служит [Б] бел. При к = 10, единицей уровня
интенсивности L
служит [дБ], децибел. В медицине принято
использовать децибельную шкалу. Если![]() ,
то L
= 0, следовательно, 3 величины
,
то L
= 0, следовательно, 3 величины
![]() и
,
и L=0
дБ – характеризуют один и тот же звук.
При
и
,
и L=0
дБ – характеризуют один и тот же звук.
При
![]() ,
L
принимает положительно значение, при
,
L
принимает положительно значение, при
![]() ,
L
< 0. Чем меньше L,
тем лучше (выше) слуховая чувствительность.
Человек лучше всего слышит звуковые
тоны с f
от 3 до 4 кГц.
,
L
< 0. Чем меньше L,
тем лучше (выше) слуховая чувствительность.
Человек лучше всего слышит звуковые
тоны с f
от 3 до 4 кГц.
Уровень громкости (Е) – это выражение различий в восприятии человеком звуков разной частоты. Уровнем громкости звука данной f называют уровень интенсивности звукового тона с f = 1 кГц, громкость которого при сравнении на слух эквивалентна громкости исследуемого звука.
Размерность физической величины Е – фон, - для звуков, частотой f = 1 кГц, значение Е и L совпадают. Так, если первая из них L составляет 0 дБ, то Е = 0 фонов (на f = 1 кГц). Средний порог слухового восприятия у человека составляет примерно 4 фонов. Болевые ощущения в ухе возникают под действием звуков, уровень громкости которых достигает примерно 130 фонов. Таким образом, все, что слышит человек, ограничивается звуковыми тонами, частота которых находится в диапазоне от 16 Гц до 20 кГц, а Е – от 4 до 130 фонов.
Громкость звука является психологической (субъективной) характеристикой. Она оценивается по шкале сонов. Для выражения громкости того или иного звука в сонах, за эталон принимается звуковой тон с f = 1 кГц, при уровне интенсивности L = 40 дБ. Дав испытуемому послушать эталонный тон, врач предъявляет ему тестированный звук, при этом, он просит определить, во сколько раз тестируемый звук громче или тише эталонного. Так, например, громкость тестируемого звука, если он слышит в 2 раза громче эталонного тона, будет равен 2 сонам, а, если в 2 раза тише – то 0,5 сон.
ЭЛЕМЕНТЫ БИОФИЗИКИ ЗРЕНИЯ
СВЕТОПРЕЛОМЛЯЮЩАЯ СИСТЕМА ГЛАЗА
Органом зрения принято называть глаз. Однако, в зрительную сенсорную систему (зрительный анализатор) вместе с фоторецепторами (палочками и колбочками) и рядом нейронов, которые находятся в сетчатой оболочке глаза входят определенные отделы ЦНС. В то же время, глаз включает не только фоторецепторы и нейроны, но также разнообразные вспомогательные элементы зрительной рецепции. Эти вспомогательные элементы сами не воспринимают свет и не участвуют в переработке (передаче) сигналов от фоторецепторов, но способствуют восприятию света. Важнейшим вспомогательным аппаратом фоторецепции является диоптрическая светопреломляющая система глаза. Упрощенная схема разреза глаза человека имеет вид:
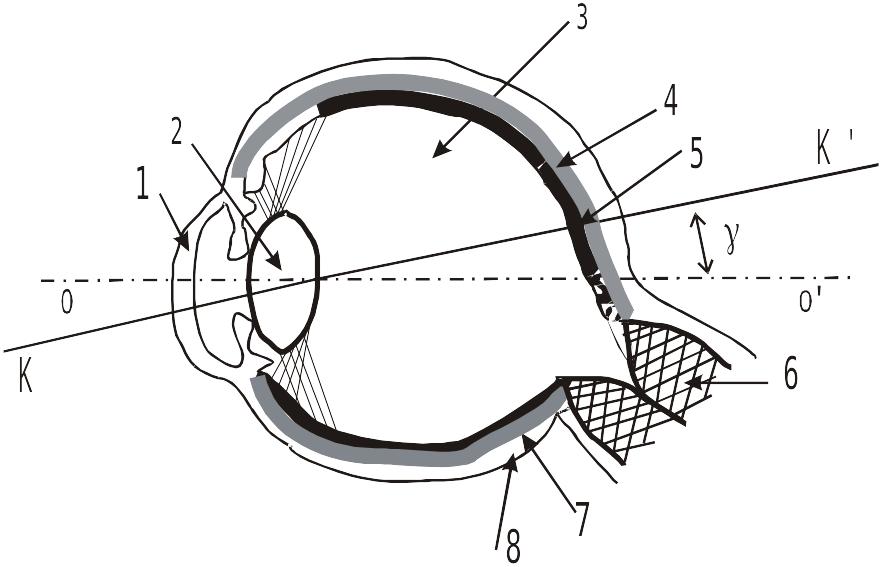
Схема горизонтального разреза глаза человека
1 – роговица; 2 – хрусталик; 3 – стекловидное тело, камерная влага;
4 – сетчатка; 5 – центральная ямка желтого пятна; 6 – зрительный нерв;
7 – сосудистая оболочка; 8 – склера; OO’ – главная оптическая ось;
KK’
-зрительная ось;
![]() -
угол между OO’
и KK’.
-
угол между OO’
и KK’.
Светопреломляющая система глаза фокусирует изображение окружающего мира на сетчатой оболочке (сетчатке), в которой расположены фоторецепторы. Диоптрический аппарат глаза подобен сильной собирающей линзе, и, при рассмотрении далеких предметов, сводит в точку на сетчатке входящие в глаз параллельные лучи, то есть, действует наподобие объектива зрительной трубы. Фокусированные изображения осуществляются за счет преломляющей системы глаза, к которой относятся:
роговица;
хрусталик;
камерная плата;
стекловидное тело.
Преломляя световые лучи, они обеспечивают формирование на сетчатке действительного уменьшенного и обратного (перевернутого) изображения рассматриваемого предмета. Отрезок прямой, проходящий через геометрические центры роговицы и хрусталика, называют главной оптической осью глаза (ОО’). Она объединяет весь его диоптрический аппарат в центрированную светопреломляющую систему. Центрированной называют такую систему светопреломляющих поверхностей, геометрические центры которых лежат на одной прямой. Обратимся к идеальной центрированной системе Гаусса и попытаемся применить ее к человеческому глазу.
Центрированная диоптрическая система при построении изображения предметов рассматривается, как одно целое, и ход лучей в ней определяется при помощи шести, так называемых кардинальных точек, которые лежат на оси ОО’. К этим точкам относятся два главных фокуса (передний F1 и задний F2), две главные точки (Н1 и Н2) и две узловые точки (N1 и N2).
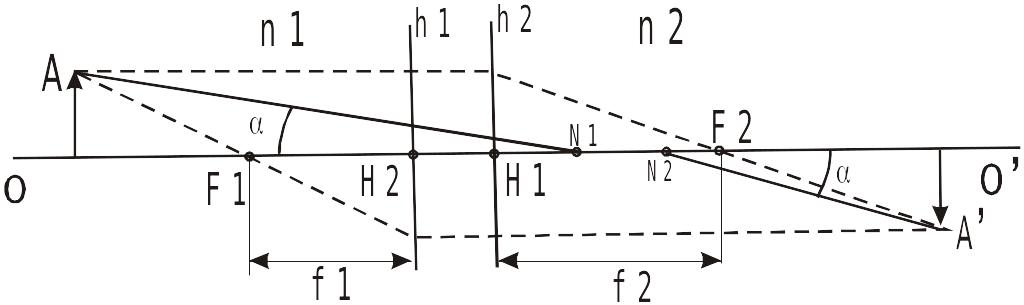
Схема построения оптического изображения
Главными
фокуcами
называют точки (F1
и F2),
в которых после преломления собираются
лучи, входящие в систему параллелей,
так как коэффициенты преломления сред
(n),
находящихся перед глазом
![]() ,
и внутри глаза (n=1,336),
не одинаковы, то и фокусные расстояния
,
и внутри глаза (n=1,336),
не одинаковы, то и фокусные расстояния
![]() .
Размеры изображений предметов,
построенных на h1
и h2,
одинаковы. В местах пересечения главных
плоскостей с главной осью расположены
главные точки Н1 и Н2, от этих точек
отсчитываются фокусные расстояния: от
Н1 – переднее, f1,
от Н2 – заднее f2.
.
Размеры изображений предметов,
построенных на h1
и h2,
одинаковы. В местах пересечения главных
плоскостей с главной осью расположены
главные точки Н1 и Н2, от этих точек
отсчитываются фокусные расстояния: от
Н1 – переднее, f1,
от Н2 – заднее f2.
Среди
лучей, исходящих от предмета, всегда
находится такой, который после преломления
диоптрической системы, покидает ее без
изменения первоначального направления.
Этот луч претерпевает в системе только
смещение, вследствие чего и распространяется
по входу и выходу из нее, параллельно
исходному направлению. Точки, в которых
продолжение падающего луча и выходящего
луча пересекают главную ось, называются
узловыми точками. N1
– передняя узловая точка, N2
– задняя. Через эти точки падающий и
преломленный лучи проходят под одинаковым
углом по отношению к главной оси (![]() ),
применительно к глазу, он называется
углом зрения или зрительным углом.
),
применительно к глазу, он называется
углом зрения или зрительным углом.
ТОНКАЯ И ТОЛСТАЯ ЛИНЗЫ
Если вся центрированная преломляющая система находится в однородной среде (n1 = n2), то N1 совпадает с Н1, а N2 с Н2. Такой относительно простой центрированной диоптрической системой является тонкая линза, когда обе ее поверхности контактируют с воздухом. В тонкой линзе можно пренебречь ее толщиной по сравнению с расстоянием соответствующих фокусов от передней и задней поверхности, тогда, в этом случае, в центрированной системе точки N1 и N2, Н1 и Н2 являются центром оптической линзы, то есть сливаются в одну. При построении изображений, которые получаются в тонкой линзе, окруженной однородной средой, расстояние от оптического центра до предмета и расстояние до изображения от оптического центра, связаны с фокусным расстоянием по следующей формуле:
![]() -
формула тонкой линзы.
-
формула тонкой линзы.
![]() –
расстояние
от предмета до оптического центра;
–
расстояние
от предмета до оптического центра;
b – расстояние от оптического центра до изображения.
Для толстой линзы, чему можно уподобить центрированную систему глаза, формулы, связывающие и b, гораздо сложнее, они включают все кардинальные точки центрированной диоптрической системы. Для упрощения расчетов прибегают к моделям реальных диоптрических систем. При этом стремятся обойтись в модели минимальным количеством кардинальных точек.
ПРИВЕДЕННЫЙ РЕДУЦИРОВАННЫЙ ГЛАЗ
ЧЕЛОВЕКА
Моделью светопреломляющего аппарата органа зрения служит, так называемый, приведенный редуцированный глаз. Редукция означает упрощение, то есть сведение сложного к простому, более доступному для анализа. Физическим аналогом приведенного редуцированного глаза является стеклянная линза, которая одной поверхностью контактирует с воздухом, а другой – с жидкостью, обладающей n = 1,336. Первая поверхность направлена в пространство предметов, а другая поверхность линзы – к пространству изображений.
Диоптрический
аппарат глаза не поддается точному
математическому описанию. Это связано
с тем, что глаза разных людей сильно
различаются, а также из-за подвижности
хрусталика, и по ряду других причин.
Так, например, главная оптическая ось
глаза человека проводится довольно
приблизительно. Она не совпадает со
зрительной осью КК’, которая проходит
через геометрические центры роговицы
и желтого пятна сетчатки. В направлении
зрительной оси глаз имеет наилучшую
разрешающую способность. Угол между
главной оптической осью ОО’ и KK’,
как правило не превышает 5 градусов. Он
обозначается
![]() обычно
учитывается в офтальмологии при
назначении очков.
