

Вопрос 24. Синаптическая передача нервного импульса.
Синаптическая передача (также называемая нейропередача) — электрические движения в синапсах вызванные распространением нервных импульсов. Каждая нервная клетка получает медиатор из пресинаптического нейрона или из терминального окончания или из постсинаптичекского нейрона или дендрида вторичного нейрона и посылает его обратно нескольким нейронам, которые повторяют данный процесс, таким образом, распространяя волну импульсов до тех пор, пока импульс не достигнет опроделенного органа или специфической группы нейронов.
Нервные импульсы необходимы для распространения сигналов. Эти сигналы посылаются в и исходят из центральной нервной системы через эфферентные и афферентные нейроны для координации гладких, скелетных и сердечных мышц, секреции желез и функционирования органов, важных для долгосрочного выживания многоклеточных позвоночных организмов, таких как млекопитающие.
Нейроны образуют нейронные сети, по которым передаются нервные импульсы. Каждый нейрон образует не менее 15,000 соединений с другими нейронами. Нейроны не соприкасаются друг с другом; они образуют точки соприкосновения, называемые синапсами. Нейрон передают информацию с помощью нервного импульса. Когда импульс нейрона достигает синапса это приводит к выделению медиаторов, которые влияют на другие клетки, приводя к угнетению или облегчению. Следующий нейрон может соединяться с множеством других нейронов, и если возбуждающие процессы превалируют над угнетающими, то будет развит потенциал действия в основании аксона, таким образом передавая информацию к следующему нейрону, приводя к памяти или действию.
Примером распространения с помощью нейронов является сердечное сокращение. Сокращение осуществляется когда сигнал поступает из синоатриального узла с частотой, заставляющей сердце полностью сократиться, выбросив всю кровь и последующему наполнению новой порцией крови. Важно, что что импульс опосылается из синоатриального узла, так как направление импульса между нейронами заставляет мышцу сокращаться полностью. Если импульс будет поступать из синоатриального узла сердце будет сокращаться неуверенно и не будет выбрасывать всю кровь в систему.
Вопрос №25. Межклеточные соединения. Плотные или замыкающие контакты
Шпора 10
В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания.
Вопрос №26.метаболический аппарат клетки. Этапы внутриклеточного транспорта.
МЕТАБОЛИЗМ - это обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода. Ассимиля́ция — совокупность процессов анаболизма (биосинтеза) в живом организме, в ходе которых различные вещества включаются в его состав. В ходе ассимиляции простые вещества (сложные первоначально расщепляются до простых), неспецифические для какого-либо организма, превращаются в сложные, характерные для данного вида соединения (усваиваются). Ассимиляция уравновешивается суммой процессов диссимиляции (распада). Метаболический аппарат клетки - совокупность ее структур, участвующих в метаболизме. Органоиды, участвующие в ассимиляции: шЭПС- На каналах шероховатой ЭПС в виде полисом расположены рибосомы. Здесь протекает синтез белков, преимущественно продуцируемых клеткой на экспорт (удаление ич клегки), например, секретов железистых клеток. Здесь же происходят образование липи-дов и белков цитоплазматической мембраны и их сборка. Плотно упакованные цистерны и каналы гранулярной ЭПС образуют слоистую структуру, где наиболее активно протекает синтез белка. Это место называется эргастоплазмой. Гладкая ЭПС-на мембранах гладкой ЭПС рибосом нет. Здесь протекает в основном синтез жиров и подобных им веществ (например, стероидных гормонов), а также углеводов. По каналам гладкой ЭПС также происходит перемещение готового материала к месту его упаковки в гранулы (в зону комплекса Гольджи). В печеночных клетках гладкая ЭПС принимает участие в