

Раздел III
№1
Индивидуальное развитие
Онтогенез – это индивидуальное развитие организма.
Существует несколько типов онтогенеза. Перечислим некоторые из них:
Онтогенез организмов с бесполым размножением и/или при зиготном мейозе (прокариоты и некоторые низшие эукариоты).
Онтогенез организмов с чередованием ядерных фаз при споровом мейозе (большинство растений и грибов).
Онтогенез животных с чередованием полового и бесполого размножения без смены ядерных фаз. Чередование поколений с половым и бесполым размножением у Кишечнополостных называется метагенезом. Чередование партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых называется гетерогонией.
Онтогенез животных с наличием личиночных и промежуточных стадий: от первично-личиночного анаморфоза до полного метаморфоза.
Онтогенез животных с утратой личиночных стадий и/или стадий бесполого размножения: пресноводные гидры, олигохеты, наземные и вторично-водные брюхоногие моллюски.
Онтогенез животных с утратой конечных стадий и размножением на ранних этапах онтогенеза; проявляется в виде педоморфозов (сохранении личиночных черт) и неотении (размножения на личиночной стадии).
Прямое развитие — развитие, при котором появившийся организм идентичен по строению взрослому организму, но имеет меньшие размеры и не обладает половой зрелостью. Дальнейшее развитие связано с увеличением размеров и приобретением половой зрелости. Например: развитие рептилий, птиц, млекопитающих.
Непрямое развитие, или развитие с метаморфозом — появившийся организм отличается по строению от взрослого организма, обычно устроен проще, может иметь специфические органы, такой зародыш называется личинкой. Личинка питается, растет и со временем личиночные органы заменяются органами, свойственными взрослому организму (имаго). Например: развитие лягушки, некоторых насекомых, различных червей.
Существует несколько классификаций периодов онтогенеза. Например, классификация по временным отрезкам онтогенеза со способностью особи осуществлять функцию размножения. Тогда его можно разделить на три периода: дорепродуктивный, репродуктивный и пострепродуктивный.
В дорепродуктивном периоде особь не способна к размножению. Основное содержание его заключается в развитии зрелого в половом отношении фенотипа. В этом периоде происходят наиболее выраженные структурные и функциональные преобразования, реализуется основная часть наследственной информации, организм обладает высокой чувствительностью ко всевозможным воздействиям.
В репродуктивном периоде особь осуществляет функцию полового размножения, отличается наиболее стабильным функционированием органов и систем, а также относительной устойчивостью к воздействиям.
Пострепродуктивный период связан со старением организма и характеризуется ослаблением или полным прекращением участия в размножении. Снижаются приспособительные возможности и устойчивость к разнообразным воздействиям. Применительно к онтогенезу человека названные периоды дополнительно характеризуются специфическими социальными моментами (образование, трудоспособность, творчество). Для каждого из указанных периодов характерны свои особенности заболеваемости.
Дорепродуктивный период подразделяют еще на четыре периода: эмбриональный, личиночный, метаморфоз и ювенильный.
Эмбриональный, или зародышевый, период онтогенеза начинается с момента оплодотворения и продолжается до выхода зародыша из яйцевых оболочек. Эмбриональный период отличается выраженностью процессов преобразования зиготы в организм, способный к более или менее самостоятельному существованию. У большинства позвоночных он включает стадии (фазы) дробления, гаструляции, а также гисто- и органогенеза. Продолжительность его бывает различна. У плацентарных млекопитающих он особенно укорочен. Единственная яйцевая оболочка растворяется перед имплантацией бластоцисты в слизистую матки. Зародыш к этому моменту успевает пройти только стадии зиготы и дробления. Все дальнейшие процессы протекают под защитой и при участии материнского организма. Эволюционное значение этих особенностей будет рассмотрено ниже.
Дроблению предшествуют процессы гаметогенеза и оплодотворения, которые не являются непосредственно индивидуальным развитием и могут даже не привести к нему, но которые во многом определяют дальнейшее развитие зародыша в том случае, если зачатие состоится. Эти процессы называют прогенезом, предшествующим собственно онтогенезу. Цитологически процессы гаметогенеза и оплодотворения представляют собой промежуточное звено, связывающее онтогенезы родителей с онтогенезом их потомства.