обычно
учитывается в офтальмологии при
назначении очков.
Итак, человеческий глаз можно рассматривать в качестве центрированной преломляющей системы весьма условно. На рисунке приведена количественная характеристика одной из наиболее адекватных физических моделей человеческого глаза.
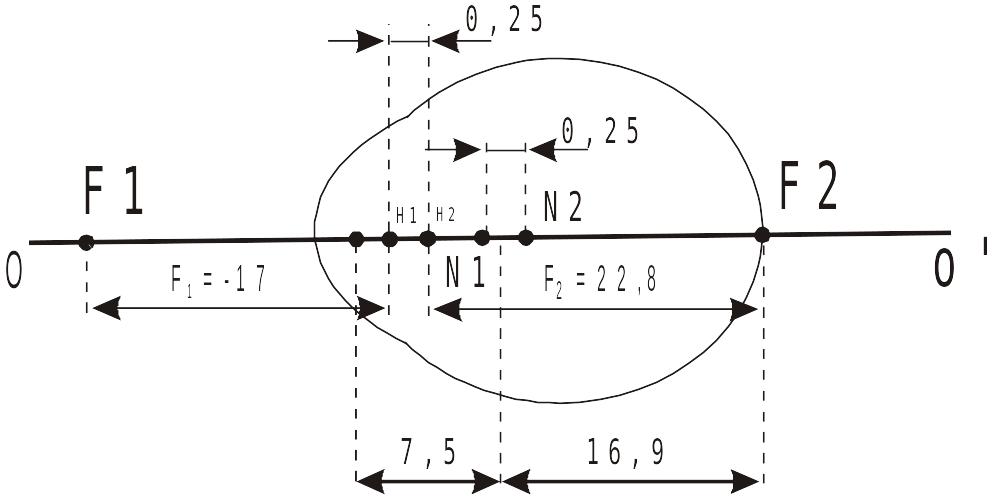
Глаз как центрированная светопреломляющая система
Из рисунка видно, что расстояние между Н1 и Н2 составляет примерно 0,25 мм. Столь близкое расстояние допускает замену двух главных плоскостей (h1 и h2) на одну главную плоскость. Точки N1 и N2 практически также совпадают в единую узловую точку глаза, ее называют оптическим центром глаза (между N1 и N2). В замене каждой из названных пар кардинальных точек на единые угловую и узловую и состоит смысл редукции центрированной диоптрической системы реального глаза. В схеме Гульстранда единая плоскость h и единая узловая точка N находятся на расстоянии 1,6 мм и 7,5 мм соответственно от передней поверхности роговицы. Все расстояния, кроме фокусных (f1, f2), отсчитываются от передней поверхности роговицы. По ходу лучей внутрь глаза эти расстояния имеют знак «+». В противоположном (наружном) направлении расстояния имеют знак «-».
Числа, характеризующие расстояние единых главной и узловой точек глаза, необходимы для расчета диоптрического аппарата человеческого глаза.
БИОФИЗИЧЕСКИЙ МЕХАНИЗМ
ВОСПРИЯТИЯ СВЕТА РЕЦЕПТОРАМИ
Свет, сфокусированный диоптрическим аппаратом глаза на сетчатке, возбуждает ее фоторецепторы. Он достигает фоторецепторов, пройдя всю толщу сетчатки, поскольку палочки и колбочки располагаются в тех слоях сетчатки, которые прилежат ближе к сосудистой оболочке. Свет проходит в сетчатке следующие слои, прежде чем достигнуть фоторецепторов:
Слой ганглеозных клеток.
Внутренний синоптический слой.
Внутренний ядерный слой, состоящий из амакриловых клеток, горизонтальных клеток (Г), биполярных нейронов (Б).
Наружный синоптический слой палочек и колбочек (П и К).
Внутренний ядерный слой П и К.
Каждый глаз человека состоит из примерно 125 млн палочек и примерно 5 млн колбочек.
Фоторецепторы П или К схематично можно представить в виде:
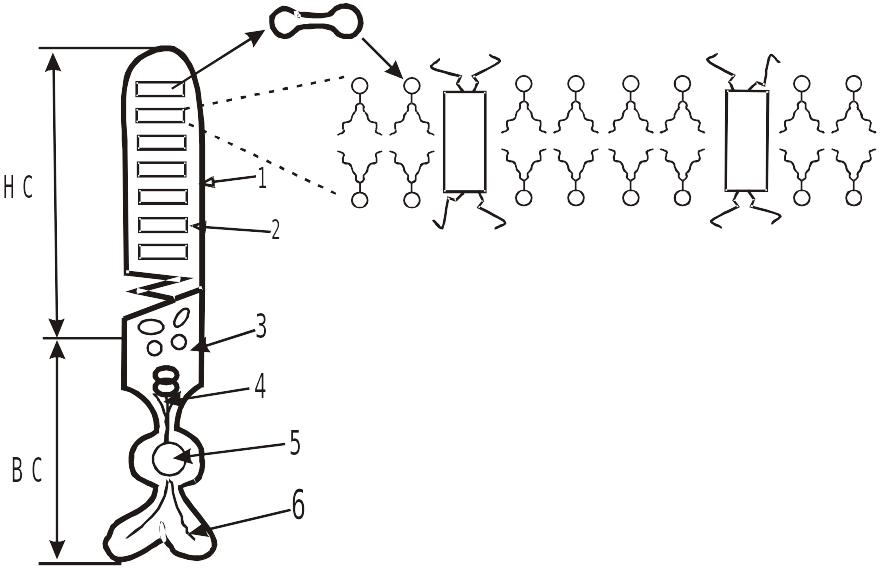
Схема строения фоторецепторной клетки (палочки или колбочки)
1 – наружная мембрана П или К; 2 – диски; 3 – митохондрии; 4 – аппарат Гольджи;
5 – ядро клетки; 6 – синоптическая зона: НС – наружный сегмент;
ВС – внутренний сегмент.
Диаметр П (К) составляет примерно 2 мкм, длина – примерно 50-60 мкм. Длина НС составляет примерно 20-30 мкм. Вся цитоплазма НС заполнена своеобразными органеллами, которые называют дисками. Промежутки между ними не превышают 15 нкм. В НС каждой П (К) содержится от 700 до 1000 дисков, которые напоминают собой сплющенные воздушные шары, сложенные в стопку. Диаметр каждого диска почти соответствует диаметру наружного слоя (примерно 2 мкм), а толщина диска составляет примерно 15-18 нм. Его внутреннее пространство представляет собой узкую щель, шириной = 1 нм. Эта щель расширяется до нескольких нм у краев диска. Таким образом, на внутридисковое содержимое вместе с цитозолем приходится не более половины объема НС, а другую половину занимают мембраны дисков.
НС буквально нафарширован мембранами, и именно в них осуществляется первичный процесс восприятия света. В этой связи, мембрану диска называют фоторецепторной мембраной.
Молекулярная организация фоторецепторной мембраны у позвоночных и беспозвоночных животных практически одинакова. Подобно всем БМ, она состоит из липидов и белков (протеинов), однако, ей присущи две кардинальные отличительные особенности. Во-первых, ее основным белковым компонентом является зрительный пигмент – хромогликопротеид родопсин. Его концентрация достигает 90 % от общего содержания протеинов в фоторецепторной мембране. Во-вторых, более 40 % жирокислотного состава фосфолипидов, образующих дисковые мембраны, приходится на полиненасыщенную декозагексоеновую кислоту, в состав которой входит 22 атома углерода с присутствием 6 двойных связей. Кроме нее, в состав мембранных фосфолипидов входят и другие полиненасыщенные длинноцепочечные жирные кислоты, содержащие от 4 до 6 двойных связей. Вместе они составляют более половины всех жирных кислотных остатков. Высокая степень ненасыщенности жирнокислотных «хвостов» фосфолипидных молекул придает гидрофобному бимолекулярному каркасу фоторецепторной мембраны необычайно высокую подвижность, вследствие чего данная мембрана обладает малой вязкостью по сравнению с другими мембранами.
РЕАКЦИЯ РОДОПСИНА НА ДЕЙСТВИЕ СВЕТА
Первичный механизм возбуждения П светом связан со сложными превращениями родопсина в фоторецепторной мембране. Родопсин – это высокомолекулярное соединение, состоящее из двух основных компонентов:
Альдегид витамина А (ретиналь).
Липопротеин под названием опсин (белок глаза).
Спектр поглощения родопсина имеет вид:
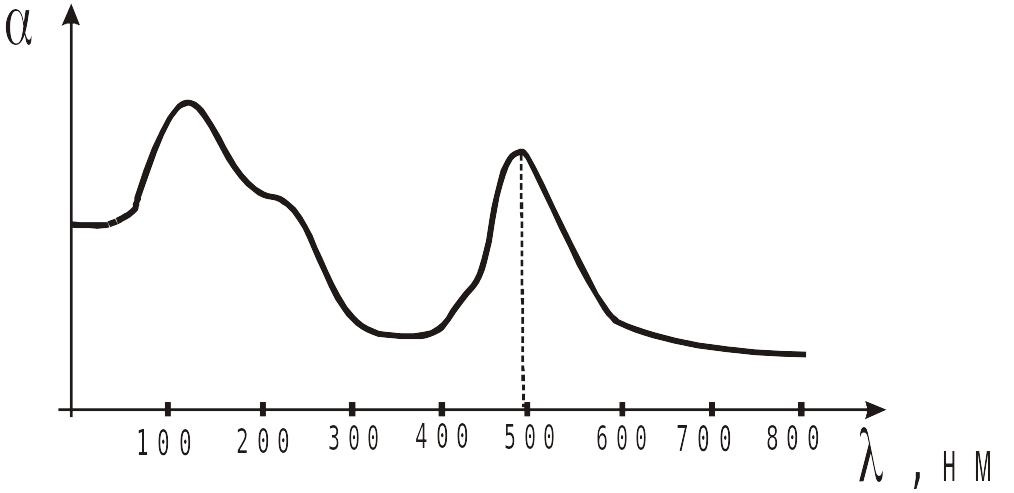
Спектр поглощения родопсина
Спектр поглощения родопсина хорошо приспособлен к восприятию света в достаточно широкой области солнечного излучения, но лучше всего этот зрительный пигмент поглощает свет в зеленой части этого спектра (500 нм). Красные и синие лучи он отражает, причем, в отраженном свете преобладает красный компонент, хотя присутствует и синий. Поэтому пигмент имеет пурпурный цвет, определивший и его название. Родопсин относится к наиболее интенсивно окрашиваемым соединениям из всех известных в настоящее время в органической химии. Возможность поглощения фотонов родопсина близка к теоретически возможному значению.
В темноте ретиналь пребывает в 11 цис-форме, для которой характерна некоторая скрученность молекулы. Скрученность объясняется тем, что в планарной форме между близко расположенными метильной группой у углерода (С13) и водородом у С10, возникает сильное отталкивание. Сила отталкивания не допускает плоского расположения -связи. Скрученность в молекуле 11 цис-ретиналя обеспечивает наибольшую энергию взаимодействия с опсином, вследствие чего их комплекс (ретиналя и опсина) весьма устойчив. Для его разрушения необходимо преобразовать 11 цис-ретиналь в другой изомер, в так называемый полностью транс-ретиналь. Такое преобразование из цис- в транс-ретиналь происходит под действием света. Этот процесс называется фотоизомеризацией.
При
поглощении фотона (![]() ),
молекула родопсина претерпевает
электрический переход «
),
молекула родопсина претерпевает
электрический переход «![]() »,
при этом, молекула родопсина оказывается
в синглетном возбужденном состоянии.