разрушении и обезвреживании ряда токсичных и лекарственных веществ (например, барбитуратов). В поперечно-полосатой мускулатуре канальцы и цистерны гладкой ЭПС депонируют ионы кальцин. Комплекс Гольджи- пластинчатый комплекс Гольджи что упаковочный центр клетки. Представляет собой совокупность дик тиосом (от нескольких десятков до сотен и тысяч па одну клетку). Дикгиосома— стопка из 3—12 уплощенных цистерн овальной формы, по краям которых расположены мелкие пузырьки (везикулы). Более крупные расширения цистерн дают вакуоли, содержащие резерв воды в клетке и отвечающие за поддержание тургора. Пластинчатый комплекс дает начало секреторным вакуолям, и которых содержатся вещества, предназначенные для вывода из клетки. 11ри этом просекрет, поступающий в вакуоль из зоны синтеза, (')ПС, митохондрии, рибосомы), подвергается здесь некоторым химическим превращениям. Комплекс Гольджи дает начало первичным лизосомам. В диктиосомах также синтезируются полисахариды, гликопротеиды и гликолиииды, которые затем идут на построение цитоплазматических мембран. Рибосома — важнейший немембранный органоид живой клетки сферической или слегка эллипсоидной формы, диаметром 100—200 ангстрем, состоящий из большой и малой субъединиц. Ф-ции: биосинтеза белка из а/к по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией. Рибосомы располагаются на мембранах эндоплазматического ретикулума, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Рибосомы представляют собой нуклеопротеид, в составе которого отношение РНК/белок составляет 1:1 у высших животных и 60-65:35-40 у бактерий. Рибосомная РНК составляет около 70 % всей РНК клетки. Рибосомы эукариот включают четыре молекулы рРНК, из них 18S, 5.8S и 28S рРНК синтезируются в ядрышке РНК полимеразой I в виде единого предшественника (45S), который затем подвергается модификациям и нарезанию. 5S рРНК синтезируется РНК полимеразой III в другой части генома и не нуждаются в дополнительных модификациях. Почти вся рРНК находится в виде магниевой соли, что необходимо для поддержания структуры; при удалении ионов магния рибосома подвергается диссоциации на субъединицы.
Константа седиментации (скорость оседания в ультрацентрифуге) рибосом эукариотических клеток равняется 80S (большая и малая субъединицы 60S и 40S, соответственно), бактериальных клеток (а также митохондрий и пластид) — 70S (большая и малая субъединицы 50S и 30S, соответственно).
Вопрос №27. Современные представления о механизме внутриклеточного транспорта.
Потоки информации, энергии и веществ осуществляются непрерывно и составляют необходимое условие сохранения клетки как живой системы.
Кроме структур и процессов, прямо включенных в названные потоки, в клетке функционируют механизмы, которые также являются жизненно необходимыми. Так, лизосомы, воздействуя ферментами на пиноцитированный или аутофагированный материал, обеспечивают гидролитическое расщепление макромолекул до низкомолекулярных соединений. Они же обусловливают разрушение внутриклеточных структур, утративших свое функциональное значение. Образовавшиеся при этом химические соединения включаются в потоки энергии, веществ и информации. Пероксисомы ликвидируют возникающие в клетке пероксиды, токсичные для живой протоплазмы. Организация внутриклеточных транспортных потоков обусловливается наличием и активностью микротрубочек, микрофибрилл.
Вопрос №28. Внутриклеточный поток энергии
Поток энергии у представителей разных групп организмов обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов концентрации веществ), электрическую, механическую, ре-гуляторную. Макроэргическим называют соединение, в химических связях которого запасена энергия в форме, доступной для использования в биологических процессах. Универсальным соединением такого рода служит АТФ. Основное количество энергии заключено в связи, присоединяющей третий остаток фосфорной кислоты.
