

3. Подтип Хелицеровые (Chelicerata)
Включает класс первичноводных мечехвостов и класс наземных паукообразных (некоторые виды — вторичноводные). Тело подразделено на головогрудь и брюшко. На головогруди 6 пар конечностей: хелицеры, педипальпы и 4 пары ходильных ног. Хелицеры и педипальпы имеют разное строение в зависимости от способа питания. На брюшке плавательные ножки с жаберными придатками имеются только у мечехвостов. У паукообразных типичные конечности отсутствуют. Они преобразованы в легочные мешки, половые крышечки, паутинные бородавки и др. Дыхание у мечехвостов с помощью жабр, у паукообразных - легких (скорпионы, часть пауков), трахей (солытуги, клещи) или тех и других (многие пауки, в том числе паук-крестовик). Представители: мечехвосты (прибрежные участки тропических побережий океанов), скорпионы, фаланги пауки (паук-крестовик, тарантул, каракурт, серебрянка и др, клещи, в том числе собачий клеш и др.).
4. Подтип Трахейнодышащие (Tracheata)
Высшие трахейнодышащие членистоногие. Дышат только с помощью трахей. Имеется одна пара усиков, которая гомологична антеннулам (1-й паре антенн) ракообразных и принадлежит акрону. Сердце трубчатое, располагается на спинной стороне полости тела. Органы выделения -мальпигиевы трубочки. Подразделяется на 2 класса.
Класс 1. Многоножки (Myriapoda)
Низшие трахейные. Тело подразделено на 2 отдела - голову и туловище, состоящее из многих сегментов. Крыльев нет. Эпикутикулы (самый наружный, очень тонкий, влагонепроницаемый слой кутикулы) нет. Ведут скрытный образ жизни. Днем прячутся под корой, стволами деревьев и в других укрытиях.
Класс 2. Насекомые (Insecta)
Высшие по своей организации и степени освоения различных сред обитания членистоногие и вообще первичноротые. Крупный ароморфоз -приобретение в процессе эволюции насекомыми крыльев позволил им освоить воздушное пространство как новую среду обитания, без утраты наземной сферы как среды обитания. Тело состоит из трех отделов -головы, груди, брюшка. Строение ротового аппарата очень разнообразно и связано с характером пищи и способом ее приема. На груди располагаются 3 пары ног, имеющих различные модификации в связи с характером передвижения и среды обитания. Крыльев две или одна пара крыльев, которые нередко частично или полностью редуцированы в связи со специфическим образом жизни, например, у эктопаразитов. Самый наружный слой кутикулы - эпикутикула (сложный комплекс восков и липидов), хорошо защищающий организм от потери воды. Особенности ротового аппарата и всей пищеварительной системы связаны с характером питания. Органы дыхания - сложно разветвленная трахейная система. Развитие у большинства с неполным или полным превращением. Играют огромную роль в природе как опылители растений, потребители живой и мертвой органической продукции, мелиораторы почвы. Для человека полезны опылением культурных и дикорастущих покрытосеменных растений, как поставщики ценных пищевых продуктов и лекарственных материалов (мед, прополис и их производных, натуральный шелк), как регуляторы численности вредных видов (паразиты и хищники). Ряд видов насекомых - вредители сельского и лесного хозяйства (клоп вредная черепашка, колорадский жук, хлебный жук кузька, бабочки капустная белянка, луговой мотылек, озимая совка и др.).
20.Насекомые как высшие трахейнодышащие членисnоногие. Ароморфозы, позволившие им широко освоить наземную и воздушную среды. Важнейшие черты организации. Классификация, разнообразие, экологические адаптации, роль в природе и для человека.
Насекомые (Insecta) двустороннесимметричные, трехслойные, первичноротые, с сегментированным телом, гетерономной метамерией. Покровы - хитиновая кутикула; конечности членистые функции: локомоторную, защитную, органов схватывания и измельчения пищевых материалов, половых придатков. Полость тела смешенная – миксоцель; кровеносная система незамкнутая; рост сопровождается линьками. При этом ароморфозом – гетерономную сегментацию, членистые конечности и их прогрессивную дифференцировку в процессе адаптивной радиации, включая дифференцировку ротового аппарата. Приобретение в процессе эволюции крыльев) позволило крылатым насекомым освоить воздушное пространство как новую среду обитания. Тело состоит из 3 отделов - головы, груди, брюшка. Скрыточелюстные (Entognata)-грудной отдел слабо обособлен от брюшного. Есть мандибулы и максилы. На голове усики, простые глазки, на брюшке - двигательные, дыхание трахейное или только кожное. отр Бестяжковые, отр. Ногохвостки, отр. Двухвостки. Насекомые открыточелюстные (Entognatha) – тело состоит головы с усиками и 3 пар ротовых конечностей, брюшка без конечностей; дыхание трахейное. Достигают прогресса благодаря жизни на суши (строение покровов, многофункц особенности выделительной системы, трахейной, появление органов полета, особ эмбриогенеза и постэмбриогенеза обеспечивают независимость от факторов сухопутной среды. Пищеварительная система состоит из 3 отделов, есть слюнные железы, пищеварительные ферменты. Органы выделения: мальпигиевые сосуды (осморегуляции, накопления) Кров.система - незамкнутая, кровь – гемолимфа (снабжение органов питат вещ-вами ,гуморальная регуляция) , есть сердце. Дыхат: трахеи и дыхальца.(транспорт О2 к тканям). Н.С.: брюшная нервная цепочка, есть г.м. С ЦНС связана сист. симпатических нервов. Размножение половое, раздельнополые; диморфизм, партеногенез. развитие с превращением или нет. П.кл. Первичнобескрылые - низкие черты организации крыльев. Рот.аппарат - грызущий; раз - прямое. Отр.Щетинохвостки. П.кл. Крылатые (Pterygota): Поденки (Ephemeroptera), стрекозы (Odonata), Таракановые (Blattodea), Богомоловые (Mantodea), Термиты (Isoptera), Прямокрылые (Orthoptera), Вши (Anoplura), Равнокрылые (Homoptera), Клопы (Hemiptera), Сетчатокрылые (Neuroptera), Жуки (Coleoptera), Перепончатокрылые (Hymenoptera), Двукрылые (Diptera), Блохи (Aphaniptera), Ручейники (Trichoptera), Бабочки (Lepidoptera). Значение: важное звено пищевых цепей; производят полезные для человека продукты (мед, воск, прополис, пчелиный яд (медицина); объекты биотехнологии (для борьбы с нас вредителями).
21.Общая характеристика иглокожих как вторичноротых древних морских животных, обладающих вторичной радиальной симметрией. Особенности организации и образа жизни морских звёзд, морских ежей и голотурий.
Тип Иглокожие(Echinodermata) составляют морские целомические животные; вторичноротые. Распространены во всех соленых морях на разных глубинах. Сочетание билатеральной симметрии с радиальной(5-лучевой). Обладают известковым скелетом (функция: защита, опора для внутренних органов). Вторичная полость тела - целом, выполняет функции: полость для органов, гомеостаз внутренней среды, опора, транспорт. Характерна амбулакральная система – двигательная, на амбулакральных ножках, которые служат еще и для подачи пищи ко рту. Н.с. неганглиозного типа, закладывается в виде тяжа путем погружения внутрь зародыша эктодермальной пластинки. Дыхание через кожу, кожными жабрами, амбулакральными ножками, щупальцами. Органов выделения нет. Амебоцисты поглощают из целома экскреты, которые затем удаляют через кожу. Н.с. состоит из нервного кольца и отходящих от нее радиальных нервных стволов. Органы чувств: чувствительные клетки; равновесия (глубоководных голотурий). Раздельнополый деморфизм выражен слабо. Оплодотворение наружное; личинка – диплевра. Есть живородящие. 4 типа питания: зоо-, фито-, детрито- ,планктонофагия. Кл. Морские звезды (Asteroidea)- звездообразные формы. Различные типы питания, движения. Особенности – наличие радиальных лучей, в которые заходят органы. Характерно скользящие движение амбулакральных ножек. (Asterias rubens) Кл. Морские ежи (Echinoidea) Малоподвижные, донные, шаровидной или яйцевидной формы. Хорошо развит скелет, образующий панцирь; движутся на амбулакральных ножках. 2 Подкласса: Правильные ежи и Неправильные ежи. У правильных наличие жевательного аппарата - Аристотелев фонарь. Личинка - эхиноплетеус. Кл. Голотурии (Holothuroidea) Медленно ползают; тело продолговатое; билатеральная симметрия преобладает; детритофаги; раздельнополые, есть гермафродиты.
Роль: в биологических цепях, в почвообразовании, в состав осадочных пород - мрамор, известняк; голотурии в пищу.
22.Позвоночные. Череп и его эволюционные преобразования в ряду позвоночных. Черты прогресса и адаптивные особенности черепа позвоночных.
Развитие головного мозга и связанных с ним органов чувств требовало образования для их защиты новых скелетных образований. Эту функцию выполняет мозговой, или осевой череп. К тому же, переход к активному питанию подвижными пищевыми объектами вызывал необходимость образования дополнительных органов активного захвата и переработки пищи. Это обеспечено формированием висцерального скелета, окружающего передний конец пищеварительной трубки, а у водных позвоночных этот отдел служит и опорой жабр. Висцеральный (splanchnocranium) и мозговой (neurocranium) череп у всех позвоночных животных объединяется в единый комплекс - череп (cranium). Впервые в эволюционном ряду позвоночных он начинает формироваться у круглоротых.
Основные этапы эволюционные преобразования черепа наглядно прослеживаются при рассмотрении эмбрионального развития позвоночных.
Мозговой череп закладывается под головным мозгом в виде двух-трех пар хрящей. Одновременно, но независимо от них возникают хрящевые капсулы органов чувств: обонятельные, слуховые и глазные. Разрастаясь и сливаясь друг с другом, они начинают обрастать головной мозг с боков. Такая стадия развития мозгового черепа, еще лишенного крыши, соответствующая ранним этапам эмбрионального развития черепа остальных позвоночных, присуща круглоротым (рис. 1).
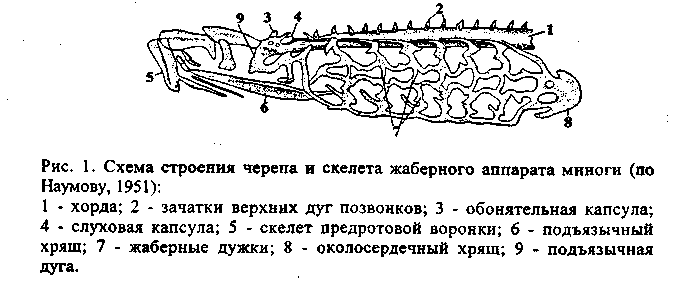
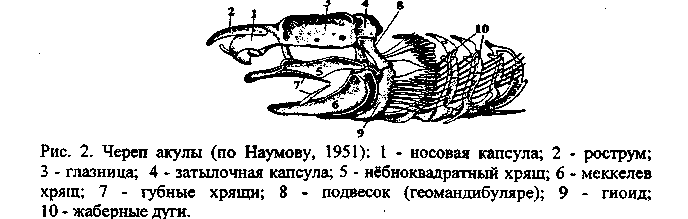
У остальных позвоночных еще на личиночной или зародышевой стадии разросшийся хрящ образует и крышу черепа. Такой, полностью хрящевой череп, у хрящевых рыб - акул и скатов (рис. 2).
У низших костных рыб (хрящевых ганоидов) мозговой череп остается хрящевым, а поверх него образуется панцирь из покровных кожных костей (рис. 3). У остальных групп костных рыб в первичном хрящевом мозговом черепе возникают окостенения, завершающиеся образованием так называемых первичных, или хрящевых, костей. Одновременно покровные кости погружаются под кожу и вступают в соединение с хрящевыми костями. У костистых рыб мозговой череп состоит из большого количества хрящевых и покровных костей; хрящ сохраняется только в области обонятельных и слуховых капсул и глазниц (рис. 4).
У современных амфибий в мозговом черепе остались значительные участки хряща и относительно небольшое число хрящевых и покровных костей (рис. 5).


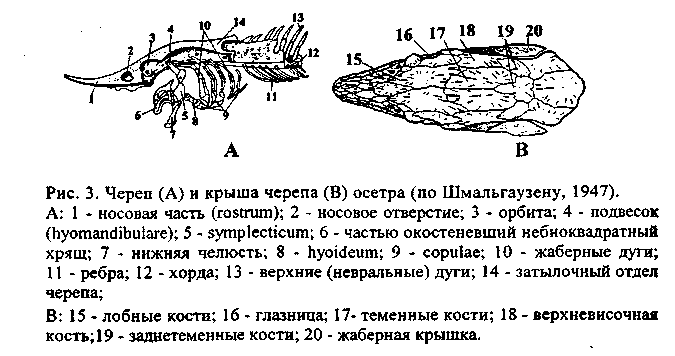
У рептилий, птиц и млекопитающих мозговой череп во взрослом :состоянии образован лишь костями. У птиц и млекопитающих большинство костей срастается между собой, так что их границ обнаружить нельзя. Впервые У некоторых пресмыкающихся, а позже и у остальных позвоночных, в мозговом черепе образуется вторичное твердое костное нёбо, разделяющее полость рта на верхний отдел - носоглоточный ход - и нижний отдел -собственно ротовую полость (рис. 6). Благодаря этому пища, находящаяся во рту, не мешает дыханию.
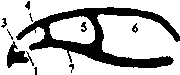
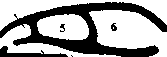
Рис. 6. Схема образования вторичного твердого костного нёба (по Шмальгаузену, 1947).
1 - первичные хоаны; 2 - вторичные хоаны; 3 - дыхательная и 4 - чувствующая части обонятельной полости; 5 - орбита; 6 - черепная полость; 7 - первичное нёбо; 8 - вторичное твёрдое нёбо; 9 - носоглоточный проход.
Эволюция жевательной мускулатуры, начиная с пресмыкающихся, сопровождалась перестройкой крыши и частично боковых стенок первично сплошного «стегального» черепа, имевшегося у древних земноводных -стегоцефалов, предков рептилий. В отдельных ветвях пресмыкающихся эволюция шла разными путями. В группе анапсида (ископаемые котилозавры и их потомки - черепахи) первичная стегальная (сплошная) крыша черепа сохранялась. В группе диапсида (гаттерия, крокодилы) образовались две височные ямы, ограниченные двумя костными дугами (рис. 7).
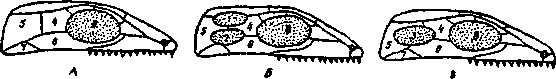
Рис. 7. Схема изменения строения черепа у пресмыкающихся (по Наумову и Карташеву, 1979):
А - стегальный, или анапсидный череп; Б - диапсидный череп с двумя височными ямами; В - синапсидный череп с одной боковой височной ямой. 1 - верхняя височная яма, 2 нижняя височная яма, 3 - единственная боковая височная яма, 4 — заглазничная кость, 5 - чешуйчатая кость, 6 - скуловая кость, 7 - квадратно-скуловая кость, 8 - орбита.
Верхняя дуга состоит из костей залобной (postfrontale) - чешуйчатой (squamosum), нижняя - из скуловой (]U8ale) - квадратноскуловой (quadratojugale). У ящериц диапсидный тип черепа с редуцированной нижней дугой, а у птиц - с редуцированной верхней дугой; у змей обе дуги редуцируются. В группе синапсид - у звероящеров (тероморфных рептилий) и происшедших от них млекопитающих - образовалась одна боковая яма и ограничивающая ее сложная дуга из костей верхней и нижней дуг: скуловой (jugale) - квадратноскуловой (quadratojugale) - чешуйчатой (squamosum).
По особенностям расположения мозга в черепной коробке различают два основных типа мозгового черепа. Платибазальный череп (круглоротые, хрящевые рыбы, амфибии) имеет широкое основание и мозговую полость, расположенную между его глазницами, а тропибазальный (костные рыбы, рептилии, птицы, млекопитающие) - узкое основание, глазницы, разделенные лишь тонкой межглазничной перегородкой, и мозговую полостью, размещенную позади глазниц (рис. 8).

По мере движения по эволюционному ряду позвоночных от костистых рыб к млекопитающим происходит постепенное уменьшение количества костей, образующих череп. Наименьшее количество их у млекопитающих.
Висцеральный отдел черепа развивается независимо от мозгового черепа в виде жаберных дуг, лежащих в перегородках между жаберными щелями. У круглоротых эти хрящевые зачатки преобразуются в хрящевую жаберную решетку околосердечный хрящ и в хрящи, поддерживающие язык и ротовую воронки.
У водных челюстноротых позвоночных животных рудименты первых двух жаберных дуг образуют губные хрящи (развиты у акуловых рыб). Третья дуга, которую называют челюстной, расчленяется на два отдела, образуя собственно челюсти: верхний элемент называют небно-квадратным хрящом (palatoquadratum), нижний - меккелевым хрящом (cartilage meckeli). Эти элементы выполняют функцию челюстей только у низших рыб (см. рис. 2).
У высших рыб они окостеневают и лишь частично участвуют в удержании добычи; а захват добычи у них переходит вторичными челюстями, образованными покровными костями: верхнечелюстной (maxillare) и предчелюстной (praemaxillare) - в верхней челюсти; зубной (dentale) - в нижней челюсти.