Необходимо отметить, что в органических
молекулах
-орбитали,
которые свойственны основному
энергетическому состоянию, являются
связывающими, то есть, они стремятся к
перекрытию и образованию второй связи.
Напротив,
»,
при этом, молекула родопсина оказывается
в синглетном возбужденном состоянии.
Необходимо отметить, что в органических
молекулах
-орбитали,
которые свойственны основному
энергетическому состоянию, являются
связывающими, то есть, они стремятся к
перекрытию и образованию второй связи.
Напротив,
![]() -орбитали, характерные для возбужденного
состояния молекулы, относятся к
разрыхляющим орбиталям. Их перекрытие
энергетически невыгодно. В силу чего,
части молекулы, соединенные двойной
связью с участием
-орбитали,
разворачиваются на 90 градусов, и двойная
связь превращается в одинарную. В этой
связи, в молекуле ретиналя, возбужденной
светом, происходит поворот атомов
вокруг двойной связи. Такое измененние
пространственного расположения атомов,
входящих в состав ретиналя, требует
затрат энергии. Эти энергозатраты
характеризуются преодолением
потенциального барьера. Для разворота
атома вокруг двойной связи в 11 цис-ретинале
достаточно энергии одного фотона. Для
фотоизомеризации достаточно и времени,
в течение которого молекула родопсина
пребывает в сигнетном возбужденном
состоянии (
-орбитали, характерные для возбужденного
состояния молекулы, относятся к
разрыхляющим орбиталям. Их перекрытие
энергетически невыгодно. В силу чего,
части молекулы, соединенные двойной
связью с участием
-орбитали,
разворачиваются на 90 градусов, и двойная
связь превращается в одинарную. В этой
связи, в молекуле ретиналя, возбужденной
светом, происходит поворот атомов
вокруг двойной связи. Такое измененние
пространственного расположения атомов,
входящих в состав ретиналя, требует
затрат энергии. Эти энергозатраты
характеризуются преодолением
потенциального барьера. Для разворота
атома вокруг двойной связи в 11 цис-ретинале
достаточно энергии одного фотона. Для
фотоизомеризации достаточно и времени,
в течение которого молекула родопсина
пребывает в сигнетном возбужденном
состоянии (![]() с),
поскольку цис-тран-изомеризация ретиналя
происходит всего за 0,1-0,3 пс, квантовый
выход фотоизомеризации 11 цис-ретиналя
в родопсине достигает 0,7.
с),
поскольку цис-тран-изомеризация ретиналя
происходит всего за 0,1-0,3 пс, квантовый
выход фотоизомеризации 11 цис-ретиналя
в родопсине достигает 0,7.
Фотоизомеризация – есть первая и единственная фотохимическая реакция в зрительном акте. Она протекает внутри отдельной молекулы родопсина и, в отличие от фотосинтеза, не сопровождается никакими химическими реакциями. Зрение отличается от фотосинтеза, который происходит в растениях, еще и тем, что между молекулами родопсина невозможен перенос энергии, так как, в фоторецепторной мембране молекулы родопсина удалены друг от друга на расстояние более 7 нм. Непосредственным следствием фотоизомеризации, приводящей к появлению у человека ощущения света, служат конформационные перестройки молекулы родопсина. Дело в том, что пространственно 11 цис-ретиналь точно соответствует конфигурации активного центра. При образовании транс-ретиналя, это соответствие (комплементарность) нарушается. В результате чего ретиналь отщепляется от опсина, и родопсин обесцвечивается, то есть, претерпевает фотолиз.
Вне
действия света, распавшийся родопсин
восстанавливается. В ходе восстановления,
транс-ретиналь преобразуется в цис-форму.
Такой переход обратной фотоизомеризации
называется реизомеризацией. У позвоночных
животных транс-ретиналь, отщепившись
от опсина, поступает в пигментный
эпителий сетчатки, где подвергается
ферментативным превращениям. Из
пигментного эпителия фоторецептором
(П, К) доставляется 11 цис-ретиналь. Каждая
палочка ежесекундно синтезирует
![]() молекул родопсина из поступающего в
нее 11 цис-ретиналя, который соединяется
с опсином. Суточный синтез этого
зрительного пигмента в целом оценивается
величиной примерно
молекул родопсина из поступающего в
нее 11 цис-ретиналя, который соединяется
с опсином. Суточный синтез этого
зрительного пигмента в целом оценивается
величиной примерно
![]() молекул.
молекул.
Конформационная перестройка родопсина, называющаяся фотоизомеризацией ретиналя, довольно существенна. Естественно, что перестройку сопровождают значительные изменения физико-химических свойств фоторецепторной мембраны, в которую встроен родопсин. Одним из важных последствий этих изменений служит сдвиг мембранного потенциала фоторецепторной клетки.
РЕЦЕПТОРНЫЕ ПОТЕНЦИАЛЫ
Сдвиг мембранного потенциала фоторецепторной клетки (П или К), который принято называть рецепторным потенциалом, происходит на ее плазмолемме, тогда, как фотоизомеризация родопсина совершается преимущественно во внутриклеточных мембранах, то есть, в мембранных дисках наружных сегментов.
Свойство
фоторецепторных мембран изменяется
одновременно с конформационной
перестройкой в них опсина. После
поглощения
(кванта света), спустя 1,5-2 мс, наблюдается
гиперполяризация плазмолеммы
фоторецепторной клетки (П или К).
Следовательно, в палочке (К) существует
механизм передачи сигнала с фоторецепторных
мембран дисков на плазмолемму П (К). В
передаче участвует, так называемый,
внутриклеточный посредник, природа
которого до настоящего времени полностью
не установлена, однако, считают, что в
посредничестве участвуют циклический
гуанозинмонофосфат (цГМФ) и ионы
![]() .
.
В следствие конформационной перестройки молекул родопсина, в фоторецепторной мембране изменяется содержание внутриклеточного посредника в цитозоле наружного сегмента П или К. Это приводит к закрытию 100-300 натриевых каналов в плазмолемме, которые в отсутствие светового раздражителя пребывали в открытом состоянии. Они обладают свойствами потенциалнезависимых ионных каналов и, будучи управляемые светом, получили название фотозависимых натриевых каналов. Когда закрываются 100-300 каналов, электропроводимость плазмолеммы наружного сегмента падает на 1-3%. Падение мембранной электропроводимости приводит в гиперполяризации фоторецепторной клетки. Гиперполяризационный сдвиг мембранного потенциала П (К), возникающий под действием света, называют рецепторным потенциалом (РП). Принято считать, что ПП фоторецепторной клетки генерируется в темноте на плазмолемме внутреннего сегмента. По своей природе ПП является калиевым потенциалом, подобно всякому мембранному потенциалу в условиях покоя. В темноте ПП палочки (колбочки) составляет примерно от –20 до –40 мВ.
В
системе генерации энергетического
потенциала П плазмолемма наружного
сегмента выполняет роль шунта с
переменной электропроводностью.
Электропроводность регулируется
(управляется) светом при помощи
фотозависимых натриевых каналов. Когда
все натриевые каналы плазматической
мембраны наружного сегмента одной П
оказываются заблокированными светом,
удельное поверхностное сопротивление
этой мембраны достигает величины
![]() .
В темноте ионный ток, входящий через
каналы в цитоплазму, довольно значителен,
и составляет примерно 40 nА.
А при освещении сетчатки он блокируется,
при этом степень его ослабления зависит
от интенсивности света. Чем слабее
натриевый ток через плазматическую
мембрану наружного сегмента, тем больше
выражена гиперполяризация в фоторецепторной
клетке. Изменение потенциала П (К) во
времени имеет вид:
.
В темноте ионный ток, входящий через
каналы в цитоплазму, довольно значителен,
и составляет примерно 40 nА.
А при освещении сетчатки он блокируется,
при этом степень его ослабления зависит
от интенсивности света. Чем слабее
натриевый ток через плазматическую
мембрану наружного сегмента, тем больше
выражена гиперполяризация в фоторецепторной
клетке. Изменение потенциала П (К) во
времени имеет вид:
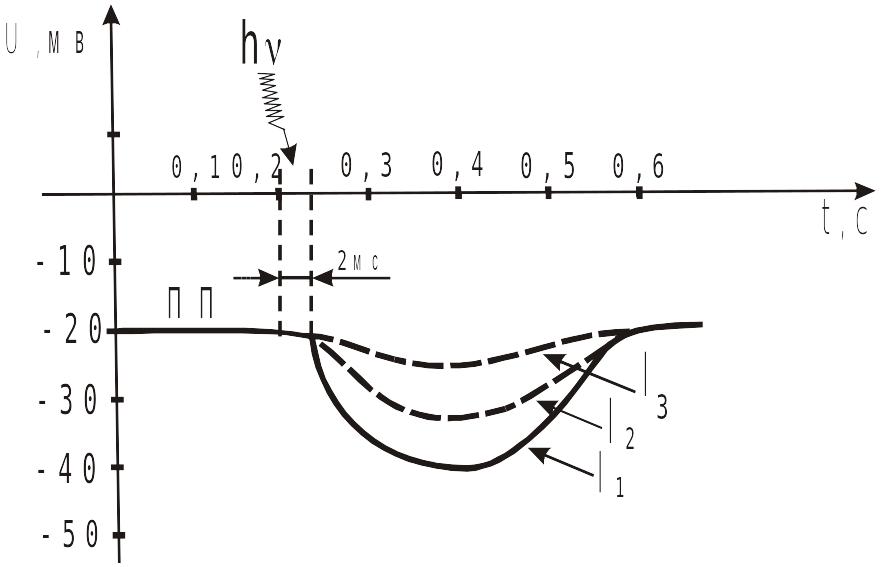
Рецепторные потенциалы одиночной колбочки, возникающие в ответ
на короткие (10 мс) вспышки света при трех разных интенсивностях света
![]() –
интенсивность
света. Чем больше I,
тем больше гиперполяризация
–
интенсивность
света. Чем больше I,
тем больше гиперполяризация
![]() .
.
При
поглощении 1 фотона (
)
гиперполяризация фоторецепторов разных
представителей животного мира
неодинакова. Так, в палочках сетчатки
позвоночных животных гиперполяризация
составляет десятки и сотни мкВ, тогда,
как в фоторецепторных клетках
членистоногих она достигает 10 мВ. Эти
приведенные числа подтверждают положение
об усилении сигнала в рецепторных
аппаратах. Действительно, квант света,
несущий энергию
![]() Дж,
вызывает фотолиз только одной молекулы
родопсина, что в свою очередь приводит
к закрытию в плазмолемме фоторецепторной
клетки позвоночных животных 100-300
натриевых каналов. Это приводит к
образованию РП. Однако, энергия РП на
3 порядка превосходит энергию вызвавшего
его фотона.
Дж,
вызывает фотолиз только одной молекулы
родопсина, что в свою очередь приводит
к закрытию в плазмолемме фоторецепторной
клетки позвоночных животных 100-300
натриевых каналов. Это приводит к
образованию РП. Однако, энергия РП на
3 порядка превосходит энергию вызвавшего
его фотона.
Амплитуда рецепторного потенциала возрастает при повышении интенсивности света, падающего на сетчатку, причем, в пределах 1000-кратного изменения интенсивности, эта зависимость имеет логарифмический характер:
![]() -
изменение амплитуды РП.
-
изменение амплитуды РП.
- коэффициент пропорциональности;
![]() -
интенсивность последующего;
-
интенсивность последующего;
- интенсивность предыдущего.
При ярком освещении амплитуда рецепторного потенциала может достигать 25 мВ. Величина РП зависит также и от длины волны света. П или К в ответ на зеленый свет ( =500 нм), генерируют наибольший РП. Среди К выделено 3 типа:
дает максимальный ответ РП на синий цвет;
дает максимальный ответ РП на зеленый цвет;
дает максимальный ответ РП на желто-красный цвет.
