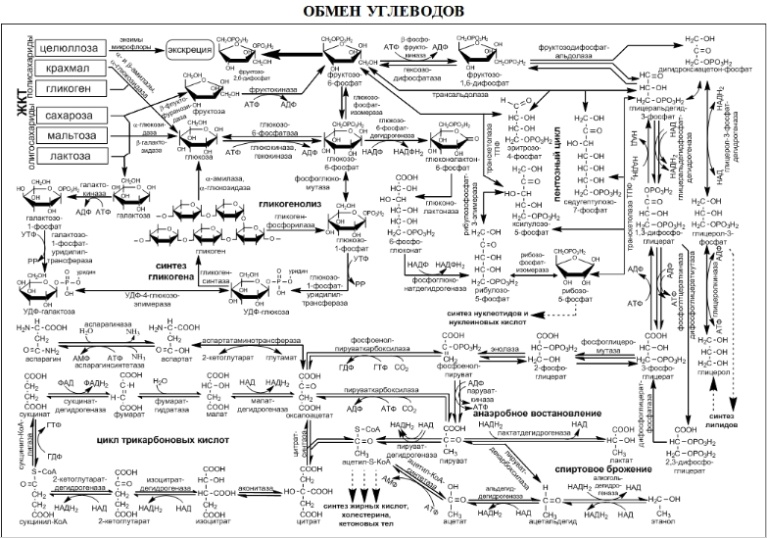
45. Цикл трикарбоновых кислот. Окислительное фосфорилирование на уровне субстрата. Прямое окисление глюкозо-6-фосфата.
Цикл трикарбоновых кислот (цикл Кребса, цитратный цикл) — центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ. Цикл Кребса — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др. У эукариот все реакции цикла Кребса протекают внутри митохондрий, причём катализирующие их ферменты, кроме одного, находятся в свободном состоянии в митохондриальном матриксе, исключение составляет сукцинатдегидрогеназа, которая локализуется на внутренней митохондриальной мембране, встраиваясь в липидный бислой. У прокариот реакции цикла протекают в цитоплазме. При работе цикла Кребса окисляются различные продукты обмена, в частности токсичные недоокисленные продукты распада алкоголя, поэтому стимуляцию цикла Кребса можно рассматривать как меру биохимической детоксикации. Цикл Кребса регулируется «по механизму отрицательной обратной связи», при наличии большого количества субстратов (ацетил-КоА, оксалоацетат), цикл активно работает, а при избытке продуктов реакции (NADH, ATP) тормозится. Регуляция осуществляется и при помощи гормонов, основным источником ацетил-КоА является глюкоза, поэтому гормоны, способствующие аэробному распаду глюкозы, способствуют работе цикла Кребса. Такими гормонами являются: инсулин и адреналин. Глюкагон стимулирует синтез глюкозы и ингибирует реакции цикла Кребса. Как правило работа цикла Кребса не прерывается за счёт анаплеротических реакций, которые пополняют цикл субстратами: Пируват + СО2 + АТФ = Оксалацетат(субстрат Цикла Кребса) + АДФ + Фн. Интегративная функция — цикл является связующим звеном между реакциями анаболизма и катаболизма. Катаболическая функция — превращение различных веществ в субстраты цикла: Жирные кислоты, пируват,Лей,Фен — Ацетил-КоА; Арг, Гис, Глу — α-кетоглутарат; Фен, тир — фумарат. Анаболическая функция — использование субстратов цикла на синтез органических веществ: Оксалацетат — глюкоза, Асп, Асн; Сукцинил-КоА — синтез гемма; CО2 — реакции карбоксилирования. Водорододонорная функция — цикл Кребса поставляет на дыхательную цепь митохондрий протоны в виде трех НАДН.Н+ и одного ФАДН2. Энергетическая функция — 3 НАДН.Н+ дает 7.5 моль АТФ, 1 ФАДН2 дает 1.5 моль АТФ на дыхательной цепи. Кроме того в цикле путем субстратного фосфорилирования синтезируется 1 ГТФ, а затем из него синтезируется АТФ посредствам трансфосфорилирования: ГТФ + АДФ = АТФ + ГДФ.
