

40. Доказательства природы выделяющегося кислорода. Роль фоторазложения воды.
Фотосинтез – это процесс, в ходе которого выделяется кислород. До появления на нашей планете первых живых организмов первичная атмосфера Земли не содержала кислорода и была похожа по химическому составу на атмосферу Венеры и Марса. Кислород стал выделяться в атмосферу Земли только с появлением первых фотосинтезирующих организмов. Постепенно в результате накопления кислорода в атмосфере условия существования на нашей планете радикально изменились. Во-первых, кислород позволил выйти на сушу животным и дышать кислородом воздуха. Во-вторых, на высоте около 30-ти километров кислород образовал озоновый экран, эффективно защищающий все живое планеты от губительного жесткого ультрафиолетового излучения Солнца.
1. Фотолиз воды . Это ферментативное расщепление воды под действием света. Универсальным способом образования АТФ является механизм нециклического фотофосфорилирования. Энергия света, поглощенная пигментами, преобразуется в энергию электронов. Свободные электроны образуются при фотолизе (фотоокислении) воды – расщеплении молекулы Н2О с затратой световой энергии. При фотолизе воды выделяется молекулярный кислород. Энергия электронов используется для создания протонных резервуаров внутри тилакоидов и формирования электрохимических потенциалов на мембранах тилакоидов. В свою очередь, энергия электрохимического потенциала используется для синтеза АТФ. Электроны, потерявшие энергию, используются для восстановления НАДФ.
4Н2О+энергия света=4Н + +4ОН - 4ОН - →2Н 2 О+4ē +О 2 ↑ 4Н + +4ē+4ĶАДФ + =4НАДФХН
41. Темновые реакции фотосинтеза (биохимический этап). Использование атф и надфн в темповых реакциях.
В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NАDРН. Эти конечные продукты световой фазы фотосинтеза стоят на входе в темно-вую фазу, где С02 восстанавливается до углевода:
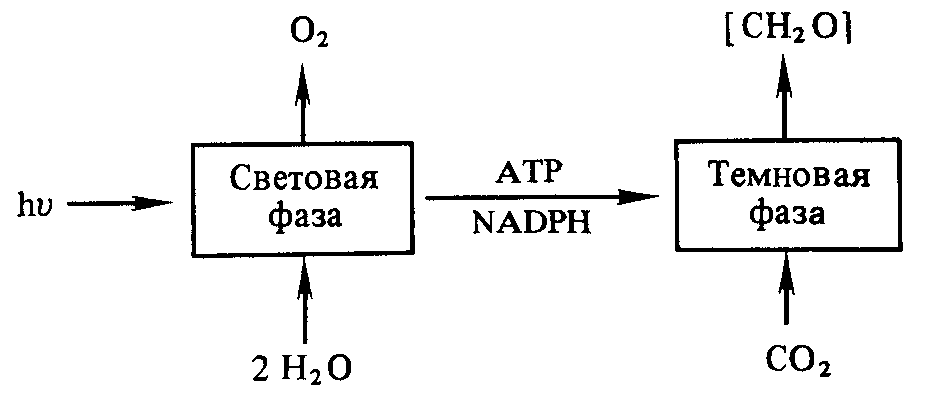
Однако сами по себе АТР и NАDРН не в состоянии восстановить С02. Очевидно, и темновая фаза фотосинтеза — сложный процесс, включающий большое количество реакций. Кроме того, существуют различные пути восстановления С02. В настоящее время известны так называемые С3-путь и С4-путь фиксации С02, фотосинтез по типу толстянковых (САМ-метаболизм) и фотодыхание. Рассмотрим каждый из этих путей в отдельности.
1. С3-путь фотосинтеза или цикл Кальвина
Этот путь ассимиляции СО2 обнаружен у всех фотосинтезирующих растений. Он был назван циклом Кальвина в честь американского биохимика М. Кальвина, который с сотрудниками открыл и изучил его в 1946-1956 годах с помощью метода меченых атомов и хроматографии. Растения разное время подкармливали 14СО2 и хроматографировали экстракты. Радиоактивные изотопы по химическим свойствам не отличаются от стабильных. Принимая участие в реакциях, они помечают те соединения, в которые входят. Цикл состоит из трех этапов: карбоксилирования, восстановления, регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза.
1. Карбоксилирование. Фосфорибулокиназа фосфорилирует при участии АТФ рибулозо-5-фосфат с образованием АДФ и рибулозо-1,5-дифосфата. Последний является акцептором СО2 и под действием рибулозодифосфаткарбоксилазы присоединяет СО2. В результате образуются 2 молекулы 3-фосфоглицериновой кислоты (3-ФГК).
2. Восстановление. Фосфоглицераткиназа при участии АТФ фосфорилирует 3-ФГК и образовавшаяся 1,3-дифосфоглицериновая кислота восстанавливается с помощью НАДФН и дегидрогеназы фосфоглицеринового альдегида до 3-фосфоглицеринового альдегида (3-ФГА).