Следующая висцеральная дуга, подъязычная, обычно состоит из двух крупных парных хрящевых элементов: подвеска, или гиомандибулярного (hyomandibulare), и подъязычного, или гиоида (hyoideum). Гиоиды правой и левой сторон соединяются друг с другом при помощи небольшого непарного элемента - копулы (copulae). У высших рыб подъязычная дуга окостеневает (рис. 4).
Остальные дуги служат опорой жабр - жаберные дуги. У примитивных рыб число их может достигать 7 пар, у большинства хрящевых рыб - 5 пар хрящевых жаберных дуг, у костистых рыб часто их 4 пары (V-рудиментарна) и они окостеневают. За счет покровных костей у костных рыб формируется скелет жаберной крышки, тоже относящийся к висцеральному черепу.
Различают несколько типов прикрепления челюстного аппарата к мозговому черепу (рис. 9).
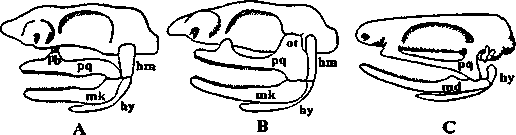
Рис. 9. Отношение передних висцеральных луг к осевому черепу (по Шмальгаузену, 1947).
А- гиостильный череп акулы; В - амфистильный череп акулы; С - аутостильный череп наземного позвоночного (лягушка); hm - hyomandibulare; hy - hyoideum; mk - меккелев хрящ; md - нижняя челюсть; ot - processus oticus; pb - processus palatobasalis; pq - palatoquadratum.
Протостилия - челюстная и подъязычная дуги независимо друг отдруга связками подвешивались к мозговому черепу. Этот гипотетический исходный тип черепа, вероятно, был свойствен примитивным челюстноротым.
Гиостилия - верхний конец гиомандибулярного отдела подъязычной дуги прикрепляется к слуховому отделу мозгового черепа и служит подвеском для челюстной дуги (многие группы рыб, в том числе большинство хрящевых и все костистые рыбы).
Амфистилия - верхний элемент челюстной дуги соединяется с мозговым черепом при помощи одного - двух специальных отростков и, кроме этого, как и при гиостилии, задние концы обоих челюстных элементов прочно связаны с нижним концом гиомандибулярного элемента (некоторые примитивные и современные акулы, костные ганоиды).
4. Аутостилия - верхний элемент челюстной дуги соединяется или срастается с мозговым черепом, гиомандибулярный элемент подъязычной дуги в прикреплении челюстей не участвует, и подъязычная дуга в большей или меньшей степени подвергается редукции (цельноголовые и двоякодышащие рыбы, земноводные и все другие наземные позвоночные).
С переходом к воздушному дыханию висцеральный скелет видоизменяется. В ряду земноводные - млекопитающие происходит усиление вторичных челюстей, а развитие аутостилии приводит к редукции подъязычной дуги: нижний ее элемент вместе с редуцирующимися жаберными датами участвует в образовании подъязычного аппарата и скелета гортани, а верхний элемент - гиомандибулярный, или подвесок - превращается в косточку среднего уха - стремечко. Скелет жаберной крышки полностью редуцируется.
У млекопитающих, в отличие от всех остальных позвоночных, нижнюю челюсть образует одна единственная парная кость - зубная (dentale).
23.Выделительная система в ряду хордовых. Особенности и направления её эволюции. Позвоночные. Способы поддержания гомеостаза. Терморегуляция. Осморегуляция.
Обеспечивает выделение жидкого конечного продукта обмена. Почки – главный орган выделения. Многие сталкиваются с проблемой осморегуляции, осморегуляция – один из параметров поддержания гомеостаза.
По величине осмотического давления растворы м.б. изотоническими (изоосмотич) – 2 р-ра им одинак давление, гипертонические – с более высок осмотич давл, гипотонич – с более низк. Водные организмы по этой величине м.б. изоосмотич, гипоосмотич.
В теле животных жидкости по своему состоянию напоминают морскую воду, они предст собой солев р-ры, электролит – NaCl, т.к. жидкость зародилась в море, организмы д были быть этой воде подобны. В то же время морская вода и жидкости тела не тождественны: и состав морск океана менялся, и состав жив менялся – проблема осморегуляции стоит и перед орг, жиб в воде. Морск орг: их жидкости менее концентрированы, чем окр среда – чистая вода поступает в тело. Удаление воды осущ ч/з жабры и почки. У морск рыбы чаще всего происх пассивн выд-е воды жабрами, нек кол-во изотонич мочи вывод ч/з почки. Пресноводные рыбы (и личинки амфибй): конц-ция вещ-в внутри тела намного превышает конц-цию вещ-в пресной воды. Пассивное поглощение воды телом и жабрами, почки выделяют оч большое кол-во гипотонич мочи с неб сод-ем солей.
Продукты экскреции: продукты распада АК и белков – соед-я, сод аминогр (токсичны) – выделение аммиака (м. олько круглоротые, рыбы и личинки аф) – амониотелия. Аммиак мгновенно растворяется в окр воде, легко вымывается, жабры выд аммиак, избыток аммиака накапл в мышцах. У др животных аммиак превращ-ся в мочевину (менее токсичное соед-е, но ещё ядовита и требует разведения). Выд-е мочевины – уреотелия. Мочевина растворима в воде – продукты выд-я оказ жидкими – выд достаточно больш кол-во мочи, осмотич св-ва м.б различны. Уреотелия: взрослые амф, млекопит. Мочевина обр в печени, поступает в кровь, ыд ч/з почки. У нек выд-ся мочевая к-та – урикотелия. Моч к-та не раств в воде, продукты выд сухие, мочи выд мало, но она высок конц – рептилии, птицы. Уринотелия – выд-е уринов (насек, паукообр) не св-на позвоночным.
У хордовых функ 2 осн гр орг:
Низшие – метанефридии – воронки с булавовидн обр со жгут – соленоциты.
Черепные – почки. Строение почек различно. Хар-на смена 3х поколений почек: в эмбриональном развитии набл 3 пок: предпочка (головная почка – пронефрос) – имеются внешние мальпигиевы клубочки, расп в целомич жидк, мальп кл – кап сеть, СОБР в клубок, со ст выд сист имеются воронки, откр в целом, имеется первичный мочеточник – пронефрос (пронефрич канал = вольфов канал – стекание мочи, выводн проток выделит сист). Сам перв почка обособл – воронка яйцевода, от неё отх ещё один канал – мюллеров. Им воронки с ресничками – нефростомы, кр сист обл мальп клубочками, прям контакта м/у кров и выд сист нет (обмен ч/з целомич полость). Пронефрос заклад-ся у эмбрионов практич у всех позвоночных, но функц только у нек круглоротых.
2е пок-е почек заклад-ся ниже – туловищная почка =первичная почка, мезонефрос). Имеется непосредств контакт кр и выд сист, жидк филтр непоср ч/з клубочки. Внтур мальп клубочки = мальп тельца. Мальп кл окружается капсулой (капсула Боумена) мальп клуб + капс Боумена место фильтрации) = мальп тельце.
У сам примит сох воронки (хрящ рыбы), в кач-ве мочеточников функц-ет Вольфов канал. У самок хорошо выражен Мюллеров канал, кот служит для выд-я прод. Тул почка функц-ет у амфибий, у хрящ и костн рыб.
У амниот (рептилии, птицы, млекопит) в эмбриональн развитии функц-ет первичн поки – мезонефросы, во взр сост ф-ет 3е пок-е почек – вторичные почки – метанефрос (тазовые почки). Развитие нефронов. Нефрон – структурно-функц ед почки. Нефрон предст собой мальп тельце, здесь происх фильтрация под давлением +в нефр им извитые канальцы (сост из канн 1го и 2го порядка). В обл канальцев им каппилярн сеть (активн экскреция – обр из первичн мочи вторичн моча, обратное всасывание воды в кровь и нек вещ-в, не подл выд-ю: глюкоза, нек соли), от метанефроса отщепл отдельн выделит канал – вторичн мочеточник, служит для выд-я конечн продукта обмена.
Рыбы, амфибии: Вольф канал – выд-е мочи и пермы. У самок от вольф канн – мюллеров канн, функц только в кач яйцевода. Выд-е ч/з вольфов канал.
Рептилии, птицы: у самцов обр вторичн почка, вторичн мочеточник, кот вп в остатки вольфова канала, вольф канал превращ в семепровод, в нижн отд происх соед-е – прод выд-я и пол прод ч/з 1 канал. Самки: вторичн мочеточник откр сам-но. Половой канал – мюллеров канал (яйцевод)
Млекопит: не им клоаки, хорошо развиты первичн пол органы.
Эволюция мочепол сист сводится к:
-смена 3х поколений поек: головн – туловищн – таз
-происх разделение каналов, лужащих для вывед-я продуктов
-усложн свзь кровеносн и выделит сист (формир-е сальп телец, затем нефронов)
Пол сист у позв связ с выд происхождение, положением и разделением канальцев.
Гомеостаз - это способность сохранять устойчивость и постоянство внутренней среды организма. За поддержание гомеостаза отвечают вегетативная нервная система и эндокринная система, которая контролируется гипоталамусом, а последний, в свою очередь, корой головного мозга. Существует два механизма гомеостаза, отрицательная обратная связь и положительная обратная связь. Первая меняет реакцию системы на противоположную, вторая приводит к дестабилизирующему эффекту. В качестве примера можно привести сохранение постоянной температуры тела, сохранение определенного числа особей внутри сообщества, повышение и понижение кровяного давления при недостатке кислорода. У животных защитные функции выполняют перья, мех, большая масса тела. Те, кто не способен поддерживать активность в холодное время, впадают в спячку. Экзотермные животные имеют приспособления, помогающие регулировать температуру тела благодаря внешним источникам тепла. Изотермия свойственна только так называемым гомойотермным, или теплокровным, животным. Волосяной покров и жировая подкожная клетчатка предохраняют тело от охлаждения, а от перегрева оно защищено благодаря испарению влаги через потовые железы.
У теплокровных животных дыхат сист участвует в процессах терморегуляции. Терморегуляция проявляется в форме взаимосочетания процессов теплообразования и теплоотдачи, регулируемых нервно-эндокринным путем. Терморегуляцию принято разделять на химическую и физическую. Химическая терморегуляция осуществляется путем изменения уровня теплообразования, т. е. усиления или ослабления интенсивности обмена веществ в клетках организма. Физическая терморегуляция осуществляется путем изменения интенсивности отдачи тепла. Наиболее интенсивное теплообразование в организме происходит в мышцах. Даже если человек лежит неподвижно, но с напряженной мускулатурой, окислительные нродес-сы, а вместе с тем и теплообразование повышаются на 10%. Небольшая двигательная активность ведет к увеличению теплообразования на 50—80%, а тяжелая мышечная работа — на 400—500%. В условиях холода теплообразование в мышцах увеличивается, даже если человек находится в неподвижном состоянии. Это обусловлено тем, что охлаждение поверхности тела, действуя на рецепторы, воспринимающие холодовое раздражение, рефлекторно возбуждает беспорядочные непроизвольные сокращения мышц, проявляющиеся в виде дрожи (озноб). При этом обменные процессы организма значительно усиливаются, увеличивается потребление кислорода и углеводов мышечной тканью, что и влечет за собой повышение теплообразования. Температура крови печеночной вены выше температуры крови печеночной артерии, что указывает на интенсивное теплообразование в этом органе. При охлаждении тела теплопродукция в печени возрастает. Освобождение энергии в организме совершается за счет окислительного распада белков, жиров и углеводов. Поэтому все механизмы, которые регулируют окислительные процессы, регулируют и теплообразование. Теплоотдача осуществляется путем теплоизлучения (радиационная теплоотдача), конвекции, т. е. движения и перемешивания нагреваемого телом воздуха, теплопроведе-ния, т. е. отдачи тепла веществам, непосредственно соприкасающимся с поверхностью тела, /И испарения воды с поверхности кожи и легких. У человека в обычных условиях потеря тепла путем теплопроведения имеет небольшое/значение, так как воздух и одежда являются плохими проводниками тепла. Радиация, испарение и конвекция протекают с различной интенсивностью в зависимости от температуры окружающей среды. У человека в состоянии покоя при температуре воздуха ' около 20 °С и суммарной теплоотдаче, равной 419 кДж (100 ккал) в час, радиация составляет 66%, испарение воды —19%, конвекция -15% общей потери тепла организмом.
Выделительная система ланцетника – нефридиального типа. По строению близка к выделительной системе некоторых кольчатых червей. Нефридии у ланцетников – короткие изогнутые трубки (до 90 пар), расположенные в области глотки. Большая часть нефридия лежит в целоме. Имеется ряд отверстий (нефростомы), окруженных булавовидными клетками (соленоцитами). Соленоцит имеет тонкий каналец с мерцательным волоском, обеспечивающим ток целомической жидкости. Выводящий конец нефридия открывается в атриальную полость. Продукты метаболизма поступают в полость целома из капилляров, пронизывающих стенки целома, а затем за счет движения мерцательных волосков соленоцитов выводятся в атриальную полость и через атриопор – наружу.
Органы выделения у взрослых рыб представлены парными туловищными почками, парными мочеточниками, мочевым пузырем и мочевым отверстием. Конечными продуктами азотного обмена являются: аммиак, мочевина, триметиламиноксид (это вещество придает мясу рыб специфический запах).
Структурно-функциональная единица почки - нефрон. Проксимальная (начальная) часть нефрона - мальпигиево тельце, сост из бокалообразной почечной капсулы (капсула Боумена–Шумлянского) и расположенного в ее полости клубочка капилляров. Капилляры отделены от полости почечной капсулы особыми клетками (подоцитами). Полость капсулы Боумена–Шумлянского переходит в извитой каналец (который у рыб еще не дифференцирован на проксимальный и дистальный отделы, разделенные изгибом – петлей Генле). Из капилляров почечного клубочка в полость капсулы фильтруется жидкость, содержащая воду, углеводы, аминокислоты и различные ионы - первичная моча. В проксимальном отделе извитого канальца происходит обратное всасывание органических веществ, в дистальном отделе – всасывание воды. В дистальном отделе извитого канальца формируется вторичная моча с повышенным содержанием вредных для организма веществ. Извитые канальцы впадают в парные мочеточники, которые впадают в мочевой пузырь. В мочевом пузыре происходит дополнительное всасывание воды.
У пресноводных рыб возникает проблема выведения избытка воды и сохранения минеральных солей. Морские рыбы, наоборот, вынуждены экономить воду и выводить избыток солей. В регуляции водно-солевого обмена у рыб значительную роль играют жабры и кишечник. У проходных рыб происходит перестройка водно-солевого обмена при переходе из моря в реки и наоборот.
Выделительная и половая система Амфибий тесно связаны между собой и образуют мочеполовую систему, типичную для анамний. Основой выделительной системы являются туловищные почки. Основой половой системы у самок являются парные яичники, а у самцов – парные семенники.
Выделительная система. Парные компактные туловищные почки расположены по бокам позвоночного столба в задней части полости тела. У переднего края каждой почки располагаются жировые тела – резерв питательных веществ для формирования половых клеток. Структурно-функциональной единицей почек является нефрон (его строение и функционирование принципиально не отличается от такового у рыб). Основным продуктом азотного обмена у лягушки является мочевина, а у головастиков и Хвостатых амфибий, живущих в воде – аммиак. По внешнему краю каждой почки тянется тонкий канал – мочеточник (вольфов канал), открывающийся в клоаку. В клоаку открывается и мочевой пузырь. Моча накапливается в мочевом пузыре, концентрируется а затем выводится наружу.
Выделительная система рептилий – типичная для амниот. Органы выделения представлены тазовыми почками. Особенностью нефронов тазовых почек является полное отсутствие связи с полостью тела, в результате полостная жидкость не выводится наружу. Имеются парные мочеточники, не гомологичные вольфовым каналам.
У крокодилов и пресноводных черепах выделяется жидкая моча, у наземных Рептилий – твердая или кашицеобразная. Основным продуктом азотного обмена является мочевая кислота. Особенности выделительной системы настолько обеспечивают независимость рептилий от воды, что позволили им захватить совершенно засушливые области Земли.
Выделительная система птиц – типичная для амниот. Почки компактные, сегментированные, заполняют углубления подвздошных костей. У большинства видов имеется петля Генле – тонкая вставка между проксимальной и дистальной частями извитого канальца. Петля Генле обеспечивает эффективное всасывание воды. Основным продуктом азотного обмена, как и у Рептилий, является мочевая кислота. Мочевого пузыря нет. Есть клоака.
У Млекопитающих, как и у всех амниот, имеются тазовые почки. Кровь поступает в почки по почечным артериям из спинной аорты. После очистки кровь возвращается в заднюю полую вену по почечным венам.
Человек:На разрезе почки выделяют два слоя: корковый (наружный) и мозговой (внутренний). Корковое вещество содержит нефроны (в состав одной почки входит свыше миллиона нефронов). Мозговое вещество состоит из собирательных трубочек и кровеносных сосудов, собранных вместе в виде почечных пирамид. Верхушки пирамид, называемые сосочками, открываются в малые почечные чашечки. Малые почечные чашечки открываются в большие почечные чашечки, из которых моча поступает в почечную лоханку. Почечная лоханка образует расширенное устье мочеточника. Мочеточники дают начало мочевыводящим органам: они выходят из почки и впадают в мочевой пузырь. Мочевой пузырь через мочеиспускательный канал открывается наружу мочеиспускательным отверстием.