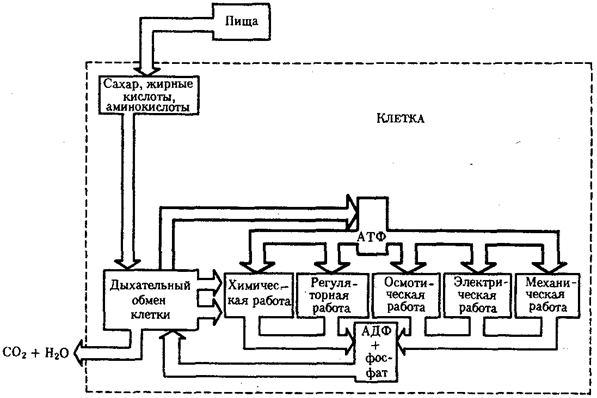
Среди органелл животной клетки особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования, а также матриксу цитоплазмы, в котором протекает процесс бескислородного расщепления глюкозы — анаэробный гликолиз (рис. 2.8). Из двух механизмов, обеспечивающих жизнедеятельность клетки энергией, анаэробный гликолиз менее эффективен. В связи с неполным (в отсутствие кислорода) окислением, прежде всего глюкозы, в процессе гликолиза для нужд клетки извлекается не более 10% энергии. Недоокисленные продукты гликолиза (пируват) поступают в митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию.
Из преобразователей энергии химических связей АТФ в работу наиболее изучена механохимическая система поперечно-полосатой мышцы. Она состоит из сократительных белков (актомиозиновый комплекс) и фермента аденозинтрифосфатазы, расщепляющего АТФ с высвобождением энергии.
Особенность потока энергии растительной клетки состоит в наличии фотосинтеза — механизма преобразования энергии солнечного света в энергию химических связей органических веществ.
Механизмы энергообеспечения клетки отличаются эффективностью. Коэффициенты полезного действия хлоропласта и митохондрий, достигая соответственно 25 и 45—60%, существенно превосходят аналогичный показатель паровой машины (8%) или двигателя внутреннего сгорания (17%).
Вопрос №29. Гипотеза эволюционного происхождения митохондрий, хлоропластов, ядра,ЭПР. Возникновение клеточной организации в ходе эволюции.
На основании сходства бактерий с митохондриями и хлоропластами эукариотических клеток можно предположить, что митохондрии и хлоропласты произошли от бактерий, которые нашли себе «убежище» в более крупных гетеротрофных клетках эукариот. Бактерии имели возможность использовать молекулярный кислород для окисления питательных веществ и использовать энергию света. Более крупные клетки ? хозяева использовали эти полезные свойства и имели с такими помощниками явное преимущество перед своими современниками. Все ныне живущие эукариоты, за малым исключением, содержат митохондрии, а все автотрофные эукариоты содержат также хлоропласты. По-видимому, они были приобретены в результате независимых случаев симбиоза. Более крупные клетки эукариот защищали свои симбиотические органеллы от неблагоприятных воздействий.
Этиопласты образуются у растений, выращиваемых в темноте, они имеются, например, у проростков, расположенных в почве, до их выхода на дневную поверхность. Этиопласты занимают промежуточное положение между пропластидами и настоящими хлоропластами. Для них характерно хорошо развитое проламеллярное тело с кристаллической структурой. На свету этиопласты тут же превращаются в зрелые хлоропласты.
Хромопласты – это пигментированные окрашенные пластиды, но в отличие от хлоропластов, они не содержат хлорофиллов, а синтезируют и накапливают каротиноиды. Каротиноиды придают этим пластидам желтую, оранжевую и красную окраску.
При этом каротиноиды синтезируются не на поверхности внутренних мембран, а в строме хромопластов. Как правило, каротиноиды растворены в жирных маслах пластоглобул. Внутренняя мембранная система у хромопластов либо не развита, либо деградировала.
Форма хромопластов весьма разнообразна. Они придают яркую окраску лепесткам цветов, зрелым плодам. Это имеет явное приспособительное значение.
Хромопласты обычно возникают из хлоропластов, реже из лейкопластов. По целому ряду признаков их можно назвать стареющими пластидами. Старение хлоропластов происходит, например, при созревании фруктов. Массовое старение хлоропластов наблюдается при пожелтении листьев осенью.