Использование микроэлектронной техники для изучения реакции фоторецепторов на облучение светом, позволило обнаружить деполяризацию П и К, при этом деполяризация предшествует гиперполяризации. Амплитуда деполяризации очень мала и составляет примерно 0,1 мкВ. Учитывая временные соотношения де- и гиперполяризации, первую из них стали называть ранним, а вторую – поздним РП, и обозначили, соответственно, РРП и ПРП. Установлено, что РРП обусловлен преимущественно перемещениями электрических диполей в мембранах наружного сегмента. Эти перемещения диполей связаны с фотолизом молекул родопсина. Другими словами, РРП обусловлен током смещения в фоторецепторных мембранах, и РРП не служит промежуточным звеном передачи сигналов от них к синоптической зоне П или К, при действии света. Эта роль принадлежит гиперполяризации. В отличие от других рецепторных аппаратов, где таким звеном служит деполяризационные сдвиги мембранного потенциала. Таким образом, только поздний РП является рецепторным потенциалом П или К в полном значении этого термина.
РАСПРЕДЕЛЕНИЕ СИГНАЛА ПО СЕТЧАТКЕ
Поздний РП, возникнув в НС, распределяется в синоптической зоне П или К по закону электротонической передачи (с декрементом). При этом, постоянная длины плазмолеммы фоторецепторной клетки, которая проводит гиперполяризацию, достигает 10 мкм. Достигнув синоптической зоны П (К) ПРП изменяет свойства ее мембраны и, тем самым, влияет на медиаторную систему, то есть, на передачу сигнала к биполярному нейрону. Химическая природа медиатора, выделяемого фоторецепторной клеткой, пока еще неизвестна точно, однако, наиболее вероятными кандидатами на эту роль признаются глутамат и аспарат. Обе аминокислоты обладают деполяризующим действием на постсинаптическую мембрану. При облучении П (К) светом, биполярные нейроны так же испытывают гипреполяризацию. Принято считать, что фоторецепторной клетке в темноте свойственен определенный темп выделения деполяризующего медиатора (глутамата или аспарата), а гиперполяризация синаптической зоны при освещении замедляет темп выброса медиатора, следовательно, постсиноптические мембраны, принадлежащие биполярным нейронам, также гиперполяризуются. Гиперполяризационный сдвиг мембранного потенциала биполярного нейрона электротонически распространяется по эго-плазмолемме к синапсу на теле третьего нейрона, который входит в состав афферентного пути сетчатки. Медиатор, поступающий от биполярного нейрона ганглиозной клетки, вызывает ее деполяризацию. Деполяризационный сдвиг потенциала распределяется электротонически с тела по аксонам ганглиозной клетки. Плазматическая мембрана аксона ганглиозной клетки (аксолемма) обладает возбудимостью, благодаря чему там и формируется ПД, когда деполяризация достигает порогового уровня. ПД, неся информацию от фоторецепторов, поступает в ЦНС по волокнам зрительного нерва. Эти волокна представляют собой аксоны ганглиозных клеток сетчатки. Схематически передачу поздних РП по сетчатке можно представить в виде:
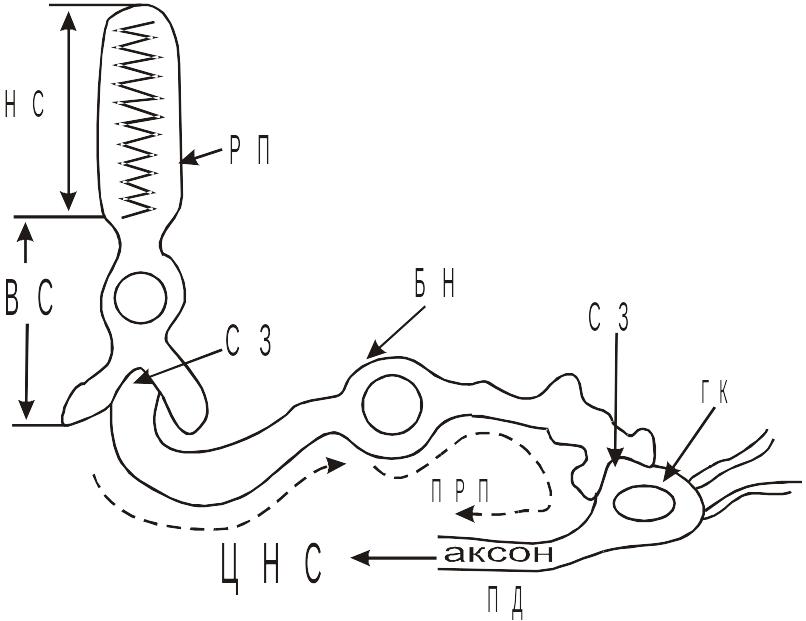
Схема передачи фоторецепторного сигнала по афферентному пути в пределах сетчатки
РП – рецепторная палочка; НС – наружный сегмент; ВС – внутренний сегмент; СЗ – синоптическая зона; ГК – ганглиозная клетка; ПРП – поздний рецепторный потенциал; ПД – потенциал действия; ЦНС – центральная нервная система
Зрительный
нерв включает примерно
![]() волокон.
По этим волокнам в мозг направляются
сигналы от 130
волокон.
По этим волокнам в мозг направляются
сигналы от 130![]() фоторецепторов. Следовательно, в среднем
1 аксон ГК собирает информацию от
множества рецепторных клеток. Все
фоторецепторы, посылающие сигналы к
одной ГК, составляют ее рецепторное
поле. Рецепторные поля, образованные
П, гораздо многочисленнее, по сравнению
с К рецепторными полями. Необходимо
отметить, что каждая К, находящаяся в
центральной ямке желтого пятна сетчатки,
связана только с одной ГК. Другими
словами, рецепторное поле нейрона
включает только одну колбочку. С этим
связана наилучшая разрешающая способность
центрального К зрения. Другие ганглиозные
клетки получают сигналы больше, чем от
1 фоторецептора. Существуют такие ГК,
которые собирают сигнализацию от
множества тысяч палочек, следовательно,
рецепторные поля сетчатки весьма
разнообразны.
фоторецепторов. Следовательно, в среднем
1 аксон ГК собирает информацию от
множества рецепторных клеток. Все
фоторецепторы, посылающие сигналы к
одной ГК, составляют ее рецепторное
поле. Рецепторные поля, образованные
П, гораздо многочисленнее, по сравнению
с К рецепторными полями. Необходимо
отметить, что каждая К, находящаяся в
центральной ямке желтого пятна сетчатки,
связана только с одной ГК. Другими
словами, рецепторное поле нейрона
включает только одну колбочку. С этим
связана наилучшая разрешающая способность
центрального К зрения. Другие ганглиозные
клетки получают сигналы больше, чем от
1 фоторецептора. Существуют такие ГК,
которые собирают сигнализацию от
множества тысяч палочек, следовательно,
рецепторные поля сетчатки весьма
разнообразны.
ОСНОВЫ ФОТОМЕТРИИ
Свойства ФР и всей зрительной системы человека оказали весьма большое влияние на фотометрию. Фотометрия – это раздел физической оптики, в котором рассматриваются энергетические характеристики светового излучения в процессах его испускания, распределения и взаимодействия с веществом. К области фотометрии относится только та часть электромагнитного спектра, которая воспринимается человеческим глазом. В необъятном спектре электромагнитных колебаний, только узкая полоса видимой области электромагнитных колебаний с длиной волн от 380 нм до 760 нм, воспринимается зрительной системой человеческого глаза, то есть, как принято говорить, обладает видимостью. Однако, к волнам разной длины, принадлежащим к видимой области, электромагнитная колебательная чувствительность зрительного анализатора человека неодинакова, следовательно, различаемые участки видимой области спектра, обладают разной видимостью. Кривые видности были построены в 20-е годы ХХ века по результатам определения чувствительности зрительного анализатора человека к свету разных длин волн. Кривые видности имеют вид:
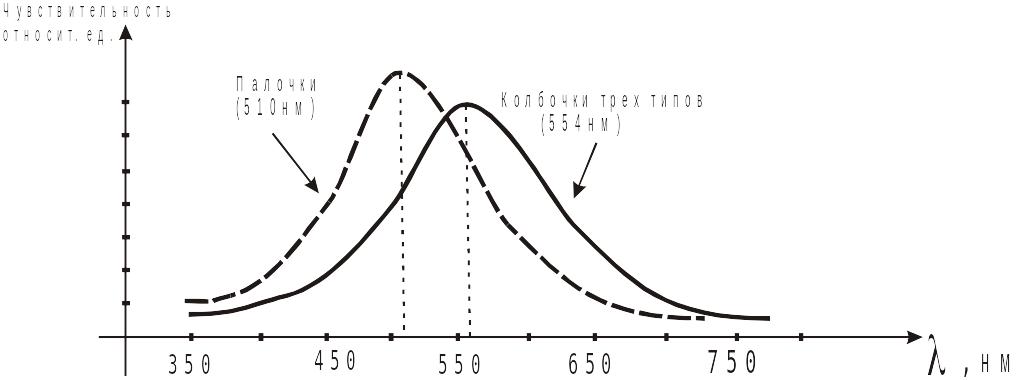
Кривые видности фотопического (сплошная линия) и скотопического
(штриховая линия) зрения
Одна из кривых построена для скотопического (палочкового) зрения (штрихованная линия). Она почти полностью совпадает со спектром поглощения родопсина в видимой области спектра. Максимальная чувствительность зрительного восприятия, когда она обеспечивается П при малой освещенности примерно 0,01 люкса (Лк) приходится на =510 нм (зеленый свет). Другая кривая видимости отображает спектральную чувствительность фотопического (колбочкового) зрения. Фотопическая кривая является результатом интегрирования функции спектральной чувствительности К трех типов. Интегрирование осуществляется в центральном отделе зрительного анализатора. Максимальная интегральная световая чувствительность К зрительной системы приходится на =554 нм.
Чувствительность
ФР необычно высока, так, возбуждение П
(К) происходит при поглощении его 1
кванта света. Абсолютная световая
чувствительность зрительной системы
человека определяется по порогу ощущения
ниже чувствительности отдельного
рецептора. Человек ощущает только такой
слабый свет, который одновременно
возбуждает не менее 5-10 палочек, для
чего им необходимо поглотить столько
же фотонов. В ходе биологической
эволюции, П сетчатки достигли предела
световой чувствительности, который
допускается квантовой теорией. Этим
обусловлена высочайшая чувствительность
к свету всей зрительной системы человека.
Так, пороговая яркость для стахопического
зрения составляет
![]() кд/
.
кд/
.
СОПОСТАВЛЕНИЕ ЭНЕРГЕТИЧЕСКИХ
И СВЕТОТЕХНИЧЕСКИХ ПАРАМЕТРОВ СВЕТА
Кривая видности фотопического зрения позволяет осуществить приведение физических величин, характеризующих световую энергию, к, так называемым, светотехническим величинам, которые выражают ее оценку с учетом особенностей зрительного восприятия человека. Две группы названных параметром сведем в таблицу:
N |
Энергетические параметры света |
N |
Психологические параметры |
1 |
Поток излучения (лучистый поток)
|
1 |
Световой
поток
|
2 |
Энергетическая
сила света
|
2 |
Сила света I [кд] |
3 |
Энергетическая
яркость
|
3 |
Яркость
|
4 |
Энергетическая
освещенность (облученность)
|
4 |
Освещенность
|
Основным техническим параметром света служит поток излучения – это энергия квантов света, излучаемая в единицу времени.
Энергетическая сила света представляет собой поток излучения, приходящийся на единицу телесного угла в данном направлении.
Под энергетической яркостью понимают энергетическую силу света, отнесенную к единице площади проекции поверхности излучающего тела на направление, перпендикулярное распределению света.