3. Регенерация. После фиксации трех молекул СО2 и образования шести молекул 3-ФГА пять из них используются для синтеза рибулозо-5-фосфата, а одна молекула 3-ФГА – для образования глюкозы.
Триозофосфатизомераза превращает 3-ФГА в фосфодиоксиацетон. Затем альдолаза образует из 3-ФГА и фосфодиоксиацетона фруктозо-1,6-дифосфат. Он теряет один остаток фосфорной кислоты под влиянием фруктозо-1,6-дифосфатазы и превращается во фруктозо-6-фосфат. Транскетолаза переносит гликолевый альдегид от фруктозо-6-фосфата на 3-ФГА с образованием эритрозо-4-фосфата и ксилулозо-5-фосфата. Альдолаза присоединяет фосфодиоксиацетон к эритрозо-4-фосфату с образованием седогептулозо-1,7-дифосфата. Он дефосфорилируется фосфатазой и под влиянием транскетолазы соединяется с 3-ФГА. Продуктами этой реакции являются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата под действием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата.
Ш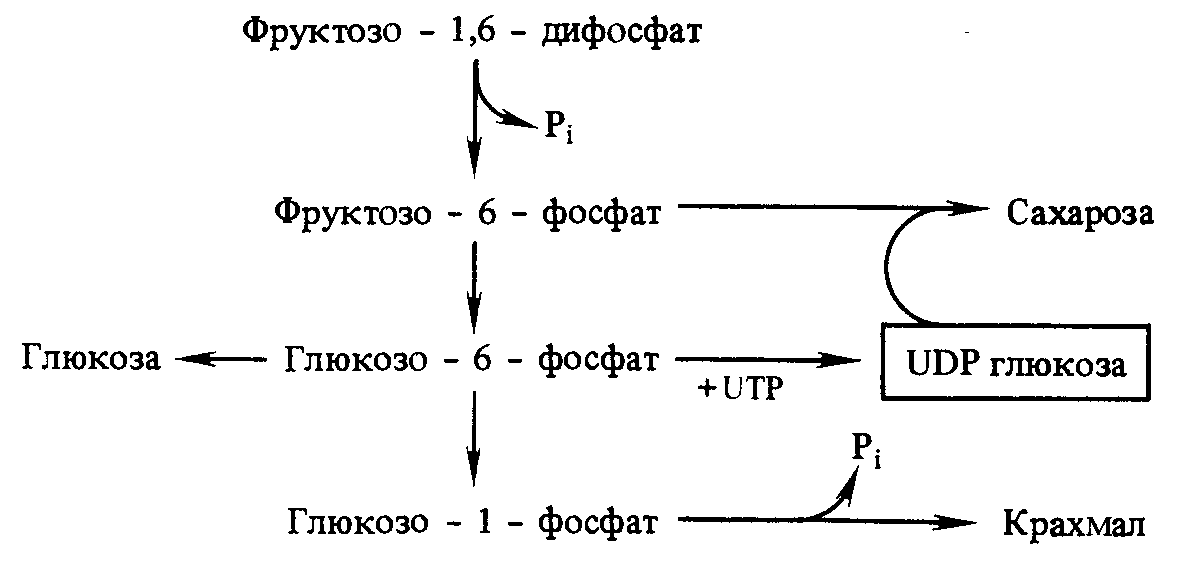 естая
молекула 3-ФГА используется для синтеза
фруктозо-1,6-дифосфата при повторении
цикла. Из двух молекул фруктозо-1,6-дифосфата
образуются фруктозо-6-фосфат и
глюкозо-1-фосфат. Последний, взаимодействуя
с уридинтрифосфатом, дает уридинфосфоглюкозу.
Она и фруктозо-6-фосфат образуют
сахарозофосфат, который после
дефосфорилирования превращается в
сахарозу. Следовательно, для образования
одной молекулы сахарозы необходимо
прохождение четырех циклов Кальвина.
Крахмал синтезируется из уридинфосфоглюкозы
под влиянием амилосинтетазы.
естая
молекула 3-ФГА используется для синтеза
фруктозо-1,6-дифосфата при повторении
цикла. Из двух молекул фруктозо-1,6-дифосфата
образуются фруктозо-6-фосфат и
глюкозо-1-фосфат. Последний, взаимодействуя
с уридинтрифосфатом, дает уридинфосфоглюкозу.
Она и фруктозо-6-фосфат образуют
сахарозофосфат, который после
дефосфорилирования превращается в
сахарозу. Следовательно, для образования
одной молекулы сахарозы необходимо
прохождение четырех циклов Кальвина.
Крахмал синтезируется из уридинфосфоглюкозы
под влиянием амилосинтетазы.
Среди продуктов фотосинтеза обнаружены аминокислоты. При недостатке НАДФН 3-ФГК превращается не в 3-ФГА, а в пировиноградную кислоту. Она, присоединяя аммиак, образует аланин. Из пировиноградной кислоты в цикле Кребса образуются органические кислоты, которые в ходе реакций аминирования и переаминирования дают аминокислоты.