Структурно-функциональной единицей почки является нефрон. Нефрон состоит из почечной капсулы (капсулы Боумена–Шумлянского) и почечного канальца. Капсула имеет вид двуслойного бокала. Внутренний слой капсулы образован специализированными клетками – подоцитами. Между слоями капсулы имеется полость. Внутри капсулы располагается клубочек капилляров (мальпигиев клубочек). Почечная капсула и мальпигиев клубочек образуют почечное тельце.
От почечной капсулы отходит извитой каналец, в состав которого входят: проксимальная часть (каналец I порядка), петля Генле и дистальная часть (каналец II порядка). Стенки извитых канальцев образованы кубическим эпителием, который имеет щетковидную кайму – мельчайшие выросты, увеличивающие всасывающую поверхность. Почечная капсула, канальцы I и II порядка расположены в корковом слое, а петля Генле опускается в мозговой слой.
В капсулу каждого нефрона входит приносящая артериола (ответвление почечной артерии) и распадается на капилляры, образующие мальпигиев клубочек. Капилляры собираются в выносящую артериолу, диаметр которой в 2 раза меньше диаметра приносящей артериолы. Разность в диаметрах приносящей и выносящей артериол обусловливает высокое давление в капиллярах мальпигиева клубочка – 70...90 мм рт. ст. Выносящая артериола вновь распадается на капилляры, оплетающие извитые канальцы и петлю Генле. После этого капилляры образуют вены, впадающие в почечную вену.
Канальцы II порядка от нескольких нефронов впадают в собирательную трубочку; собирательные трубочки объединяются в более крупные выводные протоки. Выводные протоки проходят через мозговое вещество и открываются на верхушках пирамид через почечный сосочек в малую почечную чашечку. Через малые и большие почечные чашечки, почечную лоханку и мочеточники моча поступает в мочевой пузырь.
В почечных тельцах происходит ультрафильтрация: за счет высокого давления в капиллярах мальпигиева клубочка в почечную капсулу поступает водный раствор всех веществ с молекулярной массой до 68 тысяч а. е. м. (минеральные соли, мочевина, мочевая кислота, пигменты, глюкоза, аминокислоты, витамины, гормоны). Белки в норме через стенки капилляров и почечной капсулы не проходят, однако при заболевании почек, при напряженной мышечной работе в моче обнаруживается белок. В результате фильтрации в полости капсулы образуется клубочковый фильтрат – первичная моча. За сутки образуется до 150 литров первичной мочи.
В проксимальном отделе извитого канальца (в канальцах I порядка) происходит реабсорбция (обратное всасывание) более 80% веществ: все органические вещества, около 50% мочевины; около 85% NaCl и воды. Реабсорбция через эпителий почечных канальцев протекает с затратой энергии АТФ. Образуется фильтрат, изотонический по отношению к плазме крови. В этот фильтрат секретируются некоторые вещества: креатин, органические кислоты, ионы натрия, калия, водорода, лекарственные вещества.
На протяжении петли Генле создается и поддерживается продольный градиент осмотического давления в мозговом веществе по направлению от коркового слоя к сосочкам, то есть осмотическое давление в эпителиальных клетках возрастает. Поэтому эпителий петли Генле постоянно всасывает воду, и благодаря этому образуется гипертоническая моча.
В дистальном отделе извитого канальца (в канальцах II порядка) происходит тонкая регуляция водно-солевого баланса и рН крови. В результате образуется вторичная моча, содержащая 95% воды, мочевину, мочевую кислоту, креатин, соли калия, натрия и другие ионы. Вторичная моча отличается от первичной отсутствием сахара, аминокислот, повышенной концентрацией мочевины (≈ в 70 раз). За сутки образуется около 1,5 литров вторичной мочи.
Вторичная моча поступает в собирательные трубочки. Проницаемость стенок собирательных трубочек регулируется антидиуретическим гормоном. Собирательная трубочка вместе с дистальным отделом извитого канальца участвует в образовании гипертонической или гипотонической мочи (в зависимости от потребности организма в воде).
Таким образом, окончательный состав мочи определяется тремя почечными процессами: фильтрацией, реабсорбцией и секрецией.
24.Сравнительная характеристика кровеносной системы в ряду позвоночных. Сердце. Круги кровообращения. Сосуды и их преобразования.
У позвоночных – замкнутая. Кровь циркулирует только по сосудам, центр орган – сердце, м состоять из 2х камер (личинки костных рыб, личинки амфибий), 3х камер (амфибии, рептилии), 4 камеры (птицы, млекопитающие, крокодилы). Предсердия принимают кровь, желудочки – камеры, кот выталкивают кровь из сердца (опр напр-е тока крови). Кров сосуды представлены венами (несут кровь к сердцу), артериями (несут кровь от сердца), капиллярами (соединяют венозн и арт части кров сист, обр «чудесные» сети.
У низших позвоночных (рыбы, круглоротые, личинки амфибий) – 1 круг кровообращеня, сердце сост из желудочка и предсердия, в сердце только венозная кровь.
У наземных позвоночных в связи с появлением лёгких формир 2 круга кровообра – большой (несёт кровь от сердца ко всем внутр органам, кроме лёгких) и малый (лёгочный – несёт кровь к лёгким).
Круглоротые: сердце из предс и желудочка, к предс прилег венозный синус. От брюшной аорты – приносящ жаберн артерии (прох в жаб перегородках) – выносящ жаб арт – корни аорты – (вперёд: сонные артерии несут арт кровь в голову, назад – спинная аорта, расп над хордой). Венозн сист: от головы венозн кр собирается в парные кардинальные вены – впадают в венозны синус. От туловища кр соб в задн кард вены – вп в венозн синус. От кишечника соб в подкиечн вену, вх в печень (воротная система печени – расщепл всех яд вещ-в) – печеночная вена – вп в венозе синус. Венозн синус – кр в сердце.
Воротн сист почек нет.
3 дополнит сердца: в голове, в печени, в хвосте, имеются мышцы.
Хрящевые рыбы: сердце венозное, двухкамерное. Имеется артериальный конус, отходящ от желудочка (обособл участок желудочка), в нём имеются парные клапаны. Стенки желудочка способны к ритмич сокращениям. 1 круг кровообращения.
Арт сист: от арт конуса берёт начало брюшная аорта – к жабрам 5 пар жаб артерий – корни аорты – (спереди: сонные артерии, сзади – спинная аорта – многочисл артерии к внутр органам). Венозн сист: от головы кровь собирается в парныя ярёмные вены. От туловище – в задние кардинаяльные вены. От пер яр вен и задн кард – вблии сердца кювьеровы протоки – венозн синус. Кардинальные вены образуют воротную систему почек, имеется воротная система печени.
Костные рыбы: у большинства нет артериального конуса, брюшная аорта в начале имеет вздутие артериальная луковица, жаберных артерий – 4 пары (по числу жабер), прав кард вена непрерывна, левая образует воротную систему левой почки. Спереди в головном отделе сосуды сливаются.
Амфибии (земноводные): сердце трёхкамерное. Венозе пазуха, 2 предсердия, 1 желудочек, артериальный ствол. От общего артериального конуса до общего артериального ствола отх 3 пары арт дуг: 1я пара – сонные артерии (отпр арт кровь в голову), 2я пара – дуги аорты (смешанная кровь к органам тела), 3я пара – кожно-лёгочные артерии (несут венозную кровь к органам дыхания – лёгким и коже). Оба предсердия открываются в желудочек одним общим отверсием. Кров сист амф изм в связи с заменой жаб дыхания на кожно-лёгочное. Внутр пов-ть желудочка имеет углубления – кровь в нём не полностью успевает смешаться. В прав половине сердца содержится венозная кровь (поступ по полым венам), средн ч сердца сод смеш кровь, лев ч сод арт кровь (т.к. кожно-лег вены мал кр кровообр вп в лев предс).
У бесхвостых амфибий: 1я и 2я пары жаб артерий рыб редуцируются, 3я пара преобр в сонные артерии, 4я пара даёт дуги аорты – все соед-ся в спинную аорту. 5я пара исчезает за искл хвостат амф. 6я пара жаб арт рыб преобразуется в кожно-лёгочн арт.
Венозе сст близка к сист двоякодышащих рыб, м 2 воротн сист почек, кювьеровы протоки, сохр появл кожно-лёгочн вены (нес кр в прав предс), сохр протоки, кот связ кожно-лёгочн арт и дуги аорты – баталовы протоки.
Рептилии: сердце трёхкамерное, в желудочке появл неполная перегородка (у крокодила пер нет), нет общего арт ствола. Все 3 пары сосудов самот отх от сердца (справа – лёгочный сосуд, слева – прав дуга аорты, от сер желуд – левая дуга аорты). Слева – арт кр, спр – венозн, в сер – смеш. Арт комп-ты в смеш крови больше (70%). Происх дальнейшее разделение артерий и вен на более мелкие – появл доп сосуды.
Птицы: сердце 4хкам, перегородка в желудочке полная. Редуцируется полность левая дуга аорты. Большой круг кровообращения начинается не дугами аорты, а правой дугой аорты. Арт кровь снабжает все внутр органы, пост темп тела, теплокровность, от пра дуги аорты отделяются безымянные артерии, от спинной аорты ответвл артерии к внутр орг., подключичн арт раздел-ся на сонные (идут к голове), подключичные (идут к крыльям) и грудные (несут кровь к грудным мышцам).
Венозн сист: по небольш хвост венам кр изливается в сеть воротной ситемы почек, часть крови проходит ч/з в-во почек по особ сосудам, выходит из почек напрямую, слив с бедренными венами, обр подвздошные вены, кот вливаются в нижнюю полую вену. От внутр органов кровь собирается в копчико-брыжеечную вену. Брыжейка – мезодермальная выстилка целома. От брюшн ст тела кр поступ в надключичн вену. Все вены сливаются в воротн сит печени. Печент – печеночн вена – нижн пол вена – прав предсердие (венозн кровь)
Малый круг: нач общим стволом от правого желудочка. Лёгочн арт – прав, лев (несут венозн кровь к лёгким, газообмен) – легочные вены (арт кровь в левое предсердие).
Млекопитающие: сердце 4х кам с лев дугой аорты. Остатки кардинальных вен – непарные вены. Правая непарая вена самост впадает в переднюю полую вену, левая непарная вена утрач связь с полой веной, вп в прав непарн вену ч/з особ поперечн сосуд – поперечная вена. Нет воротной системы почек.
25.Особенности развития животных. Анамнии и амниоты. Гаструляция. Гисто- и органогенез. Типы дробления. Появление яйцеживорождения и живорождения.
Анамнии связаны с водой. Либо вся жизнь, либо половое размножение. Имеются редкие исключения носящие вторичный характер. Земноводные первичноводные, предки чисто водные. Все рыбы и многие земноводные всю жизнь проводят в воде. Но есть искл. живущие во влажном воздухе. Но для размножения нужна вода. Яйца бедны желтком, оплодотворение в воде наружное. У хрящевых рыб и некот земноводных внутреннее. Личинка ведет водный образ жизни. После метаморфоза может жить вне воды. Амниоты на суше. Но есть и живущие в воде, это вторичный переход. Различия между этими 2 группами проявляются в строение яйца, характере эмбрионального развития и строения взрослых особей. Яйцо. У анамний имеет слизистую оболочку, (сохранение формы в воде), желтка мало. В яйцах амниот кол-во желтка больше, содержит все в-ва для развития. Имеется большая белковая оболочка( много воды). Образуется наружная оболочка – плотная, сохраняет форму в воздушной среде. У пресмыкающихся имеются волокна рогоподобного веществава(вид пергамента). М/у волокнами кристаллики извести. У птиц скорлупа имеет извитые канальцы(поступление О2 и роль фильтра). Эмбрион. У анамний полное не равномерное дробление, личинка дышит жабрами, метаморфоз (обр. конечности и легочное дыхание, перестраивается кровеносная система). Связано с переходом к наземному образу жизни. У амниот яйцо перегружено желтком. Дробление на анимальном полюсе однослойный зародышевый диск первичная зародышевая бороздка часть клеток под него(энто и мезодерма) сомиты все др. органы. Образуется полая кольцевая складка внутри нее находится полость, она окружает зародыш и края срастаются. Амниотическая полость защищает зародыш от пересыхания. появляется аллантоис – мочевой пузырь. Его наружная стенка примыкает к серозной оболочке, имеет капиллярную сеть. у амниот производные кожи волосы и перья –теплоизолирующая ф-ция. У пресмыкающихся по с равнению с земноводными разобщенный малый и большой круг кровообращения, а у птиц и млекопитающих они разделены. Характерные для анамний мезентерические почки, функционируют только у зародыша амниот. У амниот больший размер головного мозга. У пресмыкающихся и птиц больше содержание нервных клеток, а у млекопитающих появляется кора больших полушарий.
Гаструляция — сложный процесс морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки (эктодерма, мезодерма и энтодерма) — источники зачатков тканей и органов. Второй после дробления этап онтогенеза. При гаструляции происходит перемещение клеточных масс с образованием из бластулы двухслойного или трёхслойного зародыша — гаструлы.
Тип бластулы определяет способ гаструляции.
Зародыш на этой стадии состоит из явно разделенных пластов клеток - зародышевых листков: наружного (эктодерма) и внутреннего (энтодерма).
У многоклеточных животных, кроме кишечнополостных, параллельно с гаструляцией или, как у ланцетника, вслед за ней возникает и третий зародышевый листок - мезодерма, который представляет собой совокупность клеточных элементов, расположенных между эктодермой и эндодермой. Вследствие появления мезодермы зародыш становится трехслойным.
На этой стадии начинается использование генетической информации клеток зародыша, появляются первые признаки дифференцировки.
Дифференцировка (дифференцирование) - процесс возникновения и нарастания структурных и функциональных различий между отдельными клетками и частями зародыша.
Образуются несколько сотен типов клеток специфичного строения, отличающихся друг от друга. Синтез определенных белков, свойственных только данному типу клеток.Биохимическая специализация клеток обеспечивается дифференциальной активностью генов.
Из эктодермы образуется нервная система, органы чувств, эпителий кожи, эмаль зубов; из энтодермы - эпителий средней кишки, пищеварительные железы, эпителий жабр и легких; из мезодермы - мышечная ткань, соединительная ткань, кровеносная система, почки, половые железы и др.
У разных типов животных одни и те же зародышевые листки дают начало одним и тем же органам и тканям.
Способы гаструляции
Инвагинация — происходит путем впячивания стенки бластулы в бластоцель; характерна для большинства групп животных.
Деляминация (характерна для кишечнополостных) — клетки, находящиеся снаружи, преобразуются в эпителиальный пласт эктодермы, а из оставшихся клеток формируется энтодерма. Обычно деляминация сопровождается делениями клеток бластулы, плоскость которых проходит «по касательной» к поверхности.
Иммиграция — миграция отдельных клеток стенки бластулы внутрь бластоцеля.
Униполярная — на одном участке стенки бластулы, обычно на вегетативном полюсе;
Мультиполярная — на нескольких участках стенки бластулы.
Эпиболия — обрастание одних клеток быстро делящимися другими клетками или обрастание клетками внутренней массы желтка (при неполном дроблении).
Инволюция — вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток.
Дробление — это ряд последовательных митотических делений зиготы и далее бластомеров, заканчивающихся образованием многоклеточного зародыша — бластулы. Первое деление дробления начинается после объединения наследственного материала пронуклеусов и образования общей метафазной пластинки. Возникающие при дроблении клетки называют бластомерами (от греч. бласте—росток, зачаток). Особенностью митотических делений дробления является то, что с каждым делением клетки становятся все мельче и мельче, пока не достигнут обычного для соматических клеток соотношения объемов ядра и цитоплазмы. У морского ежа, например, для этого требуется шесть делений и зародыш состоит из 64 клеток. Между очередными делениями не происходит роста клеток, но обязательно синтезируется ДНК.
Все предшественники ДНК и необходимые ферменты накоплены в процессе овогенеза. В результате митотические циклы укорочены и деления следуют друг за другом значительно быстрее, чем в обычных соматических клетках. Сначала бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость — бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, образуя стенку бластулы — бластодерму. Общий размер зародыша к концу дробления на стадии бластулы не превышает размера зиготы.
Главным результатом периода дробления является превращение зиготы в многоклеточный односменный зародыш.
Морфология дробления. Как правило, бластомеры располагаются в строгом порядке друг относительно друга и полярной оси яйца. Порядок, или способ, дробления зависит от количества, плотности и характера распределения желтка в яйце. По правилам Сакса — Гертвига клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы, а веретено клеточного деления — в направлении наибольшей протяженности этой зоны.
В олиго- и мезолецитальных яйцах дробление полное, или голобластическое. Такой тип дробления встречается у миног, некоторых рыб, всех амфибий, а также у сумчатых и плацентарных млекопитающих. При полном дроблении плоскость первого деления соответствует плоскости двусторонней симметрии. Плоскость второго деления проходит перпендикулярно плоскости первого. Обе борозды первых двух делений меридианные, т.е. начинаются на анимальном полюсе и распространяются к вегетативному полюсу. Яйцевая клетка оказывается разделенной на четыре более или менее равных по размеру бластомера. Плоскость третьего деления проходит перпендикулярно первым двум в широтном направлении. После этого в мезолецитальных яйцах на стадии восьми бластомеров проявляется неравномерность дробления. На анимальном полюсе четыре более мелких бластомера — микромеры, на вегетативном — четыре более крупных — макромеры. Затем деление опять идет в меридианных плоскостях, а потом опять в широтных.