Энергетическая освещенность – это поток излучения, падающий на единицу площади облучаемой поверхности. Следовательно, энергетическая яркость является характеристикой источника излучения, а энергетическая освещенность характеризует облучаемую поверхность.
За
основную светотехническую величину
принята сила света. Ее размерность –
[кд] Значение [кд] принимают таким, что
яркость полного излучения при температуре
затвердевания Pt
(2042 К или
![]() С) равна 60 kд/
С) равна 60 kд/![]() .
Применявшаяся ранее международная
свеча составляет 1,005 kд.
Производными параметрами от силы света
являются яркость и световой поток.
Единицей яркости служит kд/
,
то есть, одна kд/
характеризует яркость источника каждого
излучающей поверхности, которая имеет
в данном направлении силу света = 1 kд.
В медицине широко используются и
внесистемные единицы яркости света:
нит, стильб (сб), апостильб (апб), ламберд
(Лб).
.
Применявшаяся ранее международная
свеча составляет 1,005 kд.
Производными параметрами от силы света
являются яркость и световой поток.
Единицей яркости служит kд/
,
то есть, одна kд/
характеризует яркость источника каждого
излучающей поверхности, которая имеет
в данном направлении силу света = 1 kд.
В медицине широко используются и
внесистемные единицы яркости света:
нит, стильб (сб), апостильб (апб), ламберд
(Лб).
![]() ;
;
![]() ;
;
![]()
Яркость
снега в солнечный день достигает
примерно
![]() kд/
,
а яркость земляного грунта беззвездной
ночью – примерно
kд/
,
а яркость земляного грунта беззвездной
ночью – примерно
![]() kд/
.
kд/
.
Световой поток определяется произведением силы света на телесный угол, в который он распределяется.
Производной величиной от светового потока является освещенность, представляющая собой световой поток, приходящийся на единицу облучаемой поверхности. Соотношение между яркостью объекта и освещенностью выражается формулой:
![]()
– коэффициент отражения света;
В – яркость;
E – освещенность.
ПРИВЕДЕНИЕ ЭНЕРГЕТИЧЕСКИХ ПАРАМЕТРОВ К СВЕТОТЕХНИЧЕСКИМ
Приведение осуществляется посредством величины, которая получила название видности. Абсолютной видностью – v – называют соотношение светового потока к тому энергетическому потоку излучения, который вызывает ощущение данного потока.
![]()
Размерность [v]=[лм/Вт], и отражает субъективную «стоимость» в лм зрительного ощущения, вызываемого потока излучения в 1 Вт. Известно, что 1 Вт излучения разных вызывает неодинаковые по интенсивности ощущения света. Самым ярким ощущается свет с = 554 нм. На эту приходится максимум кривой видности фотопического зрения. Поток излучения в 1 Вт, при = 554 нм, создает у человека ощущение светового потока в 683 лм, следовательно, максимальная абсолютная видность равна 683 лм/Вт.
Для
излучения на другую
,
абсолютная видность ниже, что наглядно
выражается кривой видностью. Однако,
при решении фотометрических задач,
удобно использовать относительную
видность (![]() ),
под которой понимают отношение абсолютной
видности при данной
к максимуму видности.
),
под которой понимают отношение абсолютной
видности при данной
к максимуму видности.
![]()
Соответственно, что относительная видность равна 1 при =554 нм. При всех других , относительная видность меньше 1.
БИОФИЗИЧЕСКИЕ ОСНОВЫ ВКУСА
ВОСПРИЯТИЕ ВКУСА
В эволюции животных огромное значение имело контактное дифференцированное распознавание пищи, в результате которой происходило или поедание пищи или отклонение ее.
Химические соединения, входящие в состав пищи животных, действуя на вкусовые рецепторы, обусловливают ее соленый, кислый, сладкий или горький вкус. Имеются более сложные системы квалификации вкусовых ощущений, однако, для человека предполагают справедливым предположение о наличии только четырех элементарных вкусовых ощущений.
У человека вкусовые луковицы, содержащие вкусовые рецепторы, находятся во вкусовых сосочках, которые расположены на поверхности языка, на задней стенке глотки, в мягком небе, в миндалинах и в надгортаннике. Вкусовые луковицы имеют овальную форму и содержат, кроме вкусовых клеток, также и опорные клетки. Вкусовые клетки сообщаются с поверхностью слизистой оболочки с помощью вкусовых пор.
Рецепторные клетки в опекальной области имеют микровиллы (микроворсинки), которые в виде кисточки через вкусовые поры сообщаются с жидкой средой слизистой оболочки. Предполагают, что на поверхности микровилл расположены активные белковые центры, с которыми реагируют молекулы или ионы вкусовых веществ. Пространство между микровиллами заполнено особой субстанцией мукополисахаридной и мукопротеидной природы. Некоторые ученые считают, что эта субстанция функционирует наподобие ионно-обменной смолы, давая возможность молекулам или ионам одних веществ контактировать с поверхностью микровилл, а других – нет. У основания луковицы вкусовые клетки образуют синопсы с окончаниями афферентных нервных волокон лицевого языкоглоточного и блуждающего нервов.
В последние десятилетия были проведены большие исследования вкусового анализатора с помощью микро-ЭВМ. Было установлено, что вкусовые клетки имеют потенциал покоя ПП величиной, которая изменяется в пределах от –30 до –50 мВ. При действии вкусового стимула на клетку, наблюдается медленная деполяризация клеточных мембран, то есть, формируется генераторный потенциал ГП. В зависимости от интенсивности стимула, его величина обычно меняется в пределах от 15 до 45 мВ.
Механизм передачи возбуждения от вкусовой клетки к нервному волокну окончательно не выяснен. Однако, известно, что передача в синапсе осуществляется при участии ацетилхолина.
Оказалось, что одна и та же клетка может реагировать на сладкие, соленые и кислые стимулы тогда, как другие клетки обладают повышенной чувствительностью только к одному или двум стимулам. В отличие от вкусовых клеток, вкусовые сосочки строго специфичны, при этом, электрическое раздражение отдельных сосочков вызывает, как правило, одно из четырех вкусовых ощущений. Химическая стимуляция этих же сосочков у человека вызывает аналогичное ощущение. На основе подобных опытов, Бекеши в 1966 г. пришел к заключению, что существует полное совпадение вкусовых качеств для каждого отдельного сосочка, стимулируемого как электрическим, так и химическим путем. Сосочки отличаются строгой локализацией на поверхности языка, и делятся на соленые, кислые, сладкие и горькие. Возможно, что строгая специфичность сосочков обусловлена ионно-обменными свойствами субстрации, которая заполняет пространство между микровиллами. При этом, субстрация обусловливает избирательную проницаемость вкусовых веществ.
В опытах с отведением потенциалов от афферентных нервов, было обнаружено, что небольшое количество нервных волокон обладает узкой вкусовой специфичностью, а большая часть волокон обладает или относительной специфичностью или вообще, не обладает специфичностью. Необходимо отметить, что наличие специфичности в периферическом отделе вкусового анализатора, обусловливает формирование дифференцированных вкусовых ощущений в центральном отделе головного мозга. Натуральные пищевые раздражение одновременно возбуждает несколько или все вкусо-воспринимающие элементы. В результате чего возникает сложное вкусовое ощущение.
ХИМИЧЕСКИЕ СВОЙСТВА ВКУСОВЫХ ВЕЩЕСТВ И ТЕОРИЯ ВКУСА
Все вкусовые вещества условно можно разделить на три группы:
Вещества, близкие по своей химической структуре и вызывающие примерно одинаковые вкусовые ощущения.
Вещества, близкие по химическим свойствам, но обладающие разным вкусом.
Вещества разной структуры, но сходного вкуса.
К первой группе относятся сахара: глюкоза, сахароза, галактоза, лактоза. Все они обладают сладким вкусом. Кислым вкусом обладают диссоциированные неорганические и органические кислоты.
Ко второй группе относится ряд изомеров, например, -валин, имеющий горьковато-сладкий вкус; 1-валин, имеющий сладкий вкус; d-лейцин, имеющий сладкий вкус; 1-лейцин, имеющий горький вкус.
К третьей группе можно отнести, например, полисахариды: глицерин, гликокол, сахарин, нитробензол, хлористый метил, хлороформ, - имеющий сладкий вкус, а так же, хинин, нитропроизводные бензола и многие неорганические соли и кислоты, имеющий горький вкус. Аналогичность вкуса при таком несходстве химической структуры вещества, главным образом, относится к горькому и сладкому вкусам.
Соленый
вкус NaCl
присущ и другим хлоридам (K,
![]() ,
Ca
и т.д.). Можно было бы предположить, что
свободный анион
,
Ca
и т.д.). Можно было бы предположить, что
свободный анион
![]() ,
а не катионы
,
,
,
а не катионы
,
,
![]() ,
,
ответственен за соленый вкус, однако,
значение катионов также нельзя исключить.
Так, например, в слабой концентрации
NaCl
обладает соленым вкусом, NaBr
– горьковато-соленым, NaI
– солоновато-горьким. Из этого можно
заключить, что ион
придает растворам всех этих солей
присущий им солоноватый вкус. В настоящее
время считают, что в основе соленого
стимула лежит действие низкомолекулярных
анионов и катионов. При этом, принимают,
что интенсивность вкуса низкомолекулярных
солей зависит от катионов (
,
,
,
,
ответственен за соленый вкус, однако,
значение катионов также нельзя исключить.
Так, например, в слабой концентрации
NaCl
обладает соленым вкусом, NaBr
– горьковато-соленым, NaI
– солоновато-горьким. Из этого можно
заключить, что ион
придает растворам всех этих солей
присущий им солоноватый вкус. В настоящее
время считают, что в основе соленого
стимула лежит действие низкомолекулярных
анионов и катионов. При этом, принимают,
что интенсивность вкуса низкомолекулярных
солей зависит от катионов (
,
,
![]() ,
,
![]() и т.д.), а характер вкуса от анионов (
,
и т.д.), а характер вкуса от анионов (
,
![]() и т.д.).
и т.д.).
Кислым вкусом обладает большинство органических и неорганических кислот. Как известно, общей чертой всех кислот является их способность к диссоциации в водных растворах на анионы и катионы . Кислый стимул связывают с воздействием на вкусовые клетки ионов . В этом убедились при помощи опытов. Так, например, HCl ощущается кислотой при разбавлении в пропорции 1:800, в то время, как при этой же концентрации, весь диссоциированный NaCl безвкусен. Воздействие ионов пропорционально его концентрации, однако, более сильные кислоты в одинаковой концентрации со слабыми, не всегда вызывают более сильное вкусовое ощущение. Это объясняется тем, что для ионов проницаемость субстанции окружающей микровиллы вкусовых клеток имеет низкое значение. Слабые же кислоты в виде недиссоциированных молекул, достигают микровилл вкусовых клеток, и только затем диссоциируют. При этом, образовавшиеся ионы взаимодействуют с активными центрами микровилл.
Сладкий
вкус вызывается большим числом
органических веществ, которые, как
правило, не диссоциируют. В настоящее
время окончательно не выяснено, какие
свойства молекул вызывают сладкий
вкус. Многие сладкие вещества являются
многоатомными спиртами с повторяющейся
группой
![]() (сахара,
глицерин). Сахарин, который обладает
сладким вкусом, имеет совершенно другую
структуру. По мнению Акри, сладкий
стимул молекул связан с их возможностью
образовывать слабые водородные связи,
с активным центром рецепторной молекулы,
которая локализована во вкусовой
клетке. Во всех случаях, структура
молекулы должна соответствовать
стерическому положению, и возможностям
водородных связей рецепторной молекулы.