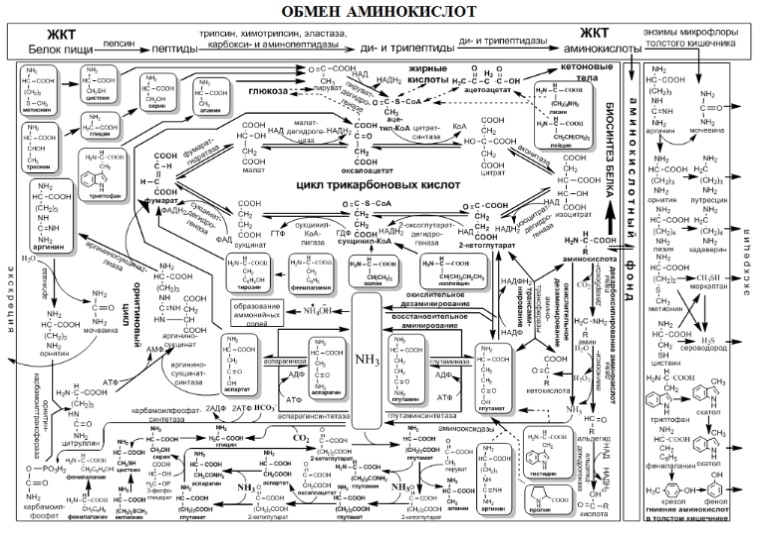
46.Пентозофосфатный путь обмена углеводов, его биологическая роль. Пентозофосфатный (фосфоглюконатный) путь обеспечивает клетку гидрированным НАДФ для восстановительных синтезов и пентозами для синтеза нуклеотидов. Следовательно, этот процесс выполняет анаболитические функции. В пентозофосфатном пути можно выделить две стадии – окислительный и неокислительный пути образования пентоз. Пентозофосфатный цикл осуществляется в цитозоле (жидкой фазе) клеток животных, растений (особенно в темноте) и микроорганизмов. У растений часть р-ций пентозофосфатного цикла участвует также в образовании гексоз при фотосинтезе. Регуляция направленности р-ций в пентозофосфатном цикле осуществляется гл. обр. ферментами, участвующими в этом цикле: избыток того или иного субстрата подавляет активность фермента, катализирующего его синтез, или активирует фермент, катализирующий его трансформацию в др. соединение. Относит. кол-ва глюкозы, превращающиеся через пентозофосфатный цикл, неодинаковы в разных тканях. В мышцах скорость пентозофосфатного цикла очень низка, а в печени не менее 30% CO2 образуется при окислении глюкозы в пентозофосфатном цикле. В др. тканях, где активно проходит биосинтез жирных к-т и стероидов (семенниках, жировой ткани, лейкоцитах, коре надпочечников, молочной железе), доля пентозофосфатного цикла в окислит. метаболизме глюкозы также очень значительна. Интенсивность пентозофосфатного цикла зависит от функцион. состояния ткани и от гормонального статуса (напр., в печени резко снижается при голодании из-за инактивации дегидрогеназ пентозофосфатного цикла и восстанавливается вскоре после кормления). Скорость пентозофосфатного цикла регулируется в первую очередь концентрацией НАДФН. Обе дегидрогеназы пентозофосфатного цикла (р-ции 1 и 3) чувствительны к изменению величины отношения НАДФ/НАДФН: при его величине 0,02 активность дегидрогеназ в печени максимальна, а при величине 0,01 снижается на 90%. Интенсивный пентозофосфатный цикл происходит в эритроцитах, что связано с необходимостью НАДФН-зависимого восстановления глутатиона кофактора глутатионредуктазы эритроцитов. Нарушения функционирования нек-рых ферментов пентозофосфатного цикла приводят к развитию тяжелых заболеваний человека. Недостаточность глюкозо-6-фосфат-дегидрогеназы в эритроцитах служит причиной лек. гемолитич. анемии, а снижение активности транскетолазы в результате нарушения ее способности связывать тиамин приводит к развитию нервно-психич. расстройства синдрома Вернике Корсакова. Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пиримидиновых нуклеотидов и гидрированным ко-ферментом NADPH, который используется в восстановительных процессах. Протекание пентозофосфатного цикла позволяет клеткам продуцировать NADPH, необходимый для синтеза жиров, не накапливая пентозы. Энергия, выделяющаяся при распаде глюкозы, трансформируется в энергию высокоэнергетического донора водорода - NADPH. Гидрированный NADPH служит источником водорода для восстановительных синтезов, а энергия NADPH преобразуется и сохраняется во вновь синтезированных веществах.