В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или мероб-ластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным.
При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным. Оно типично для хордовых и иглокожих. В природе встречаются и другие варианты пространственного расположения бластомеров при дроблении, что определяет такие его типы, как спиральное у моллюсков, билатеральное у аскариды, анархичное у медузы.
Замечена зависимость между распределением желтка и степенью синхронности деления анимальных и вегетативных бластомеров. В олиголецитальных яйцах иглокожих дробление почти синхронное, в мезолецитальных яйцевых клетках синхронность нарушена после третьего деления, так как вегетативные бластомеры из-за большого количества желтка делятся медленнее. У форм с частичным дроблением деления с самого начала асинхронны и бластомеры, занимающие центральное положение, делятся быстрее.
26.Особенности организации, классификация и происхождение птиц.
Палеонтологические материалы по птицам очень скудны и отрывочны. Поэтому говорить о предках птиц среди рептилий и о происхождении современных групп птиц приходится преимущественно предположительно, основываясь на морфологических особенностях ныне живущих групп и на отрывочных палеонтологических данных.
По современным представлениям, предками птиц были архозавры (Archosauria) - обширная и очень разнообразная группа рептилий, включавшая различных динозавров, крокодилов, летающих ящеров. Непосредственных предков птиц, видимо, нужно искать среди наиболее древних и примитивных архозавров - текодонтов, или псевдозухий (Thecodontia, или Pseudosuchia), давших начало и остальным архозаврам. Они были похожи на ящериц, вели наземный образ жизни и при необходимости передвигались на задних конечностях. По строению черепа, таза, задних конечностей псевдозухий весьма сходны с птицами. Полагают, что переход к древесному образу жизни и приспособление к прыжкам с ветки на ветку сопровождался разрастанием и усложнением роговых чешуи на боках туловища, по краям конечностей и хвоста, что обеспечивало возможность планирования и удлинения прыжков.
Дальнейшая специализация и отбор привели к развитию оперения крыла, обеспечившего возможность сначала планирующего, а затем активного полета. Оперение всего тела птиц, вероятно, развивалось не столько как приспособление к полету (обтекаемость тела), сколько как приспособление к термоизоляции.
Обособление птиц от пресмыкающихся, видимо, произошло в конце триаса - начале юры (190-170 млн. лет назад). Однако ископаемых остатков птиц этого времени пока не найдено. В настоящее время палеонтологами изучены остатки только двух самых древних известных нам птиц - Archaeopteryx lithographica и Archaeornis simensi, найденные в Баварии в сланцевых песчаниках (возраст около 150 млн. лет). Судя по внешнему облику к ряду'внутренних признаков, несмотря на сходство с пресмыкающимися, многие исследователи считают, что архептерикс и археорнис - древние, примитивные, но все же птицы.
Птицы подкласса настоящих, или веерохвостых, птиц (Neomithes), видимо, являются потомками каких-то ящерохвостых пткц (Archaeornithes), но не обязательно археоптерикса и археорниса. Происхождение же современных отрядов птиц остаются очень гипотетическими.
Ископаемые остатки веерохвостых птиц - гесперорнисов обнаружены только в меловом периоде (примерно 90-80 млн. лет). Эти крупные птицы, похожие на гагар или поганок, имели недоразвитые передние конечности и поэтому не летали, но хорошо плавали и ныряли. Челюсти их несли зубы.
Другие зубастые птицы меловых отложений - ихтиорнисы - были примерно с голубя, имели крылья, киль на грудине и, видимо, обладали активным полетом.
Судя по более поздним материалам, можно предполагать, что интенсивная адаптивная радиация птиц и становление большинства современных отрядов проходило в самом конце мелового периода мезозойской эры - в третичный период кайнозойской эры (40-70 млн. лет назад).
Единого взгляда исследователей на ход эволюции класса птиц до настоящего времени не выработано.
Подкласс Древние
Отряд 1 Археоптериксы
Подкласс Настоящие птицы
Надотряд 1 Зубастые птицы
Отряд 2 Гесперорнисообразные
Надотряд 2 Ихтиорнисы
Отряд 3 Ихтиорнисообразные
Надотряд 3 Плавающие
Отряд 4 Пингвинообразные
Надотряд 4 Типичные птицы
Отряд 5 Страусообразные
Отряд 6 Нандуобразные
Отряд 7 Казуарообразные
Отряд 8 Эпиорнисообразные
Отряд 9 Маообразные
Отряд 10 Кивиобразные
Отряд 11 Тинамуобразные
Отряд 12 Гагарообразные
Отряд 13 Поганкообразные
Отряд 14 Трубконосообразные
Отряд 15 Пеликанообразные
Отряд 16 Аистообразные
Отряд 17 Гусеобразные
Отряд 18 Соколообразные
Отряд 19 Курообразные
Отряд 20 Журавлеобразные
Отряд 21 Диатримообразные
Отряд 22 Ржанкообразные
Отряд 23 Голубеобразные
Отряд 24 Попугаеобразные
Отряд 25 Кукушкообразные
Отряд 26 Совообразные
Отряд 27 Козодоеобразные
Отряд 28 Стрижеобразные
Отряд 29 Птицы-мыши
Отряд 30 Трогонообразные
Отряд 31 Ракшеобразные
Отряд 32 Дятлообразные
Отряд 33 ВоробьинообразныеВероятно, эволюция в классе птиц шла по двум направлениям - от древних птиц к ветви наземных и водных птиц и ветви древесных птиц. От первой ветви отделились тинаму и бескилевые птицы, включая всех ныне живущих страусоподобных. Далее обособляются куриные птицы и самостоятельной ветвью - гагарообразные (включающие ископаемых меловых зубастых птиц гесперорнисов, гагар и поганок). Тесно связаны друг с другом и несколько противопоставляются другим группам отряды журавлеобразных (пастушки, настоящие журавли, дрофы и др.) и болотно- и чайкообразных птиц (кулики, чайки, чистики, а из ископаемых -ихтиорнисы). Далее обособляются отряды гусеобразных, фламинго, голенастых, дневных хищников и веслоногих. От форм, близких к предкам веслоногих, отделяется ветвь, которая затем разделяется на отряды трубконосых и пингвинов.
Развитие второй ветви - древесных птиц, начинается от каких-то предковых форм, видимо, более древних, чем предки первой ветви. Сначала общим стволом обособляются 3 отряда: кукушкообразных, голубей (голуби, рябки и дронты) и попугаев. Следующая обособившаяся ветвь разделяется на 2 ответвления, каждое из которых образует по 2 отряда. Одно из них дает начало совам и козодоям, другое - трогонам и ракшеобразньш. Дальше отделяются отряды птиц-мышей и стрижеобразных (стрижи и колибри), самостоятельной ветвью отходит отряд дятлов, а основной ствол ветвится на многочисленные подразделения отряда воробьиных.
Особенности дыхательной системы птиц. Гортань птиц (larinx) существенно не отличается от гортани рептилий. Голосовых связок в ней также нет. Голосовой аппарат у птиц, в отличие от всех прочих обладающих голосом животных, находится в месте разветвления трахеи на бронхи. Здесь образуется сложная нижняя гортань (syrinx). Голосовыми связками служат перепончатые стенки бронхов между раздавшимися хрящевыми или костными кольцами. Они вдаются в просвет гортани складками и, напрягаясь, вибрируют от проходящего выдыхаемого воздуха. Кроме того, на месте самого расхождения бронхов внутри, на так называемом козелке, образуется еще голосовая перепонка. Управление голосовыми связками осуществляется особой довольно сложной мускулатурой нижней гортани. Строение нижней гортани у разных птиц бывает очень различным. У некоторых уток, например, здесь образуется большой асимметричный пузырь-резонатор. Различна сложность мускулатуры и строения самих голосовых перепонок (связок). Легкие птиц, в отличие от легких рептилий, полностью расчленены на бронхи и легочные пузырьки-альвеолы и имеют притом своеобразное строение. Бронхи ветвятся очень сложно, главные их ветви выходят из легких в нескольких местах, оканчиваются обширными легочными воздушными мешками: парой шейных с подмышечными, непарным межключичным, двумя - тремя парами грудных и парой брюшных. Мешки располагаются в полости тела и под кожей, а также дают ответвления в некоторые трубчатые кости (в зависимости от" степени заполнения полостей костным мозгом), а также и в некоторые губчатые кости. Между ветвями бронхов имеется множество соединительных трубок-парабронхов, на которых сидят бесчисленные ветвистые легочные альвеолы, обвитые легочными капиллярными кровеносными сосудами. Благодаря такой расчлененности, дыхательная поверхность легких птиц больше, чем у других животных, что соответствует повышенной потребности в кислороде при работе мощного летательного аппарата. Так, у человека дыхательная поверхность легких равна 13 см2 на 1 кг живого веса, у голубя — 173 см2, а у колибри — до 666 см .В полете грудная клетка совершает дыхательные движения: расширяется — вдох, сжимается — выдох. При работе крыльев, в лад их подниманию и опусканию, происходит также расширение и сжатие полости тела. Расширение полости передается не столько самим легким, которые плотно притянуты к спинной части полости, сколько воздушным мешкам, которые собственно и набирают дыхательный воздух при расширении, и продувают его через легкие при сжатии. Понятие «вдох» и «выдох» здесь приобретает несколько иное содержание, чем в случае рептилий и млекопитающих. При поднимании крыльев расширяется грудная клетка и полость тела и осуществляется 1-я фаза вдоха: воздух, проходя по бронхам сквозь легкие, заполняет задние воздушные мешки. Затем (2-я фаза вдоха) воздух при сжатии задних мешков продувается под давлением сквозь всю систему парабронхов и легочных альвеол, где осуществляется процесс дыхания в физиологическом смысле, т.е. газообмен. Отсюда воздух, обогащенный углекислым газом, отсасывается расширяющими передними воздушными мешками (1-я фаза выдоха) и, при очередном сжатии, отработанный воздух выходит в трахею и наружу (2-я фаза выдоха). Работа задних и передних мешков происходит попеременно. Мешки тонкостенные, специальной дыхательной сети кровеносных сосудов не имеют и в газообмене не участвуют. В покое акт дыхания у птиц совершается обычным для наземных позвоночных способом: путем сужения и расширения грудной клетки, что достигается активным подтягиванием грудины к позвоночнику (вспомните строение ребер у птиц!) и последующим опусканием ее при расслаблении мышц. При ходьбе вентиляции легких способствует периодическое давление бедер на брюшную стенку и, следовательно, на задние воздушные мешки.
Следует иметь в виду и побочные функции воздушных мешков, из которых главные:
1) устранение перегрева внутренних органов и мускулатуры при усиленной работе летательного аппарата, что достигается постоянной вентиляцией воздушных мешков);
предохранение внутренних органов от сотрясения и толчков при беге, полете, посадке и т.д. (воздушные мешки отделены от внутренних органов в полости тела системой пленчатых перегородок-диафрагм и образуют упругие прокладки между органами);
уменьшение удельного веса и повышение плавучести водоплавающих.
Основы биохимии.
1.Белки и их функции. Выделение очистка белков (хроматография, электрофорез).
Белками называют полимеры АК, связанные пептидной связью, с числом мономеров более 100. Размеры белка могут существенно варьировать и достичь тысяч АК остатков. Помимо АК в состав белков м входить и др компаненты: ионы Ме, производные витаминов, липидов, углеводов. В природе сущ достаточно большое разнообразие: у чел-ка 200 тыс, в 1 кл – 30-50, всё многообразие – млн.
Классификация беков.
-по сотаву: простые (неконъюгированные, состоят только из АК: альфа, бета-кератин, коллаген, фебоин, эластин), сложные (конъюгированные, дают и небелковые группы при гидролизе(НК, липиды, сахара, ионы Ме, гемм), сложные белки подразделяют на группы: гемопротеины, металлопротеины (ферритин), липопротеины (бета-липопротеин крови), гликопротеины (иммуноглобулин), фосфопротеины (казеин), нуклеопротеины (остаток НК, ибонуклеопротеин), флавопротеины (флавин – производн вит В2),
-по пространственной структуре: фибриллярные (альфа-кератин): линейная молекула – фибрилла, плохо растворимы в воде, вх большое кол-во гидрофобных кислот, часто выполняют структурную функцию, глобулярные (альбумин, гемоглобин): объёмная молекула – глобула, имеют более сложную структуру – гидрофобное ядро и гидрофильную пов-ть.
-по функциям:
1.Белки как ферменты. Фермент – биологический катализатор. Биол катализ происх в живой клетке – ограниченные условия (не м.б. эекстримальными).
2.Транспортная. способны доставлять множество вещ-в: межмембранный транспорт (фермиаза, K/Na-АТФаза, ферритин), транспорт м/у органами внутри организма (альбумин, гемоглобин).
3.Защитная: защита от внешней среды (внешние покровы:рога, копыта, панцирь, шерсть), защита внутри организма (иммунитет, яды).
4.Запасные, или пищевые белки (альбумин – белок яйца, козеин – белок молока).
5.Сократительная и двигательная ф-ция. Мышечн сокращения – актин-миозиновый комплекс. Флагеллин – жгутики, реснички.
6.Структурная ф-ция (альфа, бета-кертин, эластин).
7.Регуляторная ф-ция. Множество белков выступает гармонами, а также выполняет рецепторную ф-цию.
8.Прочие. обеспечивают осмогомеостаз, ьермогенез и др.
Методы выделения и очистки белков.
В основе выделения белков лежат 2 гр принципов: их дифференциальное осаждение и хроматографический метод. Для дифф осаждения исп-ся агенты, разрушающие гидратную оболочку белка. При этом падает его растворимость и он при различной концентрации осадителя выпадает в осадок: высаливание сульфатом аммония, осаждение ацетоном, осаждение спиртами.
В основе хроматографии лежит разделение молекул по их сродству к твёрдому носителю и растворителю (Цвет): ионообменная хр-я, гель-хр-я, аффинная хр-я. При ионообменной хр-ии к носителю привязываются функциональные группы, кот имеют заряд. Наиб часто исп – карбокси. Диэтиламиноэтилсефадекс. На колонку наносится смесь белков и те из них, кот заряжены максимально противоположно заряду колонки сорбируются макс эффективно. Те, у кого заряда нет, или он такого же знака, с колонки элюируется сразу. Для десорбции исп р-р с высокой ионной силой.
При гель-хр-ии разделение белков идёт по массе, что достигается применением пористого носителя. Мелкие молекулы, попадая внутрь этих пор, элюируются значительно медленнее, чем крупные.
Аффинная хр-я основана на разделении вещ-в по сродсту к определённому лиганду (либо аналог субстрату, либо антитело) самый точный, быстрый эффективный способ. Для анализа эфф-ти выдел-я белков и белкового полиморфизма исп-ся электрофорез, позволяющий получить данные о том, какие белки и сколько содержатся в данной пробе.
Электрофорез бывает 2х типов: нативный (неразрушающий) и денатурирующий. При ативном белки не разрушаются и идёт их разделение как по заряду, так и по размеру молекул. При денатуррующем электрофорезе в пробу добавляется детергент (додецилсульфат натрия), который разворачивает белковую молекулу. Вся молекула становится линейной, облепленной молекулами детергента. Разделение белков идёт по их размеру. Чем больше мол-ла, тем медленней она разделяется.
5.Гликолиз – центральный путь катаболизма глюкозы.
Катаболизм – фаза метаболизма, на которой происходит распад молекул, поступающих с пищей или запасенных живым организмом до неорганических веществ (H2O, CO2, NH4). В ходе катаболических процессов высвободившаяся при окислении энергия запасается в форме АТР, NAD(P)H и мембранного потенциала. на первой стадии происходит активация субстратов за счет фосфорилирования (затрачивается 2 АТР), включение в данный процесс других сахаров, распад на две С3 молекулы (глицеральдегид-3-фосфат и дигидроксиацетонфосфат).На втором этапе происходит окисление глицеральдегид-3-фосфата до пирувата (в аэробных условиях) или до лактата (при анаэробиозе) с образованием 4-х молекул АТР.
Глюкоза гексокиназа Глюкозо-6-фосфат фосфоглюкоизомераза Фруктозо-6-фосфат фосфофруктокиназа Фруктозо-1,6-бисфосфат альдолаза Глицеральдегид-3- фосфат Дигидроксиацетонфосфат триозофосфатизомераза
Запасание энергии
Глицеральдегид-3- фосфат
глицеральдегид-3-фосфатдегидрогеназа
(NAD+ NADH)
1,3-дифосфоглицерат
фосфоглицераткиназа (АДР АТР)
3-фосфоглицерат
фосфоглицератмутаза
2-фосфоглицерат енолаза фосфоенолпируват пируваткиназа (АДР АТР) пируват
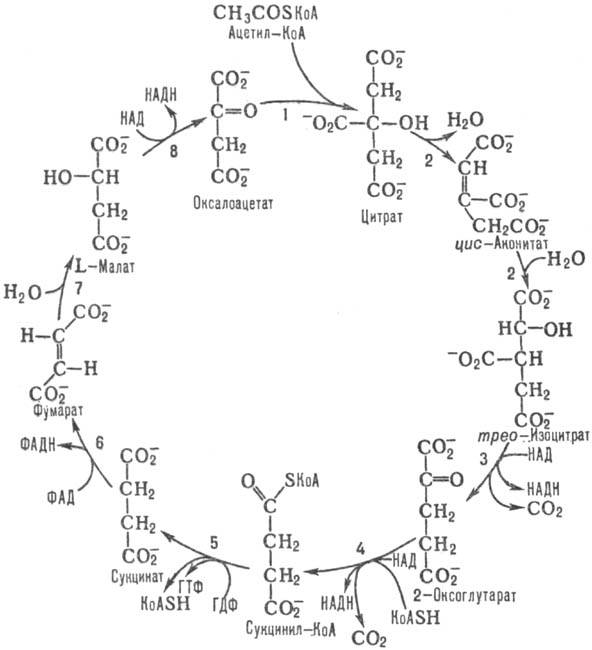
6.Цикл трикарбоновых кислот.