(сахара,
глицерин). Сахарин, который обладает
сладким вкусом, имеет совершенно другую
структуру. По мнению Акри, сладкий
стимул молекул связан с их возможностью
образовывать слабые водородные связи,
с активным центром рецепторной молекулы,
которая локализована во вкусовой
клетке. Во всех случаях, структура
молекулы должна соответствовать
стерическому положению, и возможностям
водородных связей рецепторной молекулы.
Менее всего изучена природа горького стимула вкусовых веществ. Было отмечено, что соли, по мере увеличения молекулярного веса, становятся все более горькими. Так, например, раствор NaCl имеет соленый вкус, цезий-хлор – горький, KI – горький. Имеются данные, что горький стимул связан с возможностью образования слабых дисперсионных связей типа водородных.
Существует несколько теорий, которые пытаются объяснить конкретные процессы взаимодействия молекул вкусовых веществ с рецепторными клетками. Так, Лазарев еще в 1920 г., исходя из роли ионов в процессе возбуждения, допускал, что вкусовые луковицы каждого сосочка содержат высокочувствительные вещества белковой природы, которые разлагаются под влиянием адекватного стимула. При этом ионизируемые продукты распада возбуждают соответствующие нервные окончания.
В настоящее время наиболее разработанной является теория Бейдлера. Он предположил, что вкусовые стимулы (молекулы или ионы) взаимодействуют с определенными участками плазматической мембраны вкусовой клетки, которые он выделяет в качестве активных центров. Активные центры представляют собой полиэлектролиты белковой природы, содержащие большое количество заряженных боковых цепей. Взаимодействие вкусовых стимулов с активными центрами протекает по типу мономолекулярной реакции, которая имеет вид:
С+(N-Z)![]() (1)
(1)
С – концентрация стимулирующего вкусового вещества;
N – общее число активных центров рецепторов;
Z – число связанных активных центров, при концентрации вкусового вещества, равного С.
Согласно закона действующих масс, постоянная равновесия данной реакции:
K=![]() (2)
(2)
Величина ответа рецепторной клетки прямо пропорциональна числу активных связанных центров R=Z, а максимальный отклик имеет место в том случае, когда все центры заняты. Тогда уравнение (2) можно записать в виде:
![]()
![]() (3)
(3)
Уравнение (3) представляет собой основное уравнение вкусовой рецепции. Величина C/R выражает суммарную реакцию вкусовых рецепторов. Согласно данному уравнению, между C/R и С существует линейная зависимость. Данное положение получило экспериментальное подтверждение.
Бейдлер предполагал, что взаимодействие частиц вкусового вещества с активными центрами рецептора, представляет собой физический процесс адсорбции, в основе которого лежит слабое дисперсионное взаимодействие. Это предположение согласуется с предсказанием Акри об образовании водородных связей между молекулами сладкого стимула и рецептора. На основании экспериментальных данных и уравнения вкусовой рецепции (3), можно найти величину (константы) равновесия конкретной реакции. Она изменяется от 7,7 для бутирата натрия до 9,8 для NaCl. Эти величины – константы равновесия, очень близки к коэффициенту для адсорбции солей на белках. Это, в свою очередь, соответствует исходному допущению Бейдлера об адсорбции вкусового вещества на поверхности рецептора.
О
физической природе процесса свидетельствует
также и то, что реакция вкусовых
рецепторов на соленый стимул не
изменяется при повышении температуры
от 20 до
![]() С.
Кроме того, величина реакции заметно
не изменяется при сдвиге pH
от 3 до 11, что исключает возможность
сильного электростатического
взаимодействия с иногенными группами
белковых молекул. Термодинамические
расчеты показывают, что процесс адсорбции
частиц приводит либо к уменьшению
количества гидратационной воды на
поверхности рецептора, либо к небольшому
изменению конформации молекул рецептора.
Возможно, что изменение конформаций
рецепторных молекул лежит в основе
дальнейшего процесса, приводящего в
конечном счете к возбуждению вкусовой
клетки.
С.
Кроме того, величина реакции заметно
не изменяется при сдвиге pH
от 3 до 11, что исключает возможность
сильного электростатического
взаимодействия с иногенными группами
белковых молекул. Термодинамические
расчеты показывают, что процесс адсорбции
частиц приводит либо к уменьшению
количества гидратационной воды на
поверхности рецептора, либо к небольшому
изменению конформации молекул рецептора.
Возможно, что изменение конформаций
рецепторных молекул лежит в основе
дальнейшего процесса, приводящего в
конечном счете к возбуждению вкусовой
клетки.
Дальнейшее развитие теории вкусового восприятия было связано с открытием Дастоли сладкочувствительных и горькочувствительных белков во вкусовых луковицах млекопитающих. Дастоли удалось выделить из специфических сладкочувствительных и горькочувствительных сосочков белки, которые избирательно взаимодействуют со сладкими или горькими веществами. Термодинамическая и кинетическая особенности взаимодействия сладких и горьких веществ с молекулами выделенных белков, совпадают с особенностями мономолекулярных реакций, описанных Бейдлером. В этой связи в настоящее время считают, что именно этими белками представлены активные центры вкусовых клеток, однако, локализация этих белков во вкусовых клетках еще не установлена.
БИОФИЗИЧЕСКИЕ ОСНОВЫ ВОСПРИЯТИЯ ЗАПАХА
В основе обоняния лежит взаимодействие молекул пахучих веществ с обонятельными клетками. Молекулы пахучих веществ, отделяясь от своей основной массы и передвигаясь потоками воздуха, могут действовать на расстоянии.
Обонятельные
рецепторы входят в состав обонятельного
эпителия, расположенного в задней части
носовой перегородки и в верхнем носовом
ходе. Рецепторные клетки представляют
собой биполярные нейроны d
примерно 5-10 мкм, расположенные вокруг
цилиндрических опорных клеток. У
человека количество обонятельных
рецепторов (ОР) достигает примерно![]() ,
а площадь обонятельного эпителия (ОЭ)
составляет примерно 5
.
Поверхность ОЭ покрыта водянистой
средой. Периферические отростки
обонятельных клеток заканчиваются на
своей вершине утолщение грушевидной
формы, которое называют обонятельной
булавой. Как показали электронно-микроскопические
исследования, на поверхности каждой
обонятельной булавы (ОБ) расположено
от 9 до 16 очень тонких ресничек. Центральные
отростки отходят от противоположного
конца обонятельных клеток и образуют
тонкие нервные нити, которые вступают
в полость черепа. На нижней поверхности
лобной доли нервные нити сходятся,
образуя обонятельную луковицу, в
клубочках которой и заканчивается
первый нейрон обонятельного афферентного
пути.
,
а площадь обонятельного эпителия (ОЭ)
составляет примерно 5
.
Поверхность ОЭ покрыта водянистой
средой. Периферические отростки
обонятельных клеток заканчиваются на
своей вершине утолщение грушевидной
формы, которое называют обонятельной
булавой. Как показали электронно-микроскопические
исследования, на поверхности каждой
обонятельной булавы (ОБ) расположено
от 9 до 16 очень тонких ресничек. Центральные
отростки отходят от противоположного
конца обонятельных клеток и образуют
тонкие нервные нити, которые вступают
в полость черепа. На нижней поверхности
лобной доли нервные нити сходятся,
образуя обонятельную луковицу, в
клубочках которой и заканчивается
первый нейрон обонятельного афферентного
пути.
При действии обонятельных стимулов на обонятельные рецепторы, происходит формирование генераторных потенциалов, которые вызывают в центральных отростках рецепторов нервные импульсы. Суммарный генераторный потенциал ОР, то есть, электроольфактограмму, можно зарегистрировать с помощью электрода, приложенного к поверхности ОЭ. Вопрос ОБ избирательности ОР к молекулам пахучих веществ окончательно не выяснен. До настоящего времени не было обнаружено строгой избирательности рецепторов. Так, например, некоторыми учеными с помощью микроэлектродов регистрировались потенциалы одиночных волокон обонятельного нерва лягушки, при действии различных пахучих веществ. Все исследуемые рецепторные нейроны обнаружили грубую избирательную чувствительность запаха. При этом, каждый из них, отвечал на одни запахи, не отвечал на другие. Большинство из них дает особо сильный ответ, по крайней мере, на 1 из 25 использованных запахов, и более слабо отвечает на остальные.
ТЕОРИЯ ОБОНЯНИЯ
В основе возникновения обонятельного ощущения лежит взаимодействие молекул или частиц пахучего вещества с ОР, однако, до настоящего времени неясно, какие свойства молекул обусловливают это взаимодействие. Не ясно также, с молекулами каких веществ, локализованных в рецепторных клетках, происходит взаимодействие. Попытки объяснить пахучие свойства молекул их химическими свойствами и структурой, не увенчались успехом. Вещества со сходными свойствами и структурой, могут обладать разными запахами и наоборот. В настоящее время, из всех теорий восприятия запаха, наибольшее внимание заслуживают две:
квантовая;
стереохимическая.
Согласно квантовой теории запаха, молекулам пахучего вещества свойственны внутримолекулярного колебания, в результате которых они испускают электромагнитное излучение в инфракрасной области спектра (ИК). Это излучение взаимодействует с молекулами обонятельного рецептора пигмента, типа каротиноидов, находящихся в мембране рецепторов. По мнению авторов данной теории, молекулы веществ с похожими запахами, должны характеризоваться сходными НЧ колебаниями. В качестве подтверждения, Райд приводит вещества, обладающие миндальным запахом и близкой частотой колебаний: нитробензол, бензонитрил, бутиронитрил и т.д. из обонятельных клеток удалось выделить ряд каротиноидов и витамин А, которые, по мнению Райда, поглощают электромагнитные излучения молекул пахучих веществ.
Однако, несмотря на известные успехи, данная теория встречает ряд очень серьезных возражений. Так, многие вещества со сходными запахами имеют разные частоты колебаний и разные спектры поглощения в ИК области (например, многие спирты). Молекулы, в которых атом водорода замещен изотопом дейтерия, обладают сходными запахами. Хотя, при этом значительно меняются частоты основных колебаний, с другой стороны молекулы с почти одинаковыми частотами колебаний и спектрами ИК поглощения, обладают несходными запахами, кроме того, не установлена локализация пигментов в обонятельных клетках и не доказано их участие в первичном процессе взаимодействия с пахучими молекулами.
Более обоснованной является стереохимическая теория восприятия запаха, выдвинутая Монкрифом и подробно разработанная Эймуром. Согласно этой теории, запах вещества обусловлен не химическим составом молекулы, а их формой и размерами. Обонятельная система состоит из рецепторов разных типов, каждый из которых соответствует отдельному первичному запаху. Поверхности рецепторных клеток имеют углубления (лунки) определенной формы. Молекулы пахучих веществ вызывают ощущение запаха только в том случае, если их форма соответствует форме лунок рецепторов, в которые они плотно входят. Этот принцип аналогичен принципу «замка и ключа», при взаимодействии фермента и субстрата. Согласно представлениям Эймура, вещества, имеющие сходный запах, должны обладать сходной формой своих молекул. Эймур, сопоставив большое количество данных о форме молекул с ощущениями, которые они вызывают, пришел к выводу о наличии 7 первичных простых запахов:
камфорноподобного;
мускусного;
цветочного;
мятного;
эфирного;
острого;
гнилостного.
В этом смысле, первичные запахи аналогичны трем основным цветам зрительного и четырем вкусам вкусовых анализаторов. Каждому первичному запаху соответствует определенная форма молекулы и лунки на поверхности рецептора. Так, эфирная молекула отличается палочковой формой, при этом, она должна иметь вытянутую лунку с размерами: длина = 1,8 нм, ширина = 0,5 нм, глубина = 0,4 нм. Камфорная молекула имеет сферическую форму, диаметром 0,7 нм, а ее лунка имеет форму эллиптической чаши, глубиной 0,4 нм, длиной = 0,2 нм, шириной = 0,75 нм. Молекулы, вызывающие другие ощущения, имеют более сложную форму. Если сложная молекула внедряется сразу в две лунки, то возникает сложный запах.
Теория Эймура нашла свое подтверждение, что, в некоторых случаях, оказывалось возможным предсказывать запах веществ, исходя из формы их молекул. Так было синтезировано несколько органических веществ, была рассчитана их вероятная форма и предсказан их запах. Однако есть недостатки: острые и гнилостные запахи не вмещаются в стереохимическую схему. Для молекул, вызывающих острые и гнилостные запахи, форма и размер не имеют значения. Решающую роль играет электрический заряд. Острые едкие запахи свойственны тем соединениям молекулы, которые из-за нехватки электронов, имеют «+» заряд и сильное сродство к электронам. Гнилостные запахи, наоборот, вызываются молекулами, обладающими избытком электронов.
Необходимо заметить, что данная теория обходит молчанием вопрос о том, что происходит после попадания молекулы в соответствующую лунку на поверхности рецептора? С чем она взаимодействует в ней при этом? Теория не объясняет, как возникают потенциалы в ответ на заполнение лунок?
ЭЛЕКТРИЧЕСКИЕ И МАГНИТНЫЕ СВОЙСТВА ТКАНЕЙ ОРГАНИЗМА
Современные преставления о магнитных свойствах живых тканей основаны на фактах о молекулярной организации БМ и, в значительно меньшей степени, на сведениях о кванотовомеханических свойствах физиологически активных молекул. Характеризуя электрические свойства живых тканей, следует учитывать, что они являются композиционными средами, поскольку одни структурные элементы обладают свойствами проводников, а другие - диэлектриков. Предполагают, что ряд биологически важных макромолекул проявляет полупроводниковые свойства.
ЭЛЕКТРОПРОВОДНОСТЬ ЖИВЫХ ТКАНЕЙ
Электропроводность
органов и тканей связана с присутствием
в них ионов, которые являются свободными
зарядами, создающими в организме ток
проводимости под действием ЭМП,
излучаемыми как внешними источниками,
так и генерируемых живыми клетками.
Электропроводность живых тканей
определяется прежде всего электрическими
свойствами крови, лимфы, межклеточной
жидкости и цитозоля. Удельная
электропроводность (
)
этих электролитов составляет 0,1-1
![]() .
Подвижность ионов в биологических
жидкостях примерно такая же, как в
растворах соответствующих солей,
приготовленных на дистиллированной
воде.
.
Подвижность ионов в биологических
жидкостях примерно такая же, как в
растворах соответствующих солей,
приготовленных на дистиллированной
воде.
Однако целых органов на 4-6 ниже жидкостей, выделенных из них. Причиной не совпадений результатов измерений являются малые объемы, занимаемые свободными электролитами в органах и тканях животных. В клетке электролиты заключены в мельчайшие отсеки (компартаменты), образованные БМ, составляющие более 50% массы клетки. По существу каждая клеточная органелла представляет собой компартамент. Ее содержимое и окружающий цитозоль обладают относительно высокой электропроводностью, тогда как разделяющая их мембрана является типичным диэлектриком.
Удельное
сопротивление плазматических мембран
измерить достаточно трудно, так как
невозможно измерить толщину мембраны.
В этой связи удается измерить только
удельное поверхностное сопротивление.
У различных клеток удельное поверхностное
сопротивление изменяется в пределах
от 0,4
![]() до 30
.
до 30
.
Живым
тканям свойственна зависимость
электропроводности от частоты
воздействующего ЭМП. Это явление
получило название дисперсии
электропроводности.
С повышением частоты электропроводность
тканей увеличивается. Д исперсия
электропроводности присуща всем средам,
а не только биологическим. Она наблюдается
в том диапазоне частот ЭМП, которые
соответствуют характеристическим
частотам (
исперсия
электропроводности присуща всем средам,
а не только биологическим. Она наблюдается
в том диапазоне частот ЭМП, которые
соответствуют характеристическим
частотам (![]() )
заряженных частиц, входящих в состав
той или иной среды. Поскольку однородные
среды образованы частицами с близкими
значениями
, то дисперсия электропроводности в
них выражена слабо. В этой связи
сопротивление резисторов в цепях
переменного тока называют активным, в
отличие от реактивных сопротивлений
емкости и индуктивности (
)
заряженных частиц, входящих в состав
той или иной среды. Поскольку однородные
среды образованы частицами с близкими
значениями
, то дисперсия электропроводности в
них выражена слабо. В этой связи
сопротивление резисторов в цепях
переменного тока называют активным, в
отличие от реактивных сопротивлений
емкости и индуктивности (![]() и
и
![]() ),
которые гораздо сильнее зависят от
частоты, чем R.
По той же причине редко утверждается,
будто дисперсия электропроводности –
специфическое свойство биологических
систем. На самом же деле у них зависимость
),
которые гораздо сильнее зависят от
частоты, чем R.
По той же причине редко утверждается,
будто дисперсия электропроводности –
специфическое свойство биологических
систем. На самом же деле у них зависимость
![]() гораздо отчетливее, чем у сред с менее
сложной организацией, и обнаруживается
в широком частотном диапазоне. Это
обусловлено сложной мембранной
структурой тканей и большим разнообразием
релаксационных способностей их
заряженных частиц, причем такое
разнообразие связано, как с различием
в размерах, так и с влияние на их
подвижность БМ. Повреждение клеточных
мембран стирает в значительной мере
грань между тканями и органическими
электролитами в дисперсии
на низких частотах.
гораздо отчетливее, чем у сред с менее
сложной организацией, и обнаруживается
в широком частотном диапазоне. Это
обусловлено сложной мембранной
структурой тканей и большим разнообразием
релаксационных способностей их
заряженных частиц, причем такое
разнообразие связано, как с различием
в размерах, так и с влияние на их
подвижность БМ. Повреждение клеточных
мембран стирает в значительной мере
грань между тканями и органическими
электролитами в дисперсии
на низких частотах.
ДИЭЛЕКТРИЧЕСКИЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ
Диэлектрические
свойства биологических тканей
определяются присутствием в них воды,
растворенных в ней макромолекул, а
также компартамезацией клеточных
структур. Компартамезация, обеспечивая
оптимальные условия для внутриклеточных
биохимических процессов, вместе с тем
придает живым тканям сегнетоэлектрические
свойства. Показано, что физические
среды, в которых чередуются слои очень
низкой и высокой
, обладают сегнетоэлектрическими
свойствами, если по обе стороны каждого
слоя с низкой
присутствуют разные электрические
заряды. Такие структуры подобны доменам
с одинаковой ориентацией дипольных
моментов. Цитозоль и содержимое многих
органелл обладают относительно высокой
![]() ,
а БМ – крайне низкой
(порядка
,
а БМ – крайне низкой
(порядка
![]() ). Поэтому каждая органелла на БМ которой
поддерживается разность потенциалов
между цитозолем и ее содержимым, имеет
значительный дипольный момент подобно
домену в сегнетоэлектрике. За счет
таких заряженных компартаментов живые
ткани обладают высокой
. В постоянном электрическом поле оно
достигает десятка тысяч.
). Поэтому каждая органелла на БМ которой
поддерживается разность потенциалов
между цитозолем и ее содержимым, имеет
значительный дипольный момент подобно
домену в сегнетоэлектрике. За счет
таких заряженных компартаментов живые
ткани обладают высокой
. В постоянном электрическом поле оно
достигает десятка тысяч.
Как
и всякому домену, каждому внутриклеточному
компартаменту присуща не высокая
характеристическая частота релаксации.
Применительно к диполям
![]() соответствует максимальной частоте
ЭМП, которую они способны воспроизвести
своими поворотами в нем, за счет чего
достигается максимально возможная
компенсация внешнего поля собственным
ЭП диэлектрика. Д
соответствует максимальной частоте
ЭМП, которую они способны воспроизвести
своими поворотами в нем, за счет чего
достигается максимально возможная
компенсация внешнего поля собственным
ЭП диэлектрика. Д иапазон
разных внутриклеточных компартаментов
находиться в пределах от Гц до 1 кГц.
Поэтому компартаменты вносят основной
вклад в диэлектрические свойства
биологических тканей именно на низких
частотах.
иапазон
разных внутриклеточных компартаментов
находиться в пределах от Гц до 1 кГц.
Поэтому компартаменты вносят основной
вклад в диэлектрические свойства
биологических тканей именно на низких
частотах.
На более высоких частотах диэлектрические свойства определяются полярными макромолекулами, сосредоточенными как в цитоплазме, так и в клеточных мембранах. В СВЧ полях основной вклад в эти свойства вносит вода.
У разных белковых молекул охватывает диапазон от 10 кГц до 100 мГц и зависит от их размеров, а также от вязкости окружающей среды. Эту зависимость выражает формула:
![]() ,
,
где K – постоянная Больцмана; T – температура среды, в которой вращаются молекулы, под действием ЭМП; - вязкость этой среды; -радиус полярной молекулы.
Важно отметить, что молекулы одинаковых размеров, пребывая в средах с разной вязкостью (например в цитозоле и БМ) обладают не одинаковой .
Характеристическая
частота релаксации внутриклеточный
воды такая же, как и дистиллированной
(![]() ).
Дипольные моменты у них также одинаковы
(~1,84 дебая). Именно в воде, входящей в
состав живых тканей, происходят основные
диэлектрические потери при действии
на организм СВЧ излучений поскольку
).
Дипольные моменты у них также одинаковы
(~1,84 дебая). Именно в воде, входящей в
состав живых тканей, происходят основные
диэлектрические потери при действии
на организм СВЧ излучений поскольку
![]() совпадает частота сантиметровых
радиоволн.
совпадает частота сантиметровых
радиоволн.
Неодинаковые
величины
разных тканевых компартаментов,
способных поляризоваться в ЭМП,
обуславливают неравномерный ход кривой
![]() ,
отображающей зависимость
ткани от
,
отображающей зависимость
ткани от
![]() ЭМ колебаний, воздействующих на нее.
На зависимости можно выделить три
участка, где кривая идет круче, чем в
промежутках. Эти участки называют
зонами дисперсии (релаксации) и их
обозначают греческими буквами
ЭМ колебаний, воздействующих на нее.
На зависимости можно выделить три
участка, где кривая идет круче, чем в
промежутках. Эти участки называют
зонами дисперсии (релаксации) и их
обозначают греческими буквами
![]() .
.
Первый
участок (
-
дисперсия) соответствует низкочастотному
диапазону до 1 кГц. Он обусловлен
поляризацией внутриклеточных
компартаментов, с которыми связаны
сегнетоэлектрические свойства живых
тканей. В силу значительной инерционности
релаксационных процессов в доменах -
компартаментах вращение этих гигантских
диполей запаздывает относительно
направления
![]() внешнего ЭМП даже на низких частотах,
что проявляется в уменьшении
по мере увеличения
. Некоторый вклад в
- дисперсию вносит релаксация зарядов
на фасциях, внутриорганных соединительно
тканных прослойках, клеточных
поверхностях.
внешнего ЭМП даже на низких частотах,
что проявляется в уменьшении
по мере увеличения
. Некоторый вклад в
- дисперсию вносит релаксация зарядов
на фасциях, внутриорганных соединительно
тканных прослойках, клеточных
поверхностях.