Цикл трикарбоновых кислот (цикл Кребса, цитратный цикл) — центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ.Цикл Кребса — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др.Цикл превращения лимонной кислоты в живых клетках был открыт и изучен немецким биохимиком Хансом Кребсом, за эту свою работу он (совместно с Ф. Липманом) был удостоен Нобелевской премии (1953 год).У эукариот все реакции цикла Кребса протекают внутри митохондрий, причём катализирующие их ферменты, кроме одного, находятся в свободном состоянии в митохондриальном матриксе, исключение составляет сукцинатдегидрогеназа, которая локализуется на внутренней митохондриальной мембране, встраиваясь в липидный бислой. У прокариот реакции цикла протекают в цитоплазме.При работе цикла Кребса окисляются различные продукты обмена, в частности токсичные недоокисленные продукты распада алкоголя, поэтому стимуляцию цикла Кребса можно рассматривать как меру биохимической детоксикации.
7.Уровни структурной организации белков.
Белки́ (протеи́ны, полипепти́ды) - высокомолекулярные органические вещества, состоящие из соединённых в цепочку пептидной связью альфа-АКт. В живых организмах аминокислотный состав белков определяется генетическим кодом, при синтезе в большинстве случаев используется 20 стандартных АКт. Множество их комбинаций дают большое разнообразие свойств молекул белков. Кроме того, АКты в составе белка часто подвергаются посттрансляционным модификациям, которые могут возникать и до того, как белок начинает выполнять свою функцию, и во время его «работы» в клетке. Часто в живых организмах несколько молекул белков образуют сложные комплексы, например, фотосинтетический комплекс. Первичная структура — последовательность АК в полипептидной цепи. Важными особенностями I являются консервативные мотивы — сочетания АКт, играющих ключевую роль в функциях белка. Консервативные мотивы сохраняются в процессе эволюции видов, по ним часто удаётся предсказать функцию неизвестного белка.
Вторичная структура — локальное упорядочивание фрагмента полипептидной цепи, стабилизированное водородными связями. Ниже приведены самые распространённые типы вторичной структуры белков: α-спирали — плотные витки вокруг длинной оси молекулы, один виток составляют 3,6 аминокислотных остатка, и шаг спирали составляет 0,54 нм[15] (так что на один аминокислотный остаток приходится 0,15 нм), спираль стабилизирована водородными связями между H и O пептидных групп, отстоящих друг от друга на 4 звена. Спираль построена исключительно из одного типа стереоизомеров аминокислот (L). Хотя она может быть как левозакрученной, так и правозакрученной, в белках преобладает правозакрученная. Спираль нарушают электростатические взаимодействия глутаминовой кислоты, лизина, аргинина. Расположенные близко друг к другу остатки аспарагина, серина, треонина и лейцина могут стерически мешать образованию спирали, остатки пролина вызывает изгиб цепи и также нарушает α-спирали.β-листы (складчатые слои) — несколько зигзагообразных полипептидных цепей, в которых водородные связи образуются между относительно удалёнными друг от друга (0,347 нм на аминокислотный остаток[15]) в первичной структуре аминокислотами или разными цепями белка, а не близко расположенными, как имеет место в α-спирали. Эти цепи обычно направлены N-концами в противоположные стороны (антипараллельная ориентация). Для образования β-листов важны небольшие размеры боковых групп аминокислот, преобладают обычно глицин и аланин.π-спирали;310-спирали;неупорядоченные фрагменты.
Третичная или трёхмерная структура — пространственное строение полипептидной цепи (набор пространственных координат составляющих белок атомов). Структурно состоит из элементов вторичной структуры, стабилизированных различными типами взаимодействий, в которых гидрофобные взаимодействия играют важнейшую роль. В стабилизации третичной структуры принимают участие: ковалентные связи (между двумя остатками цистеина — дисульфидные мостики);ионные связи между противоположно заряженными боковыми группами аминокислотных остатков;водородные связи;гидрофильно-гидрофобные взаимодействия. При взаимодействии с окружающими молекулами воды белковая молекула «стремится» свернуться так, чтобы неполярные боковые группы аминокислот оказались изолированы от водного раствора; на поверхности молекулы оказываются полярные гидрофильные боковые группы.
Четверичная структура (или субъединичная, доменная) — взаимное расположение нескольких полипептидных цепей в составе единого белкового комплекса. Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру. В состав белка с четвертичной структурой могут входить как идентичные, так и различающиеся полипептидные цепочки. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и в стабилизации третичной. Надмолекулярные белковые комплексы могут состоять из десятков молекул.
8.Хемиосмотическая теория Митчелла. b-окисление жирных кислот.
в соответствии с хемиосмотической теорией П. Митчелла , энергия, освобождаемая в результате работы; электронтранспортной цепи, первоначально накапливается в форме трансмембранного градиента ионов водорода. Разрядка образующегося DmH+ происходит с участием локализованного в той же мембране протонного АТФ-синтазного комплекса: Н+ возвращаются по градиенту DmH+ через Н+–АТФ-синтазу, при этом без возникновения каких-либо промежуточных высокоэнергетических соединений из АДФ и неорганического фосфата образуется АТФ. (Сами сопрягающие мембраны в интактном состоянии непроницаемы для ионов, особенно Н+ и ОН–.) Предположительно, для синтеза одной молекулы АТФ достаточен перенос двух протонов, т. е. Н+/АТФ=2. Однако не исключено, что Н+/АТФ может быть больше.Локализованная в мембране H+–АТФ-синтаза катализирует реакции синтеза и гидролиза АТФ в соответствии с уравнением:
АДФ + ФН + nHНАР+ « АТФ + H2O + nHВНУТР+
Реакция, протекающая слева направо, сопряжена с транспортом H+ по градиенту DmH+, что приводит к его разрядке и синтезу АТФ. Протекающая в противоположном направлении реакция гидролиза АТФ, сопровождающаяся переносом Н+ против градиента, приводит к образованию (или возрастанию) DmH+ на мембране. Таким образом, АТФ-синтазный ферментный комплекс служит механизмом, обеспечивающим взаимное превращение двух форм клеточной энергии (DmH+ « АТФ), устройством, сопрягающим процессы окислительной природы с фосфорилированием.
b-окисление жирных кислот.Процесс b-окисления является циклическим. За каждый оборот цикла от жирной кислоты отщепляется 2 углеродных атома в виде ацетильного остатка.После этого укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает в новый цикл реакций b-окисления). Образующийся Ацетил-КоА может дальше вступить в цикл трикарбоновых кислот.Нужно уметь рассчитывать энергетический выход при распаде жирных кислот. Представленная формула верна для любой насыщенной жирной кислоты, содержащей n углеродных атомов.При распаде ненасыщенных жирных кислот образуется меньше АТФ. Каждая двойная связь в жирной кислоте - это потеря 2-х молекул АТФ.b-окисление наиболее интенсивно протекает в мышечной ткани, почках, печени.В результате b-окисления ЖК образуется Ацетил-КоА. Скорость окисления определяется скоростью процессов липолиза. Ускорение липолиза характерно для состояния углеводного голодания и интенсивной мышечной работы. Ускорение b-окисления наблюдается во многих тканях, в том числе и в печени. В печени образуется больше Ацетил-КоА, чем ей требуется. Печень - "орган-альтруист" и поэтому печень отправляет глюкозу в другие ткани.Печень стремится направить в другие ткани и свой собственный Ацетил-КоА, но не может, так как для Ацетил-КоА клеточные мембраны непроницаемы. Поэтому в печени из Ацетил-КоА синтезируются специальные вещества, которые называются "кетоновые тела". Кетоновые тела - это особая транспортная форма ацетил-КоА.
9.Биосинтез жирных кислот.
В настоящее время в достаточной степени изучен механизм биосинтеза жирных кислот в организме животных и человека, а также катализирующие этот процесс ферментные системы. Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме печеночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома.Иными словами, митохондриальная система биосинтеза жирных кислот, включающая несколько модифицированную последовательность реакций β-окисления, осуществляет только удлинение существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой кислоты из ацетил-КоА активно протекает в цитозоле, т.е. вне митохондрий, по совершенно другому пути.Внемитохондриальная система биосинтеза de novo жирных кислот (ли-погенез). Эта система находится в растворимой (цитозольной) фракции клеток многих органов, в частности печени, почек, мозга, легких, молочной железы, а также в жировой ткани. Биосинтез жирных кислот протекает с участием НАДФН, АТФ, Мn2+ и НСО3– (в качестве источника СО2); субстратом является ацетил-КоА, конечным продуктом – пальмитиновая кислота. Потребности в кофакторах процессов биосинтеза и β-окисления жирных кислот значительно различаются.Как отмечалось, строительным блоком для синтеза жирных кислот в цитозоле клетки служит ацетил-КоА, который в основном поступает из митохондрий. Было выявлено, что цитрат стимулирует синтез жирных кислот в цитозоле клетки. Известно также, что образующийся в митохондриях в процессе окислительного декарбоксилирования пирувата и окисления жирных кислот ацетил-КоА не может диффундировать в цито-золь клетки, так как митохондриальная мембрана непроницаема для данного субстрата. Поэтому вначале внутримитохондриальный ацетил-КоА взаимодействует с оксалоацетатом, в результате чего образуется цитрат. Реакция катализируется ферментом цитрат-синтазой. Образовавшийся цитрат переносится через мембрану митохондрий в цитозоль при помощи специальной трикарбоксилаттранспортирующей системы.В цитозоле цитрат реагирует с HS-KoA и АТФ, вновь распадаясь на ацетил-КоА и оксалоацетат. Эта реакция катализируется АТФ-цитрат-лиа-зой. Уже в цитозоле оксалоацетат при участии цитозольной малатдегидро-геназы восстанавливается до малата. Последний при помощи дикарбокси-латтранспортирующей системы возвращается в митохондриальный мат-рикс, где окисляется до оксалоацетата, завершая тем самым так называемый челночный цикл. уществует еще один путь переноса внутримитохондриального аце-тил-КоА в цитозоль клетки – с участием карнитина. Как отмечалось, кар-нитин играет роль переносчика ацильных групп из цитозоля в митохондрии при окислении жирных кислот. По-видимому, он может выполнять эту роль и в обратном процессе, т.е. в переносе ацильных радикалов, в том числе ацетильного радикала, из митохондрий в цитозоль клетки. Однако, когда речь идет о синтезе жирных кислот, данный путь переноса ацетил-КоА не является главным.Образование малонил-КоА. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-кар-боксилаза. Фермент содержит в качестве простетической группы биотин. Авидин – ингибитор биотина угнетает эту реакцию, как и синтез жирных кислот в целом.Установлено, что ацетил-КоА-карбоксилаза состоит из переменного числа одинаковых субъединиц, каждая из которых содержит биотин, биотинкарбоксилазу, карбоксибиотинпереносящий белок, транскарбоксилазу, а также регуляторный ал-лостерический центр, т.е. представляет собой полиферментный комплекс.Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА. Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты.Энзиматические системы, осуществляющие синтез жирных кислот, называются жирно-кислотными синтетазами. Они широко встречаются в природе и могут быть изолированы из различных одноклеточных организмов, растений и животных тканей.Жирно-кислотные синтетазы делятся на 2 группы. К первой группе относятся полиэнзимные, не поддающиеся фракционированию комплексы с мол. м. порядка 500000, в которых все индивидуальные энзимы собраны в компактную структуру. В частности, в эту группу входят жирно-кислотные синтетазы животных тканей и дрожжей.Вторая группа включает жирно-кислотные синтетазы, из которых отдельные энзимы могут быть выделены методами белкового фракционирования. Такие синтетазы встречаются у ряда микроорганизмов (в частности, у E.coli) и растений. Иными словами, в этих случаях все индивидуальные ферменты синтетазной системы находятся в виде автономных полипептидов.Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы (цистеина и фосфопантетеинового остатка, присоединенного к ОН-группе серина) и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах. Мол. масса АПБ составляет около 10000. Данный белок в синтетазной системе выполняет роль КоА. Заметим, что в животных тканях не удалось обнаружить свободного АПБ, подобного микробному. Из печени выделен полиэнзимный комплекс, содержащий все энзимы, необходимые для синтеза жирных кислот. Энзимы комплекса настолько прочно связаны друг с другом, что все попытки изолировать их в индивидуальном виде не увенчались успехом. Приводим последовательность реакций, происходящих при синтезе жирных кислот. Далее цикл реакций повторяется. Допустим, что идет синтез пальмитиновой кислоты (С16). В этом случае образованием бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы малонил-АПБ к карбоксильному концу растущей цепи жирной кислоты. При этом отщепляется дистальная карбоксильная группа малонил-АПБ в виде СО2. Завершается синтез жирной кислоты отщеплением HS-АПБ от ацил-АПБ под влиянием фермента деацилазы. Или, учитывая, что на образование одной молекулы малонил-КоА из ацетил-КоА расходуются одна молекула АТФ и одна молекула СО2, которая затем отщепляется. По сравнению с β-окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитозоле клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путем связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ); при биосинтезе образуется D(–)-изомер 3-гидроксикис-лоты, а не L(+)-изомер, как это имеет место при β-окислении жирных кислот; необходимость для синтеза жирных кислот кофермента НАДФН. Последний в организме частично (на 50%) образуется в реакциях пен-тозофосфатного цикла, частично – в других реакциях, в частности в реакциях:
Малат + НАДФ+-> Пируват + С02 + НАДФН + Н+ Изоцитрат + НАДФ+-> α-Кетоглутарат + С02 + НАДФН + Н +.Образование ненасыщенных жирных кислот. Элонгация жирных кислот. В отличие от растительных тканей ткани животных обладают весьма ограниченной способностью превращать насыщенные жирные кислоты в ненасыщенные.Установлено, что две наиболее распространенные мононенасыщенные жирные кислоты – пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и стеариновой кислот.Эти превращения протекают в микросомах клеток печени и жировой ткани при участии молекулярного кислорода, восстановленной системы пиридиновых нуклеотидов и цитохрома b5. Превращению подвергаются только активированные формы пальмитиновой и стеариновой кислот. Ферменты, участвующие в этих превращениях, получили название деса-тураз.Наряду с десатурацией жирных кислот (образование двойных связей) в микросомах происходит и их удлинение (элонгация), причем оба эти процесса могут сочетаться и повторяться. Удлинение цепи жирной кислоты происходит путем последовательного присоединения к соответствующему ацил-КоА двууглеродных фрагментов при участии малонил-КоА и НАДФН. Энзиматическая система, катализирующая удлинение жирных кислот, получила название элонгазы. На схеме представлены пути превращения пальмитиновой кислоты в реакциях десатурации и элонгации.
10.Световая стадия фотосинтеза: ЭТЦ хлоропластов.
Организация электрон-транспортной цепи хлоропластов . Общая характеристика "Z"-схемы фотосинтеза. Участие двух фотохимических реакций в фотосинтезе растений. Эффекты Эмерсона, Блинкса. Химическая природа основных компонентов электрон-транспортной цепи ( ЭТЦ ), последовательность их взаимодействия. Локализация ЭТЦ в мембране.Структурно-функциональная организация ФС I и ФС II. Основные пептиды комплексов, их расположение в мембране, локализация пигментов и редокс агентов. Гетерогенность фотосистем.Комплекс ФС I. Механизм образования в хлоропластах веществ с высоким восстановительным потенциалом; основные пути использования восстановительного потенциала в конструктивных и фоторегуляторных процессах. Энзиматические системы, участвующие в генерации восстановительного потенциала, их гетерогенность. Ферредоксины; основные физико-химические свойства и роль в восстановительных реакциях фотосинтеза. ФД-НАДФ-редуктаза; основные формы и функции фермента.Комплекс ФС II. Участие ФС II в процессе окисления воды. Современные представления о механизме фотоокисления воды; кинетика и химизм реакций. Структура Mn-содержащего комплекса. Фотоингибирование ФС II. Возможные пути фотодеструкции комплекса ФС II; включение защитных механизмов. Ингибиторный и флуоресцентный анализы как методы исследования акцепторной зоны ФС II; механизм действия гербицидов.Хиноны хлоропластов . Основные группы хинонов, их химическая структура, физиологическая роль. Пул пластохинонов, основные функции.Цитохромы хлоропластов . Цитохром b6/f-комплекс, его организация и функционирование в ЭТЦ . Окисление пластохинолов в реакциях Q-цикла; участие в сопряжении редокс-энергии с образованием трансмембранного электрохимического градиента протонов. Кинетика работы Q-цикла; механизмы, контролирующие скорость работы цикла. Участие цитохромного комплекса в реакциях циклического транспорта электронов. Система цитохрома b559. Основные редокс-формы, их взаимообратимость, функциональное значение.Пластоцианин. Молекулярная структура, механизм взаимодействия с цитохромным комплексом и ФС I.Основные принципы моделирования ЭТЦ . Ингибиторы транспорта электронов, локализация их центров действия. Искусственные электрон-акцепторные и электрон-донорные системы, используемые при моделировании различных участков ЭТЦ .Кинетические закономерности работы цепи. Соотношение двух фотосистем, механизмы их координированного взаимодействия. Долговременные и кратковременные механизмы адаптации тилакоидных систем к условиям освещенности. Циклические и нециклические потоки электронов. Основные механизмы и скорость переноса электронов на отдельных участках цепи. Система регуляции электронных потоков; .основные регуляторные центры; влияние конформационно-активных факторов и редокс-агентов. Взаимодействие компонентов ЭТЦ с кислородом. Фотосинтетическое фосфорилирование. Характеристика основных типов фотофосфорилирования. Циклическое и нециклическое фотофосфорилирование; физиологическая роль, пути транспорта электронов, механизмы регуляции. Механизм фотофосфорилирования. Основные положения хемиосмотической теории Митчела. Механизм и кинетика формирования градиента электрохимического потенциала ионов водорода; электрогенные и протолитические реакции хлоропластов . Сопрягающий комплекс хлоропластов. Структурно-функциональная организация и взаимодействие субъединиц CFо и CF1. Конформационная гипотеза сопряжения Бойера и ее развитие в работах последних лет. Ротационный механизм работы АТФ-синтазы. Работа каталитического центра CF1. Изменения энергии связывания нуклеотидов в каталитических центрах CF1 как основа образования АТФ в процессе фотофосфорилирования. АТФазная функция CFo-CF1 комплекса, ее физиологическая роль. Активация CFo-CF1 комплекса.