Второй
участок (
-
дисперсия) отображает изменение
поляризации макромолекул по мере
повышения частоты внешнего ЭМП. В
скелетной мышце
-дисперсия наблюдается в диапазоне
частот от
![]() до
до
![]() Гц. Снижение
по мере повышения
в этом диапазоне зависит от того, что
все менее крупные макромолекулы не
успевают поворачиваться в соответствии
с частотой внешнего ЭМП, когда она не
успевает превосходить
Гц. Снижение
по мере повышения
в этом диапазоне зависит от того, что
все менее крупные макромолекулы не
успевают поворачиваться в соответствии
с частотой внешнего ЭМП, когда она не
успевает превосходить
![]() той или иной полярной молекулы. Очевидно,
что
Гц является частотой соответствующей
наименее инерционных пептидных молекул.
той или иной полярной молекулы. Очевидно,
что
Гц является частотой соответствующей
наименее инерционных пептидных молекул.
Третий
участок (
-
дисперсия) приходиться на частоты выше
![]() Гц, чему соответствует
Гц, чему соответствует
![]() .
Поскольку воде свойственно несколько
значений
лежащих около 20 ГГц (явление многоструктурной
воды), то изменение
уменьшиться потому, что даже такие
мелкие молекулы, как
,
не успевают совершать повороты с
частотой, соответствующей частотному
диапазону
-дисперсии.
.
Поскольку воде свойственно несколько
значений
лежащих около 20 ГГц (явление многоструктурной
воды), то изменение
уменьшиться потому, что даже такие
мелкие молекулы, как
,
не успевают совершать повороты с
частотой, соответствующей частотному
диапазону
-дисперсии.
На
более высоких частотах (до
![]() Гц) может проявляться электронная
поляризация, характерная для атомов,
и обусловленная смещением оболочек у
атомов.
Гц) может проявляться электронная
поляризация, характерная для атомов,
и обусловленная смещением оболочек у
атомов.
В
живых тканях, ввиду того, что им наряду
с электропроводностью присущи другие
свойства, под действием внешнего ЭМП
возникают и токи проводимости, и токи
смещения. По мере повышения частоты
ЭМП роль тока смещения в биологическом
эффектах электромагнитного излучения
возрастает и становиться ведущей на
частоте выше
![]() Гц. Сказанное выше хорошо характеризует
различные виды высокочастотной
электротерапии: если при диатермин
(
Гц. Сказанное выше хорошо характеризует
различные виды высокочастотной
электротерапии: если при диатермин
(![]() МГц)
тепловой эффект связан прежде всего с
током смещения.
МГц)
тепловой эффект связан прежде всего с
током смещения.
Клеточные
мембраны принято рассматривать по
аналогии с диэлектриком в виде плоских
конденсаторов. В этой связи поведение
мембран при воздействии ЭМП характеризует
электрическая емкость, приведенная к
1
мембранной поверхности. Эта величина
(![]() )
называется удельной емкостью БМ и имеет
размерность
)
называется удельной емкостью БМ и имеет
размерность
![]() .
Очень велика
плазмолемы гладкомышечного волокна
млекопитающих - около 30
.
Нервные волокна многих животных имеют
плазматические мембраны, … которых
находиться в пределах единиц
.
.
Очень велика
плазмолемы гладкомышечного волокна
млекопитающих - около 30
.
Нервные волокна многих животных имеют
плазматические мембраны, … которых
находиться в пределах единиц
.
Чтобы учесть зависимость энергетических преобразований на емкости от частоты внешнего ЭМП, используется понятие емкостного сопротивления:
![]()
![]()
Эта величина служит коэффициентом преобразования энергии внешнего ЭМП в электрическую энергию, накапливаемую конденсатором в течение половины периода воздействующих ЭМ колебаний. 4 см больше … БМ, тем меньший электрический заряд она способна удержать на себе и тем меньшая доля энергии внешнего ЭМП превратиться в электрическую энергию.
МАГНИТНЫЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ
Любая
среда является одновременно электриком
(проводником или диэлектриком) и
магнитиком. Магнитные свойства
биологических тканей характеризуются
довольно низкой величиной магнитной
проницаемости (![]() )
близкой к 1, поскольку основные химические
компоненты биосред (белки, углеводы,
липиды, вода) относятся к диамагнетикам.
Их почти нулевая магнитная восприимчивость
(
)
близкой к 1, поскольку основные химические
компоненты биосред (белки, углеводы,
липиды, вода) относятся к диамагнетикам.
Их почти нулевая магнитная восприимчивость
(![]() )
служит одной из причин недостаточного
внимания к изучению магнитных процессов
в организме. У человека обнаружены
ферритинсодержащие включения (в
надпочечниках). Их функции остаются не
выясненными. Предполагают, что подобные
ферромагнитные включения присутствуют
в тканях пчел, бабочек, дельфинов,
обеспечивая их пространственную
ориентацию. Вопрос о ферромагнитных
свойствах биологических систем далеко
от его разрешения.
)
служит одной из причин недостаточного
внимания к изучению магнитных процессов
в организме. У человека обнаружены
ферритинсодержащие включения (в
надпочечниках). Их функции остаются не
выясненными. Предполагают, что подобные
ферромагнитные включения присутствуют
в тканях пчел, бабочек, дельфинов,
обеспечивая их пространственную
ориентацию. Вопрос о ферромагнитных
свойствах биологических систем далеко
от его разрешения.
Вместе с тем в исследованиях биомагнетизма недостаточно учитывается важное положение электродинамики, согласно которому коэффициентом преобразования энергии ЭМП в магнитную энергию среды, подвергнутой его воздействию, является не сама по себе, а индуктивность, отображающая как собственно магнитные свойства, так и ее геометрические особенности. Даже диамагнетики способны осуществлять подобное преобразование довольно эффективно, если образуют структуры в форме катушек, по которым течет электрический ток, наведенный внешним полем. (Аналог магнитные поля катушек из медного провода) Для учета зависимости энергетических преобразований на индуктивности на частоте внешнего ЭМП введено понятие индуктивного сопротивления:
![]()
где - длина катушки, по которой течет ток; S - площадь каждого витка; n - число витков в катушке.
Оказалось,
что 1
мембраны аксона кальмара толщиной
примерно 10 нм имеет
![]() Гн. Такая индуктивность присуща медному
проводу длиной в 1 милю (1600м), намотанный
на железный сердечник массой в 1 фунт
(~450гр). Это очень высокая индуктивность.
Гн. Такая индуктивность присуща медному
проводу длиной в 1 милю (1600м), намотанный
на железный сердечник массой в 1 фунт
(~450гр). Это очень высокая индуктивность.
И все же биофизические основы биомагнитных явлений еще не изучены, хотя с давних пор люди верили в действие магнитных полей на биологические системы. О лечебных свойствах магнита писал Аристотель (IV в до н.э.) Гален (III в до н.э.) применял магнит как средство от запоров. Авиценна (XI в) воздействовал на патологические процессы в селезенке Парацельс (XIV в) применял магнит при многих болезнях, будучи уверенным в том, что «… магнит оттягивает грыжу и исцеляет перелом, вытягивает желтуху и оттягивает водянку».
С.П. Боткин в 70 годах прошлого века утверждал, что магнит может создавать ощущение зуда, покалывания или боли, восстанавливать нарушенную чувствительность кожи, купировать судороги, вызывать общую слабость и сонливость. В зависимости от исходного состояния пациента магнит зачастую приводил к эффектам, противоположным тем, на которые рассчитывал врач, например вместо ослабления боли усиливал ее.
Современная медицинская литература богата сообщениями о лечебном действии магнитного поля при атеросклерозе и гипертонической болезни, бронхиальной астме, неврозе и многих других патологических состояниях. Однако нельзя быть уверенным в том, что в благоприятном действии магнита нет больше доли психотерапевтического воздействия на больного самой процедурой магнитотерапии. До сих пор в биофизике нет достаточно обоснованной рабочей гипотезы, которую можно положить в основу научного изучения биомагнетизма.
ДИСПЕРСИЯ ИМПЕДАНСА ЖИВЫХ ТКАНЕЙ
Отражением современного состояния магнитобиологии является общепринятое мнение, будто электрический импеданс (полное сопротивление переменому току) живых тканей имеет только две состаляющие – резистивное и емкостное сопротивление:
![]()
Индуктивное сопротивление не учитывается.
Как
и в электрических цепях, импеданс
биологических систем зависит от частоты
переменного тока. Для живых тканей
характерно уменьшение импеданса по
мере увеличения этой частоты. Зависимость
Z
от
получила
название дисперсии импеданса. М ежду
дисперсиями электропроводности,
диэлектрической проницаемости, о
которых идет речь шла выше, и дисперсией
импеданса существует связь, но это не
идентичные процессы. Например, крутые
и пологий участки дисперсий
и Z
в одной и той же ткани обычно не совпадают.
Дисперсия импеданса отображает,
очевидно, более широкий круг
электромагнитных процессов в биологических
системах (возможно и индуктивные
свойства). Во всяком случае, она сильнее
зависит от разнообразных нарушений
жизнедеятельности тканей.
ежду
дисперсиями электропроводности,
диэлектрической проницаемости, о
которых идет речь шла выше, и дисперсией
импеданса существует связь, но это не
идентичные процессы. Например, крутые
и пологий участки дисперсий
и Z
в одной и той же ткани обычно не совпадают.
Дисперсия импеданса отображает,
очевидно, более широкий круг
электромагнитных процессов в биологических
системах (возможно и индуктивные
свойства). Во всяком случае, она сильнее
зависит от разнообразных нарушений
жизнедеятельности тканей.
По
кривой дисперсии импеданса возможно
судить об уровне обмена веществ и его
отклонениях от нормы. В медико-биологических
экспериментах и клинике все шире
применяется метод изучения дисперсии
Z
для оценки жизнеспособности органов
и тканей. Обычно измеряют всего два
значения Z:
на низкой (![]() Гц)
и высокой (
Гц)
и высокой (![]() Гц)
частотах, соответствующим тем частотным
диапазонам, где кривая дисперсии
импеданса идет более полого, чем на
среднечастотном участке крутого спада.
Отношение этих величин называют
коэффициентом поляризации (
Гц)
частотах, соответствующим тем частотным
диапазонам, где кривая дисперсии
импеданса идет более полого, чем на
среднечастотном участке крутого спада.
Отношение этих величин называют
коэффициентом поляризации (![]() ):
):
![]()
где
![]() - импеданс на низкой частоте;
- импеданс на низкой частоте;
![]() – на высокой частоте.
– на высокой частоте.
Жизнеспособная
ткань имеет
![]() , причем
тем больше, чем выше уровень обмена
веществ в данной ткани и чем лучше
сохраняется ее структурная целостность.
При отмирании ткани ее
стремиться
к 1.
, причем
тем больше, чем выше уровень обмена
веществ в данной ткани и чем лучше
сохраняется ее структурная целостность.
При отмирании ткани ее
стремиться
к 1.
Кроме
используют, так называемый коэффициент
частотной дисперсии (![]() ):
):
![]()
Метод исследования дисперсии импеданса применяют для оценки жизнеспособности тканевых трансплантантов при пересадке органов. Изучают возможности использования его для определения зон раневого процесса в ходе хирургической обработки раны, для характеристики ишемии, отека и т.д.
Широкое распространение в медицинской практике нашла методика реоплетизмографии. Посредством ее изучают активную составляющую импеданса (R), которая зависит прежде всего от кровенаполнения исследуемого органа. Чем больше крови содержится в органе, тем ниже (при прочих равных условиях) его электрическое сопротивление. Это позволяет оценивать органное кровообращение путем измерения R органа переменному току.
По динамике электрического сопротивления кожи судят о так называемых кожно-гальванических реакциях (КГР), в которых отображаются эмоции, утомление и другие состояния организма. Наконец специалисты в области рефлексотерапии измеряют электрическое сопротивление кожи для нахождения «активных точек», воздействие на которые оказывают разнообразные лечебные эффекты. Есть и другие применения в медицинской практике тех или иных способов измерения электрического импеданса, его составляющих и арсенал различных методов исследования электрических свойств биологических систем неуклонно нарастает.