12.Фотосинтез: цикл Кальвина и Хэтча-Слейка.
В цикл вовлекаются АТФ и НАДФ·Н, образованные в ЭТЦ фотосинтеза, углекислый газ и вода; основным продуктом являеся глицеральдегид-3-фосфат. Поскольку АТФ и НАДФ·Н могут образовываться в разных метаболических путях, цикл не следует рассматривать строго привязанным к световой фазе фотосинтеза.Общий баланс реакций цикла можно представить уравнением:
3 CO2 + 6 НАДФ·Н + 5 H2O + 9 АТФ → C3H5O3-PO3 + 3 H+ + 6 НАДФ+ + 9 АДФ + 8 Фн + 3 H2O
Две молекулы глицеральдегид-3-фосфата используются для синтеза глюкозы.
Цикл состоит из трёх стадий: на первой под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа происходит присоединение CO2 к рибулозо-1,5-дифосфату и расщепление полученной гексозы на две молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй 3-ФГК восстанавливается до глицеральдегид-3-фосфата (фосфоглицеральдегида, ФГА), часть молекул которого выходит из цикла для синтеза глюкозы, а другая часть используется в третьей стадии для регенерации рибулозо-1,5-дифосфата.
КарбоксилированиеКарбоксилирование рибулозо-1,5-бисфосфата (5-углеродное соединение) осуществляется рубиско в несколько стадий. На первой кетонная группа рибулозы восстанавливается до спиртовой, между 2 и 3 атомами углерода устанавливается двойная связь. Полученное соединение нестабильно и именно оно карбоксилируется с образованием 2-карбокси-3-кето-D-арабитол-1,5-бисфосфата. Его структурный аналог 2-карбокси-D-арабитол-1,5-бисфосфат ингибирует весь процесс. Новое, уже 6-углеродное соединение, также нестабильно и распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерат, 3-ФГА).
ВосстановлениеВосстановление 3-фосфоглицериновой кислоты (3-ФГА) происходит в две реакции.Сначала каждая 3-ФГА с помощью 3-фосфоглицераткиназы и с затратой одной АТФ фосфорилируется, образуя 1,3-бисфосфоглицериновая кислота (глицерат-1,3-бисфосфат).
Затем под действием глицеральдегид-1,3-фосфатдегидрогеназы бисфосфоглицериновая кислота восстанавливается НАД(Ф)·H (у растений и цианобактерий; у пурпурных и зелёных бактерий восстановителем является НАД·H) параллельно с отщеплением одного остатка фосфорной кислоты. Образуется глицеральдегид-3-фосфат (фосфоглицеральдегид, ФГА, триозофосфат). Обе реакции обратимы.
РегенерацияНа последней стадии 5 молекул глицеральдегид-3-фосфатов превращаются в три молекулы рибулозо-1,5-бисфосфата.Вначале под действием трифосфат-изомеразы глицеральдегид-3-фосфат изомеризуется в дигидроксиацетон-фосфат. Фруктозабисфосфат-альдолаза объединяет их в фруктозо-6-фосфат с отщеплением остатка фосфорной кислоты. Затем следует ряд реакций перестройки углеродных скелетов и образуется рибулозо-5-фосфат. Он фосфорилируется фосфорибулокиназой и рибулозо-1,5-бисфосфат регенерируется.
13.Понятие о фитохромной системе растений.
Фитохромная система участвует в измерении растением длины дня и ночи. В условиях длинного дня длиннодневные растения цветут, а коротко дневные нет. В условиях короткого дня, наоборот, короткодневные растения цветут, а длинно дневные нет. Однако минутное прерывание темнового периода красным светом подавляет цветение короткодневных растений и вызывает цветение длиннодневных. Это действие красного света снимается, если вслед за красным светом следует освещение дальним красным светом. Таким образом, обратимые изменение фитохрома участвуют в регуляции цветения у растений. Это было показано исследованиями Х. Бортвика и М. Паркера. Крупный вклад внес М.Х. Чайлахян. У длиннодневных растений цветение вызывает гормон гибберллинов.
Фитохром представляет собой белок (апопротеин), к которому присоединен поглащающий свет пигмент. Молекулярная масса белка фитохрома 250 кД. Он состоит из двух одинаковых субъединиц. К каждой из субъединиц ковалентно присоединена через тиоэфирную связь одна молекула поглощающего свет пигмента – хромофора, который представляет собой тетрапиррол и называется фитохромобилином.
В результате поглощения красного света хромофор в составе фитохрома ФКк претерпевает цис-транс-изомерезацию за счет вращение молекулы относительно двойной связи между 15-м и 16-м улерода тетрапиррола. В результате хромофор ФХк формы фитохрома превращается в хромофор ФХДК формы. Изменение В хромофоре передаются белку и приводят к изменению его конформации, которые далее возбуждают в клетках цепь сигналов, приводящих к фотоморфогенезу или иным изменением в жизни растений. Под действием ДК света молекулы ФХДК формы превращаются в ФХк.
14.Ферменты, регулируемые путём ковалентной модификации.
Ферменты, регулируемые путем ковалентной модификации. Фазы метаболизма-катаболизм и анаболизм. Некоторые ферменты изменяют свою каталитическую активность в результате белок-белковых взаимодействий. Рассмотрим 2 механизма активации ферментов с помощью белок-белковых взаимодействий: активация ферментов в результате присоединения регуляторных белков;изменение каталитической активности ферментов вследствие ассоциации или диссоциации протомеров фермента.Активация ферментов в результате присоединения регуляторных белков. Этот тип регуляции можно рассмотреть на примере активации фермента аденилатциклазы, локализованной в плазматической мембране клетки. Активный центр аденилатциклазы локализован на цитоплазматической стороне плазматической мембраны. Активированная аденилатциклаза катализирует реакцию образования из АТФ циклического 3',5'-АМФ (цАМФ) - вторичного, внутриклеточного посредника действия гормонов (см. схему ниже). В мембране аденилатциклаза функционирует в комплексе с другими белками: рецептором гормона, выступающего во внеклеточную среду и взаимодействующего с гормонами; G-белком, занимающим промежуточное положение между рецептором и ферментом аденилатциклазой. G-белок - олиго-мерный белок, состоящий из 3 субъединиц - α, β, γ. α-Субъединица имеет центр связывания и расщепления ГТФ. Поэтому этот белок называется ГТФ-связывающим белком, или G-белком;в результате связывания гормона с рецептором происходит изменение конформа-ции G-белка, уменьшение его сродства к молекуле ГДФ, с которой он связан в отсутствие гормонального сигнала, и увеличение сродства к ГТФ. Присоединение ГТФ вызывает конформационные изменения в G-белке и диссоциацию его на субъединицы: субъединицу α, связанную с ГТФ (α-ГТФ), димер βγ;α-ГТФ имеет высокое сродство к аденилатциклазе, его присоединение приводит к активации последней, поэтому α-ГТФ - регуляторный белок, а данный механизм активации аденилатциклазы называют активацией ферментов в результате присоединения регуляторных белков (рис. 2-32). Регуляция каталитической активности ферментов ассоциацией/диссоциацией протомеров Протеинкиназы - группа ферментов, катализирующих перенос остатка фосфорной кислоты с АТФ на специфические ОН-группы аминокислотных остатков белков (вызывают фосфорилирование белков). Механизмы активации различных протеинкиназ неодинаковы. В качестве примера регуляции каталитической активности ферментов ассоциацией или диссоциацией протомеров можно привести регуляцию активности фермента Протеинкиназы А. Протеинкиназа А (цАМФ-зависимая) состоит из 4 субъединиц 2 типов: 2 регуляторных (R) и 2 каталитических (С). Такой тетрамер не обладает каталитической активностью. Регуляторные субъединицы имеют участки связывания для циклического 3',5'-АМФ (цАМФ), по 2 на каждую субъединицу. Присоединение 4 молекул цАМФ к 2 регуляторным субъединицам приводит к изменению конфор-мации регуляторных протомеров и к диссоциации тетрамерного комплекса, при этом высвобождаются 2 активные каталитические субъединицы (рис. 2-32). Такой механизм регуляции обратим. Отщепление молекул цАМФ от регуляторных субъединиц приведёт к ассоциации регуляторных и каталитических субъединиц Протеинкиназы А с образованием неактивного комплекса. Рис. 2-32. Регуляция активности аденилатциклазы. Гормон (Г), взаимодействуя с рецептором (R) на поверхности клеток, приводит к уменьшению сродства ГТФ-связывающего белка (G-белка, состоящего из протомеров α, β, γ) к ГТФ и увеличению сродства к ГТФ. Присоединение молекулы ГТФ к активному центру G-белка вызывает диссоциацию комплекса на субъединицы α-ГТФ и димер βγ. Комплекс α-ГТФ активирует аденилатциклазу, что способствует синтезу из АТФ внутриклеточных регуляторных молекул цАМФ. АЦ - аденилатциклаза, ПКА - протеинкиназа А, Рi - Н3РО4. Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов - фосфорилирование/дефосфорилирование. Модификации подвергаются ОН-группы фермента. Фос-форилирование осуществляется ферментами протеинкиназами, а дефосфорилирование фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической активности. При этом результат может быть двояким: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными (рис. 2-33). Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфо-рилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки. Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала - жира, гликогена и усиление его распада (мобилизация), вызванного фосфо-рилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.
Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
Рассмотрим механизм частичного протеолиза на примере активации протеолитического фермента трипсина (рис. 2-34). Трип-синоген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Следует напомнить, Рис. 2-33. Регуляция активности ферментов фосфорилированием/дефосфорилированием.что трипсин относят к семейству "сериновых" протеаз - активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз - пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.
15.Аллостерические ферменты.
-ферменты, активность которых регулируется не только количеством молекул субстрата, но и другими веществами, называемыми эффекторами. Участвующие в аллостерической регуляции эффекторы - клеточные метаболиты часто именно того пути, регуляцию которого они осуществляют.
АФ играют важную роль в метаболизме, т.к. чрезвычайно быстро реагируют на малейшие изменения внутреннего состояния клетки. Аллостерическая регуляция имеет большое значение в следующих ситуациях: при анаболических процессах. Ингибирование конечным продуктом метаболического пути и активация начальными метаболитами позволяют осуществлять регуляцию синтеза этих соединений;при катаболических процессах. В случае накопления АТФ в клетке происходит ингибирование метаболических путей, обеспечивающих синтез энергии. Субстраты при этом расходуются на реакции запасания резервных питательных веществ;для координации анаболических и катаболических путей. АТФ и АДФ - аллостерические эффекторы, действующие как антагонисты;для координации параллельно протекающих и взаимосвязанных метаболических путей (например, синтез пуриновых и пиримидиновых нуклеотидов, используемых для синтеза нуклеиновых кислот). Таким образом, конечные продукты одного метаболического пути могут быть аллостерическими эффекторами другого метаболического пути. Аллостерические эффекторы. Эффектор, вызывающий снижение (ингибирование) активности фермента, называют отрицательным эффектором, или ингибитором. Эффектор, вызываюший повышение (активацию) активности ферментов - положительный эффектор, или активатором. Аллостерическими эффекторами часто служат различные метаболиты. Конечные продукты метаболического пути - часто ингибиторы АФ, а исходные вещества - активаторы. Это так называемая гетеротропная регуляция. Такой вид аллостерической регуляции очень распространён в биологических системах. Более редкий случай аллостерической регуляции, когда сам субстрат может выступать в качестве положительного эффектора. Такая регуляция называется гомотропной (эффектор и субстрат - одно и то же вещество). Эти ферменты имеют несколько центров связывания для субстрата, которые могут выполнять двойную функцию: каталитическую и регуляторную. АФ такого типа используются в ситуации, когда субстрат накапливается в избытке и должен быстро преобразоваться в продукт. Выявить ферменты с аллостерической регуляцией можно, изучая кинетику этих ферментов. Эти ферменты не подчиняются законам Михаэлиса-Ментен, они имеют характерную S-образную кривую зависимости скорости реакции от концентрации субстрата. Особенности строения и функционирования АФ: обычно это олигомерные белки, состоящие из нескольких протомеров или имеющие доменное строение;они имеют аллостерический центр, пространственно удалённый от каталитического активного центра;эффекторы присоединяются к ферменту нековалентно в аллостерических (регуляторных) центрах;аллостерические центры, так же, как и каталитические, могут проявлять различную специфичность по отношению к лигандам: она может быть абсолютной и групповой. Некоторые ферменты имеют несколько аллостерических центров, одни из которых специфичны к активаторам, другие - к ингибиторам. протомер, на котором находится аллостерический центр, - регуляторный протомер, в отличие от каталитического протомера, содержащего активный центр, в котором проходит химическая реакция;аллостерические ферменты обладают свойством кооперативности: взаимодействие аллостерического эффектора с аллостерическим центром вызывает последовательное кооперативное изменение конформации всех субъединиц, приводящее к изменению конформации активного центра и изменению сродства фермента к субстрату, что снижает или увеличивает каталитическую активность фермента; регуляция аллостерических ферментов обратима: отсоединение эффектора от регуляторной субъединицы восстанавливает исходную каталитическую активность фермента;аллостерические ферменты катализируют ключевые реакции данного метаболического пути.
16.Ферменты, регулируемые путём ковалентной модификации.
Ферменты, регулируемые путем ковалентной модификации. Фазы метаболизма-катаболизм и анаболизм. Некоторые ферменты изменяют свою каталитическую активность в результате белок-белковых взаимодействий. Рассмотрим 2 механизма активации ферментов с помощью белок-белковых взаимодействий: активация ферментов в результате присоединения регуляторных белков;изменение каталитической активности ферментов вследствие ассоциации или диссоциации протомеров фермента.Активация ферментов в результате присоединения регуляторных белков. Этот тип регуляции можно рассмотреть на примере активации фермента аденилатциклазы, локализованной в плазматической мембране клетки. Активный центр аденилатциклазы локализован на цитоплазматической стороне плазматической мембраны. Активированная аденилатциклаза катализирует реакцию образования из АТФ циклического 3',5'-АМФ (цАМФ) - вторичного, внутриклеточного посредника действия гормонов (см. схему ниже). В мембране аденилатциклаза функционирует в комплексе с другими белками: рецептором гормона, выступающего во внеклеточную среду и взаимодействующего с гормонами; G-белком, занимающим промежуточное положение между рецептором и ферментом аденилатциклазой. G-белок - олиго-мерный белок, состоящий из 3 субъединиц - α, β, γ. α-Субъединица имеет центр связывания и расщепления ГТФ. Поэтому этот белок называется ГТФ-связывающим белком, или G-белком;в результате связывания гормона с рецептором происходит изменение конформа-ции G-белка, уменьшение его сродства к молекуле ГДФ, с которой он связан в отсутствие гормонального сигнала, и увеличение сродства к ГТФ. Присоединение ГТФ вызывает конформационные изменения в G-белке и диссоциацию его на субъединицы: субъединицу α, связанную с ГТФ (α-ГТФ), димер βγ;α-ГТФ имеет высокое сродство к аденилатциклазе, его присоединение приводит к активации последней, поэтому α-ГТФ - регуляторный белок, а данный механизм активации аденилатциклазы называют активацией ферментов в результате присоединения регуляторных белков (рис. 2-32). Регуляция каталитической активности ферментов ассоциацией/диссоциацией протомеров Протеинкиназы - группа ферментов, катализирующих перенос остатка фосфорной кислоты с АТФ на специфические ОН-группы аминокислотных остатков белков (вызывают фосфорилирование белков). Механизмы активации различных протеинкиназ неодинаковы. В качестве примера регуляции каталитической активности ферментов ассоциацией или диссоциацией протомеров можно привести регуляцию активности фермента Протеинкиназы А. Протеинкиназа А (цАМФ-зависимая) состоит из 4 субъединиц 2 типов: 2 регуляторных (R) и 2 каталитических (С). Такой тетрамер не обладает каталитической активностью. Регуляторные субъединицы имеют участки связывания для циклического 3',5'-АМФ (цАМФ), по 2 на каждую субъединицу. Присоединение 4 молекул цАМФ к 2 регуляторным субъединицам приводит к изменению конфор-мации регуляторных протомеров и к диссоциации тетрамерного комплекса, при этом высвобождаются 2 активные каталитические субъединицы (рис. 2-32). Такой механизм регуляции обратим. Отщепление молекул цАМФ от регуляторных субъединиц приведёт к ассоциации регуляторных и каталитических субъединиц Протеинкиназы А с образованием неактивного комплекса. Рис. 2-32. Регуляция активности аденилатциклазы. Гормон (Г), взаимодействуя с рецептором (R) на поверхности клеток, приводит к уменьшению сродства ГТФ-связывающего белка (G-белка, состоящего из протомеров α, β, γ) к ГТФ и увеличению сродства к ГТФ. Присоединение молекулы ГТФ к активному центру G-белка вызывает диссоциацию комплекса на субъединицы α-ГТФ и димер βγ. Комплекс α-ГТФ активирует аденилатциклазу, что способствует синтезу из АТФ внутриклеточных регуляторных молекул цАМФ. АЦ - аденилатциклаза, ПКА - протеинкиназа А, Рi - Н3РО4. Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов - фосфорилирование/дефосфорилирование. Модификации подвергаются ОН-группы фермента. Фос-форилирование осуществляется ферментами протеинкиназами, а дефосфорилирование фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической активности. При этом результат может быть двояким: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными (рис. 2-33). Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфо-рилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки. Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала - жира, гликогена и усиление его распада (мобилизация), вызванного фосфо-рилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.
Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
Рассмотрим механизм частичного протеолиза на примере активации протеолитического фермента трипсина (рис. 2-34). Трип-синоген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Следует напомнить, Рис. 2-33. Регуляция активности ферментов фосфорилированием/дефосфорилированием.что трипсин относят к семейству "сериновых" протеаз - активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз - пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.
17.Центральный путь катаболизма глюкозы.
Катаболизм – фаза метаболизма, на которой происходит распад молекул, поступающих с пищей или запасенных живым организмом до неорганических веществ (H2O, CO2, NH4). В ходе катаболических процессов высвободившаяся при окислении энергия запасается в форме АТР, NAD(P)H и мембранного потенциала. на первой стадии происходит активация субстратов за счет фосфорилирования (затрачивается 2 АТР), включение в данный процесс других сахаров, распад на две С3 молекулы (глицеральдегид-3-фосфат и дигидроксиацетонфосфат).На втором этапе происходит окисление глицеральдегид-3-фосфата до пирувата (в аэробных условиях) или до лактата (при анаэробиозе) с образованием 4-х молекул АТР.
Глюкоза гексокиназа Глюкозо-6-фосфат фосфоглюкоизомераза Фруктозо-6-фосфат фосфофруктокиназа Фруктозо-1,6-бисфосфат альдолаза Глицеральдегид-3- фосфат Дигидроксиацетонфосфат триозофосфатизомераза
Запасание энергии
Глицеральдегид-3- фосфат
глицеральдегид-3-фосфатдегидрогеназа
(NAD+ NADH)
1,3-дифосфоглицерат
фосфоглицераткиназа (АДР АТР)
3-фосфоглицерат
фосфоглицератмутаза
2-фосфоглицерат енолаза фосфоенолпируват пируваткиназа (АДР АТР) пируват
18.Регуляторные ферменты, их функция. Примеры.
Регуляторные (аллостерические) ферменты воспринимают различные метаболические сигналы и в соответствии с ними изменяют свою каталитическую активность.
Одним из уникальных свойств живых организмов является удивительная их способность к сохранению сбалансированности катаболических (биодегра-дативных) и анаболических (биосинтетических) процессов. При этом в клетках одновременно совершаются процессы синтеза, распада и взаимопревращения сотен и тысяч разнообразных веществ, которые в свою очередь регулируются множеством механизмов, обеспечивающих постоянство внутренней среды организма. Некоторые из этих регуляторных механизмов, среди которых важная роль принадлежит механизмам регуляции синтеза и каталитической активности ферментов, будут рассмотрены далее. Активность ферментов в клетке зависит от количества молекул субстрата, продукта, наличия кофакторов и коферментов. Действие ферментов в клетке, как правило, строго упорядочено: продукт одной ферментативной реакции является субстратом другой, образуя таким образом "метаболические пути". Среди множества ферментов практически каждого метаболического пути различают ютючевые, или регуляторные, ферменты, активность которых может изменяться в зависимости от потребности клетки в конечном продукте метаболического пути. Регуляторные ферменты расположены, как правило, в начале и/или в месте разветвления метаболического пути. Они катализируют либо самые медленные (скорость-лимитирующие реакции), либо необратимые реакции.
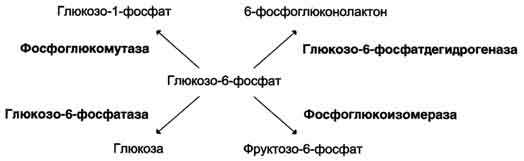
19.Энергетическая и биохимическая стадии фотосинтеза (продукты реакций).
под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества. Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.Фотохимическая суть процессаХлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции H2O → O2 E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.Светособирающие комплексыХлорофилл выполняет две функции: поглощения и передачу энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10-10-10-12 сек., расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.Основные компоненты электронтранспортной цепиФотосистема IIФотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.b6f или b/f-комплексb6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.Фотосистема IСветособирающий комплекс I содержит примерно 200 молекул хлорофилла.В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.Циклический и псевдоциклический транспорт электронаПомимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадияВ темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6H12O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
Карбоксилирования,Восстановления,Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа (Rubisco). Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы, затем НАДФН при воздействии триозофосфатдегидрогеназы её карбоксильная группа окисляется до альдегидной и она становится углеводом (ФГА).В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтезПри низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.С4 фотосинтез практические не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтезПри CAM (англ. Crassulaceae acid metabolism - кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
20.Фазы метаболизма – катаболизм и анаболизм.
Катаболизм – фаза метаболизма, на которой происходит распад молекул, поступающих с пищей или запасенных живым организмом до неорганических веществ (H2O, CO2, NH4). В ходе катаболических процессов высвободившаяся при окислении энергия запасается в форме АТР, NAD(P)H и мембранного потенциала.
Анаболизм – Синтез биологических макромолекул из низкомолекулярных предшественников. Для протекания анаболизма требуется постоянный приток энергии.
21.Трансформация световой энергии при фотосинтезе. Регуляция процесса.
Сущность фотофизического этапа фотосинтеза заключается в трансформации световой энергии в энергию разделенных зарядов.ХЛ+hV=ХЛ*+е-.ФСЕ – белковый комплекс включающий светособирающие пигменты (антенный комплекс), передающий энергию на реакционный центр (хлорофилл ловушка –хл.а, акцепторы е и доноры).ФСЕ фотосинтетическая единица состоит из светособирающих пигментов реакционного центра. Структура Р.Ц.У высших растений два типа реакционных центра. 1) В качестве хлорофилл ловушки выступает хл.а700. Он сопряжен с акцептором (вкл. белковый комплекс хл.а695 ) и донором – пластоцианином (содержит медь)2) В реакционном центре второго типа в качестве хлорофилл ловушки выступает хл680 , в качестве акцептора выступает фиофитин, а в качестве донора белок.Принцип работы реакционного центра заключается в осуществлении восстановления акцептора. Здесь протекают реакции только физического типа, для которых характерно:высокий скорость протекания (10-5-10-12 );
Независимость скорости протекания от температуры.
Сущность фотохимического этапа – трансформация энергии разделенных зарядов в макроэргическую энергию ATP, NADFH ХЛ+ + е + АДР + Pi + NADF + H2O=ХЛ + ATP + NADFH + 1/2О2.Фотосистема сложный белковый комплекс, включающий фотосинтетическую единицу и электроннотранспортную цепь.Эффект усиление Эмерсона – резкое увеличение фотосинтеза при облучении разной длины красным светом.Фотосинтез осуществляется двумя фотохимическими реакциями, которые функционируют последовательно (одна возбуждает другую).Из реакционного центра П680 под действием квантом света выделяются два е и по электроннотранспортной цепи феофитин, пластохинон, цитохром в559 (в котором происходит образование одной молекулы ATP), цитохром f и на реакционный центр П700 из которого под действием квантов света 2 е через акцептор 1, акцептор 2, фередоксин, флаупротенин и на NADF, где происходит его восстановление, два протона Н транспортируется от диализа воды при котором еще образуется 2 е которые восстанавливают Р.Ц. П680.В условиях интенсивного освещения в фотосистемах возникает дефицит NADF+. В этом случае действует циклический транспорт е.Отличие от нециклического транспорта е.ЦТЭ обеспечивается 1-й ФС I;Продуктами ЦТЭ является АТР;ЦТЭ – вспомогательный процесс включающийся для зашиты основного пути от избытка света, повышает эффективность фотосинтеза.Регуляция скорости движения е:Интенсивность освещение;состояние системы сопрягающей системы, т.е. переносчики в фотосистемах должны быть в восстановлено-окисленном состоянии;Оказывает влияние на ν кислород. Он является электрофилонным соединением, как конкурент по переноске е, в этом случае осуществляется псевдоциклический транспорт е. Его продукты АТР и перекись водорода.
22.Роль фитохромной системы в регуляции процесса цветения у растений.
Фитохромная система участвует в измерении растением длины дня и ночи. В условиях длинного дня длиннодневные растения цветут, а коротко дневные нет. В условиях короткого дня, наоборот, короткодневные растения цветут, а длинно дневные нет. Однако минутное прерывание темнового периода красным светом подавляет цветение короткодневных растений и вызывает цветение длиннодневных. Это действие красного света снимается, если вслед за красным светом следует освещение дальним красным светом. Таким образом, обратимые изменение фитохрома участвуют в регуляции цветения у растений. Это было показано исследованиями Х. Бортвика и М. Паркера. Крупный вклад внес М.Х. Чайлахян. У длиннодневных растений цветение вызывает гормон гибберллинов.
Фитохром представляет собой белок (апопротеин), к которому присоединен поглащающий свет пигмент. Молекулярная масса белка фитохрома 250 кД. Он состоит из двух одинаковых субъединиц. К каждой из субъединиц ковалентно присоединена через тиоэфирную связь одна молекула поглощающего свет пигмента – хромофора, который представляет собой тетрапиррол и называется фитохромобилином.
В результате поглощения красного света хромофор в составе фитохрома ФКк претерпевает цис-транс-изомерезацию за счет вращение молекулы относительно двойной связи между 15-м и 16-м улерода тетрапиррола. В результате хромофор ФХк формы фитохрома превращается в хромофор ФХДК формы. Изменение В хромофоре передаются белку и приводят к изменению его конформации, которые далее возбуждают в клетках цепь сигналов, приводящих к фотоморфогенезу или иным изменением в жизни растений. Под действием ДК света молекулы ФХДК формы превращаются в ФХк.
23.Образование первичных аминокислот в растениях.
Пути синтеза глутамат:
глутаминсинтетазный
глутаматдегидрогиназный
Это основной путь, он осуществляется с помощью двух реакций.
а )
глутамат + NH4++ATPглутаматсинтетаза
глутамин + АДР +Рi
)
глутамат + NH4++ATPглутаматсинтетаза
глутамин + АДР +Рi
Глутаматсинтетаза – малекулярная масса 300 кД, состоит из 8 субъединиц, локализован в хлоропластах и цитоплазме.
б) синетез глутамата – фермент глутаматсинтаза
Глутамин + 2оксоглутаровая кислота + Фдв= 2 глутамат + 2 Фдо
Глутаматсинтаза – Mr=145-180кД
Кол-во субъединиц – 2, локализован в хлоропластах
Предполагает прямое аминирование 2 –оксоглутаратовой кислоты, использую в качестве ко - фактора NADFH
2-оксоглутаровая к-та + NADFH + NH4+= глутамат + NAD
Фермент глутаматдегидрогиназа – имеет 6 субъединиц, Mr =300 кД. Локализован в митохондриях..
Главным источником аминогрупп в ракции переаминирования является глутамат +кетокислота.
глутамат + глиоксилат глицин
пируват = аланин + 2оксоглутаровая кислота
оксалоацитат аспартат
Ферменты специфические трансаминазы.
24.Биохимические пути ассимиляции углекислого газа растениями C3 и С4 – типа.
В 1957 году Кальвин разработал цикл усвоения CO2.
CO2 + АТФ + NADPH = CH2O → C6H12O6
Был использован изотопный метод. C14O2. Растение было помещено в специальную камеру, где контролировались все условия. Через 1 сек. вся рад. сосредоточилась в фосфоглицериновой кислоте. (C3 фосфол.).
Карбоксилирование первичного акцептора
РМФ + АТФ = РБФ + АДФ Р 1,5 бисфосфат – первичный акцептор CO2
РБФ + CO2 = [C6] → 2 ФГК фосфоглицериновая кислота
Фермент РУБИСКО рибулёзобисфосфаткарбоксилаза. Состоит из 2 х субъединиц А8В8
А – активный центр, ф-ция карбоксилирования
В – регулятор активности белковой молекулы
Так же фермент владеет бифункциональностью: - Карбоксилирование и в присутствии O2 обладает оксигеназной ф-цией.
Восстановление фосфо-глицириновой кислоты.
3ФГА + АТФ = 1,3 ФГК + АДФ
1,3 ФГК + NADPH = ФГА
Восстановление первичного акцептора
3РМФ + 3 CO2 + 9АТФ + 6NADPH = 6ФГА + 9АДФ + 6NADP
6ФГА: С3(1) + С3(2) = С6
С6 = С4 + С2
И т.д.
В 1964 году Карпилов обнаружил, что метка рад СО2 через 2 сек. в клетках кукурузы в яблочной кислоте (С4).
В 1967 Хетч и Слейк разработали новый цикл поглощения СО2 при фотосинтезе. Растение обладает Krans анатомией листа.
Все растения С4 типа прекрасно адаптированы к современным условиям среды.
Значение цикла:
У С4 растений функционирует ФЕПК карбоксилаза, обладающая более высоким сродством к СО2, т.к. современные растения существуют в условиях дефицита СО2
ФЕПК – термофильный фермент, способен активно работать при 35-40 градусах (РУБИСКО 25+30).
Он не способен к оксигеназной реакции
Следовательно С4 растения не имеют фотодыхания, процесса, который в присутствии кислорода разрушает органическое вещество до СО2.
Продуктивность С4 растений почти в 2 раза больше С3.
Отличая цикла Х-С от Кальвина
Осуществляется в клетках мезофилла
Производит псевдоассимиляцию СО2
Роль цикла Х-С:
Кооперативный цикл, т.е. взаимодействие с циклом Кальвина, обеспечивая его СО2
Наличие ФЕПК обеспечивает эффективную утилизацию СО2 в условиях её низкой концентрации.
Отсутствие или низкая интенсивность фотодыхания. Если РУБИСКО проявляет оксигеназную функцию, то выделяющийся СО2 поглощается ФЕПК.
25.Механизмы окислительного фосфорилирования.
Это процесс образования АТФ из АДФ и Рнеорг., сопряженный с переносом электронов по ЭТЦ МТХ. (Если для фотосинтеза, то на свету и в мембране тилакойдов). В настоящее время наибольшее признание получила теория П. Митчелла (для МТХ). Ее адаптировал Ягендорф для пластид.
П680 возбуждается светом и теряет электрон. Далее электрон идет на феофетин – переносчик ЭТЦ. Главным правилом ЭТЦ является то, что каждый следующий переносчик должен быть более положительным, чем предыдущий. Электрон передается на пластохинон. Он работает как протонный насос и выбрасывает Н+. П700 возбуждается светом и теряет электрон. Электрон передается на феродоксин. А с него на NADP. Внутренняя мембрана заряжается +, а наружная -. Возникает трансмембранный потенциал. При достижение значения 250мВ, энергия может быть трансформирована в АТФ. Есть специальный ферментный комплекс в мембране – АТФсинтаза. Она синтезирует АТФ из АДФ и Рнеорг. Но только если трансмембранный потенциал достигает значения 250мВ. При нем возникает конфармационная подвижка и образуется щель в молекуле, ч/з которую устремляется Н+ с внутренней стороны на наружную. Н+ соединяется с NADP и образуется NADPH. При проходе ч/з АТФсинтазу они обеспечивают ее энергией и к АДФ присоединяется Рнеорг. Дорисовать:
26.b-окисление жирных кислот.
Процесс b-окисления является циклическим. За каждый оборот цикла от жирной кислоты отщепляется 2 углеродных атома в виде ацетильного остатка.После этого укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает в новый цикл реакций b-окисления). Образующийся Ацетил-КоА может дальше вступить в цикл трикарбоновых кислот.Нужно уметь рассчитывать энергетический выход при распаде жирных кислот. Представленная формула верна для любой насыщенной жирной кислоты, содержащей n углеродных атомов.При распаде ненасыщенных жирных кислот образуется меньше АТФ. Каждая двойная связь в жирной кислоте - это потеря 2-х молекул АТФ.b-окисление наиболее интенсивно протекает в мышечной ткани, почках, печени.В результате b-окисления ЖК образуется Ацетил-КоА. Скорость окисления определяется скоростью процессов липолиза. Ускорение липолиза характерно для состояния углеводного голодания и интенсивной мышечной работы. Ускорение b-окисления наблюдается во многих тканях, в том числе и в печени. В печени образуется больше Ацетил-КоА, чем ей требуется. Печень - "орган-альтруист" и поэтому печень отправляет глюкозу в другие ткани.Печень стремится направить в другие ткани и свой собственный Ацетил-КоА, но не может, так как для Ацетил-КоА клеточные мембраны непроницаемы. Поэтому в печени из Ацетил-КоА синтезируются специальные вещества, которые называются "кетоновые тела". Кетоновые тела - это особая транспортная форма ацетил-КоА.
Основы генетики.
1.Достижения и перспективы генной инженерии.
Генная инженерия - экспериментальная наука. Возникла на стыке молекулярной биологии и генетики официально в 1972 г., когда в лаборатории П. Берга (Стенфордский университет, США) была получена первая рекомбинантная (гибридная) ДНК на базе объединения генетического материала, полный геном вируса обезьян 40, часть генома измерного бактериофага и гены галактозного оперона.
Генная инженерия нацелена на создание организмов с новыми комбинациями наследственных свойств путем конструирования функционально-активных генетических структур в форме рекомбинантных ДНК из фрагментов геномов разных организмов, которые вводились в клетку.
Достижения генетики и химии нуклеиновых кислот позво-лили разработать методологию генной инженерии:
-открытие явления рестрикции - модификации ДНК и выделение ферментов рестриктаз для получения специфических ферментов;
-создание методов химического и ферментативного синтеза генов;
-выявление векторных молекул ДНК, способных перенести в клетку чужеродную ДНК и обеспечить там экспрессию соответствующих генов;
- разработка методов трансформации у различных организмов и отбор клонов, несущих рекомбинантные ДНК.
ДНК представляет запись последовательности АК для построения молекул различных белков. В эмбриональном развитии в разное время появляются разные белки. Существуют гены-регуляторы, которые определяют время и скорость синтеза. Установлены состав и структура гена, но неизвестно как кодируется форма организма и, соответственно, как линейные спирали цепочной структуры белков соединяются в объемные структуры.
Клонирование - воспроизведение живого существа из его неполовых клеток. Это попытка прорыва сквозь запреты Природы. Клонирование органов и тканей - это задача номер один в области трансплантологии, травматологии и др. областях медицины и биологии: при пересадке не возникает реакции отторжения и возможных последствий (например, рака, развивающегося на фоне иммунодефицита). Клонирование может дать возможность бездетным людям иметь своих собственных детей, поможет людям, страдающим тяжелыми генетическими заболеваниями. Так, если гены, определяющие какую-либо подобную болезнь, содержатся в хромосомах отца, то в яйцеклетку матери пересаживается ядро ее собственной соматической клетки, тогда появится ребенок, ли-шенный опасных генов, точная копия матери, и наоборот. Регуляция пола сельскохозяйственных животных, клонирование в них человеческих генов "терапевтических белков", которые используются для лечения людей, например гемофи-ликов, у которых мутировал ген, кодирующий белок, участвующий в процессе свертывания крови. .
Применение методов клеточной инженерии позволяет существенно интенсифицировать процесс созда-ния новых форм организмов. Метод гибридизации соматических клеток -- новый метод, дающий возможность получать межвидовые гибриды, т.е. преодолевать естественный барьер межвидовой нескрещиваемости, чего нельзя было достичь тради-ционными методами селекции. Для этого в искусственно созданных условиях выделяют и сливают протопласты - клетки, лишенные стенок, - обоих родительских растений и получают гибридные клетки, которые могут затем регенерировать целое гибридное растение с признаками обоих родителей. Это позволяет получать совершенно новые организмы, не существовавшие в природе. Но при этом возникает опасность, что искус-ственно созданные организмы могут вызвать непредсказуемые и необратимые последствия для всего живого на Земле, в том числе, и для человека.
Достижения и перспективы генной инженерии. Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы.Первым искусственно изменённым продуктом стал помидор. Его новым свойством стала способность месяцами лежать в недоспелом виде при температуре 12 градусов. Но как только такой помидор помещают в тепло, онза несколько часов становится спелым. Американские компании Origen Therapeutics и Embrex планируют наладитьмассовое производство клонированных цыплят. Смысл всей затеи очевиден:тиражирование одной единственной жирной птички, которая мало ест, быстрорастет и не болеет, представляется делом необыкновенно выгодным.Исследования, которые проводятся при поддержке Национального институтанауки и технологий, выделившего на проект 4,7 миллиона долларов, уже даликонкретные результаты. Технология клонирования в своем обычном виде,предполагающая перенос ядра клетки-донора в яйцеклетку с последующей ееимплантацией суррогатной матери, к птицам неприменима, поскольку, какизвестно, их эмбрионы развиваются не в матке, а в скорлупе. Генетическиекопии цыплят создаются иным образом. Ученые выделяют и размножаютэмбриональные стволовые клетки донора, из которых с ростом эмбрионаразвиваются все ткани. Затем эти клетки имплантируются в обычное яйцо.Строго говоря, получающийся таким образом цыпленок является не генетическойкопией, а "химерой", поскольку вместе с донорскими клетками содержит иродные, те, что были в яйце. Однако ученые добились, чтобы донорских клетокбыло более 95 %, и даже создали 100-процентного клона. Для массовогопроизводства таких цыплят планируется использовать специальные машины,способные за час ввести инъекции в 50 тысяч яиц. Американцы добились изменения клубники, тюльпанов. Вывели сорткартофеля, который при жарке впитывает меньше жира. Они же скоро планируютполучить помидоры-гиганты кубической формы, чтобы их было легче упаковыватьв ящики. Швейцарцы начали выращивать кукурузу, которая выделяет собственныйяд против вредителей. Был создан "помидор с жабрами" - помидор, в который для увеличенияморозоустойчивости вживили ген североамериканской плоской рыбы. Кстати,именно этот гибрид овоща и рыбы получил кличку "завтрак Франкенштейна". В Московском институте картофелеводства выводится картофель счеловеческим интерфероном крови, который повышает иммунитет. А в Институтеживотноводства получен патент на овцу, у которой в молоке присутствуетсычужный фермент, необходимый для производства сыра. Специалистыутверждают, что при новой технологии производства сыра, достаточно будетвсего 200 овец, чтобы обеспечить сыром всю Россию. Сегодня ученые работают над созданием "умных растений", которые могутпосылать фермерам сигнал SOS, светиться, когда им не хватает воды или припервых признаках заболевания. Полным ходом идут работы по созданиюпластмассы, которая бы разрушалась, попадая в окружающую среду - вмасличные культуры вводят гены бактерий, позволяющие выращивать этубиоразлагаемую пластмассу прямо на полях. Недавно американцы заявили, чтоим удалось добавить в генную структуру обычного хлопка гены растений,цветущих голубым цветом. Появилась реальная возможность революционизироватьрынок джинсовой ткани - красильное производство прекратит сброс вокружающую среду ядовитых сточных вод. Эта технология будет запущена впроизводство в 2005 году. Эксперименты ведутся и в другой области - области запахов. Некоторые нелюбят запах роз, считая его слишком приторным, - для таких людей можновыращивать розы, благоухающие лимоном. Можно даже вырастить розу, издающуюаромат духов Кельвина Клайна - манипуляции с генами, отвечающими за запах,позволяют вывести растения с любым ароматом. Некоторые особенности новых технологий 21 века могут привести к большимопасностям, чем существующие средства массового уничтожения. Прежде всего,- это способность к саморепликации. Разрушающий и лавинносамовоспроизводящийся объект, специально созданный или случайно оказавшийсявне контроля, может стать средством массового поражения всех или избранных.Для этого не потребуются комплексы заводов, сложная организация и большиеассигнования. Угрозу будет представлять само знание: устройства,изобретённые и изготовленные в единичных экземплярах, могут содержать всебе всё, необходимое для дальнейшего размножения, действия и дажедальнейшей эволюции – изменению своих свойств в заданном направлении.Конечно, выше описаны вероятные, но не гарантированные варианты развитиягенной инженерии. Успех в этой отрасли науки сможет радикально поднятьпроизводительность труда и способствовать решению многих существующихпроблем, прежде всего, подъему уровня жизни каждого человека, но, в то жевремя, и создать новые разрушительные средства.
2.Закономерности наследования признаков, установленных Г.Менделем.
Часть открытий из области основных закономерностей наследования признаков принадлежит Менделю. Он проводил опыты по гибридизации гороха. Он отбирал растения, отличающиеся парой альтернативных признаков: желтая или зелёная окраска зерна, гладкая или морщинистая кожура, красные или белые цветки. Проверяя стойкое наследование взятых признаков, т.е. выбирал чистые линии.
Мендель шел в своих исследованиях от простого к сложному. Он вначале анализировал наследование одной пары признаков (моногибридное скрещивание). При скрещивании растений с желтыми и зелёными семенами в первом поколении (F1) всё потомство имело жёлтые семена (доминирующий признак). Зелёный – подавляемым (рецессивный). Один и тот же результат наблюдался при скрещивании ♂ жёлтых с ♀ зелёными, так и ♂ зелёных с ♀ желтыми. Такое единообразие гибридов F1 получило название правило доминирования или Первый закон Менделя.
При самоопылении гибридов F1 во втором поколении F2 наблюдалось расщепление 3:1, т.е. 3 жёлтых и 1 зелёный. Такая закономерность получила название Второй закон Менделя или закон расщепления.
Наблюдая описанные явления Мендель приходит к выводу, что за наследование признаков отвечает пара наследственных задатков (генов). При изучении наследования одного или нескольких признаков говорят о генотипе или о фенотипе конкретных признаков. Гены могут находиться в организме в гомозиготном (как по доминантным, так и по рецессивным признакам (AA, aa)) и гетерозиготном состоянии (Aa), образуя аллельные пары. Аллельные пары – гены, занимающие идентичное положение в гомологичных хромосомах и ответственные за проявление одного и того же признака. При образовании гамет в каждую из них попадает только по одному из пары наследственных факторов, определяющих признак. Это положение было высказано Бэтсоном в виде «гипотезы чистоты гамет». Для лучшего понимания Пеннет предложил использовать решётку или таблицу (решетка Пеннета). Анализ полученных данных показывает, что расщепление 3:1 – расщепление по фенотипу, а по генотипу 1:2:1. Отсюда следует, что сходные по фенотипу особи могут иметь разный генотип. И что бы проверить является ли гибрид гомозиготный или гетерозиготный проводят анализирующие скрещивание (с гомозиготой по рецессивному признаку). Если при таком скрещивании всё потомство будет однотипным, значит мы имели дело с гомозиготой. Так же следует помнить, что второй закон Менделя носит статистический характер. Сам Мендель получил 3:0,99.
При изучении результатов дигибридного скрещивания, т.е. по 2 парам альтернативных признаков, Мендель наблюдал независимое наследование признаков (9:3:3:1). При полигибридном скрещивании расщепление можно выразить по формуле (3+1)n , где n – число пар альтернативных признаков. Закон независимого расщепления по парам неаллельных признаков – 3 закон Менделя.
Причина успеха Менделя в правильном выборе объекта, в разработке и применении принципа гибридологического анализа.
Его метод гибридологического анализа используется в генетике и сейчас.
Организмы должны быть одного вида
Чётко различаться по отдельным признакам
Признаки должны быть константными, т.е. свободно передаваться из поколения в поколение.
Необходима характеристика и количественный учёт всех классов расщепления потомства в первом и последующих поколениях.
То, что признаки не исчезают в F1 , а проявляются вновь в последующих поколениях, позволило Менделю сформулировать гипотезу о дискретном характере вещества наследственности.
Установленные Менделем закономерности наследования признаков получили цитологические обоснования после открытия хромосом.
3.Генотип как система взаимодействующих генов.
Генотип – совокупность генов отдельного организма, находящихся между собой в различного рода взаимодействиях. Это генетическая конституция организма, имеющая фенотипическое проявление.
Свойства генов. Существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
-Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
-Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
-Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
-Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
-Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) — это множественные гены, или полигены.
-Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
-Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование - тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля. (пурпурная окраска цветков гороха над белой, гладкая формы семян над морщинистой, темные волосы над светлыми, карие глаза над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака. Пример: наследование окраски цветков у растений ночной красавицы. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).
Также: курчавость волос у человека, масть КРС, окраска оперения у кур.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса.
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи: наследование IV группы крови у человека (группа АВ).
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ, то
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые.
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16.
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический. Доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипическим выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2 .
Тогда все генотипы (А1 ,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растения а1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
Также: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
4.Типы взаимодействия аллельных и неаллельных генов, их молекулярные механизмы.
Различают следующие основные типы взаимодействия генов:
1) комплементарность; 2) элистаз; 3) полимерия; 4) модифицирующее действие генов.
Комплементарное действие генов. Гены называют комплементарными, когда они по своему проявлению как бы дополняют друг друга. Каждый такой ген в отдельности, сам по себе, не оказывает действия, не вызывает появления признака. Однако при скрещивании двух особей, из которых каждая имеет такого типа ген (например, одна имеет ген А, а другая ген В), получается гибрид, новый организм, у которого в наличии оказываются оба этих гена, и эффект их совместного действия проявляется в том, что под их совместным влиянием у гибридного организма возникает какой-то новый признак. Например, Куры с розовидным гребнем и меют генотипАвв, с гороховидным – ааВ, с листовидным гребнем – аавв. При скрещивании кур с розовидным и гороховидным гребнем получается новый признак – ореховидный гребень.
Эпистатическое действие генов (эпистаз)
Эпистатическое действие генов по своему характеру противоположно комплементарному действию генов. Сущность его состоит в подавлении генами супрессорами действия гипостатических генов. Явление эпистаза выражается в неаллельном подавлении действия одного доминантного гена другим доминантным геном, принадлежащим к другой аллеломорфной паре.
Гены, которые подавляют действие других неаллельных им (принадлежащих к другим парам) генов, называют эпистатичными.
Эпистаз выражается в изменении соотношения расщепления во втором поколении, которое по фенотипу отклоняется от обычного расщепления при дигибридном скрещивании, т. е. от нормы Менделя 9:3:3: 1.
Рассмотрим явление эпистаза схематически. Существуют два разных по окраске типа лошадей: доминантный ген С характеризует серую окраску, а доминантный ген В — черную, вороную.
При скрещивании генотипы родителей будут: ССвв (серая) × ссВВ (вороная), а генотип F1 CcBe.
Окраска потомства F1 будет серая, так как ген С (доминант) эпистатически подавляет проявление гена В вороной окраски.
Скрестив между собой эти генотипы, т. е. Сс × Вв, будем иметь в F2 расщепление по фенотипам, 12 : 3 : 1.
Мы видим, что: 1) все зиготы, имеющие доминантный ген С, дают серых лошадей, так как ген С серой окраски эпистатичен доминантному гену В вороной окраски; 2) все зиготы с рецессивом с и доминантом В дадут вороную окраску, так как рецессивный ген с не подавляет действия доминантного гена В; 3) двойной рецессив ссвв т. е. форма, гомозиготная по обоим рецессивным генам, дает форму, отличную по фенотипу от форм с доминантными аллелями двух генов С и В, а также от форм с одной из таких аллелей (ссВВ, ССвв). В данном случае это отличие по фенотипу выражено в рыжей окраске лошади.
Полимерное действие генов (полимерия)
Полимерия представляет собой явление взаимодействия генов, при котором несколько однотипных (однозначных) генов оказывают сходное воздействие на развитие одного и того же признака.
Иначе говоря, полимерия обусловливается действием разного числа однозначных генов, которые, суммируясь, усиливают проявление признака, а при меньшем числе таких генов этот признак проявляется в соответственно меньшей степени.
С явлением полимерии приходится сталкиваться при изучении так называемых количественных признаков.
Такие признаки, как, например, вес животного, яйценоскость кур, количество белка, в эндосперме зерна кукурузы и зерна пшеницы, содержание витаминов в растениях, скорость протекания биохимических реакций и т. п., нельзя разложить на четкие фенотипические классы; их необходимо оценивать и измерять количественно. Такие признаки называются количественными, или мерными.
Изучение наследования количественно варьирующих признаков у различных особей одного и того же поколения было начато в первом десятилетии XX в. Шведский генетик Нильсон-Эле в 1908 г., скрещивая расы пшеницы, имеющие красные и белые зерна, обнаружил в первом поколении F1 обычное моногибридное расщепление 3:1. Однако при скрещивании некоторых линий пшениц с такими признаками он во втором поколении (F2) в 1910 г. получил расщепление в соотношении 15/16 окрашенных и 1/16 белых. Окраска зерен у первой группы, т. е. у 15/16 растений, варьировала от темно-красной до бледно-красной.
По типу полимерии, т. е. полимерных генов, наследуется цвет кожи у человека. Так, от брака негра и белой женщины рождаются дети с промежуточным цветом кожи (мулаты). А у супружеской пары двух мулатов рождаются дети всех возможных типов при комбинации двух неаллельных полимерных генов - от черной до белой кожи.
Изучение полимерных (множественных) генов имеет большое значение, так как очень многие хозяйственно ценные признаки у растений и животных наследуются по типу полимерии (например, содержание сахара в корнеплодах свеклы, длина початка кукурузы и т. д.).