Личиночный период в типичном варианте наблюдается в развитии тех позвоночных, зародыши которых выходят из яйцевых оболочек и начинают вести самостоятельный образ жизни, не достигнув дефинитивных (зрелых) черт организации. Так, он встречается у некоторых представителей низших позвоночных — миног, большинства костистых рыб и земноводных. Наиболее характерными чертами личинки являются эмбриональный характер ее организации, наличие временных (провизорных) органов, раннее начало функционирования ряда органов, дающее возможность самостоятельного существования. Благодаря активному питанию личинка получает возможность завершить развитие, а благодаря активному перемещению имеет возможность выбирать условия среды, оптимальные для развития, и выйти, таким образом, из конкуренции со своими же взрослыми сородичами. Продолжительность личиночного периода в сравнении с эмбриональным у всех позвоночных существенно больше.
Метаморфоз состоит в превращении личинки в ювенильную форму. В процессе метаморфоза происходят такие важные морфогенетические преобразования, как частичное разрушение, перестройка и новообразование органов. Степень преобразований тем больше, чем больше различия между средой обитания личинки и взрослого организма, что хорошо иллюстрирует пример развития бесхвостных амфибий.
Ювенильный период начинается с момента завершения метаморфоза и заканчивается половым созреванием и началом размножения. Особенности ювенильного периода проявляются в своеобразии питания молодого организма, его поведения и степени зависимости от родителей. С морфологической точки зрения для этого периода характерны интенсивный рост, установление окончательных пропорций между различными частями тела, завершение развития скелета, кожных покровов, смена зубов, завершение развития половых желез и гормональных регуляций. Продолжительность ювенильного периода у позвоночных варьирует от минимальной, равной 13—18 сут у полевок, до максимальной, равной 18—20 годам у белуги, крокодила, альбатроса, слона. У многих представителей позвоночных, особенно у человека, достижение половой зрелости и начало размножения могут быть разделены значительным промежутком времени.
Онтогенез так же делят на:
Пренатальный (дородовый) – организм развивается внутри материнского под зацитой яйцевых оболочек.
Интанатальный (период родов)
Постнатальный (послеродовый).
№2
Морфология половых клеток
Яйцеклетка
Количество желтка сильно варьирует в яйцеклетках животных. Их подразделяют на:
Алецитальные – практически безжелтковые. Характерны для млекопитающих и человека.
Изолецитальные – с небольшим количеством равномерно распределённого желтка.
Телолецитальные:
Умереннотелолецитальные – умеренное содержание желтка. Он сосредоточен на одном полюсе (который называется вегетативным, а другой – анимальным).
Резкотелолецитальные – с большим содержанием желтка.
Центролецитальные – среднее количество желтка, распределённого вокруг ядра, занимающего центральное положение.
Поверхность яйцеклетки покрыта несколькими оболочками:
Первичная – характерна для всех животных. У человека пронизана изнутри микроворсинками яйцеклетки и микроворсинками фолликулярных клеток снаружи. Похожа на блестящий ободок (за что и названа лучистой или блестящей оболочкой).
Вторичная – образуется в яичниках и является продуктами жизнедеятельности фолликулярных клеток.
Третичная – выделяется железами яйцевода. Особенно хорошо развита у птиц и рептилий.
После проникновения сперматозоида в яйцеклетку, в ней начинается интенсивное перемещение цитоплазмы и её расслоение (овоплазматическая сегрегация). Она создаёт определённое пространственную организацию будущего зародыша.
Вегетативный полюс яйцеклетки заполнен цитоплазмой, богатой желтком и митохондриями, а анимальный полюс – безжелтковой цитоплазмой. Первая часть в последствии будет соответствовать энтодерме, а вторая – эктодерме.
Сперматозоид
Сперматозоид – удлиненная подвижная клетка. Главные сперматозоида– ядро, занимающее основной объем головки, и орган движения – жгутик, составляющий хвост. Сперматозоид представляет собой подвижное ядро. Строение сперматозоида обусловлено в первую очередь его функциями. В сперматозоиде очень мало цитоплазмы, но есть несколько вспомогательных структур:
митохондрии, которые обеспечивают его энергией
акросома, органелла, подобная лизосоме и содержащая ферменты, необходимые для проникновения сперматозоида в яйцеклетку
центриоль – начало жгутика, при оплодотворении используется в ходе первого дробления зиготы
Акросома находится впереди ядра в головке, а центриоль и митохондрии – в средней части клетки. Ядро содержит гаплоидный набор хромосом (см. также КЛЕТКА), оно плотное, конденсированное; длинный жгутик сходен по строению со жгутиками простейших и ресничками мерцательного эпителия многоклеточных животных. Сперматозоиды очень живучие клетки и при соответствующих условиях (в матке) они сохраняют жизнеспособность до пяти дней.
№3
Оплодотворение
Оплодотворение — это процесс слияния половых клеток. Образующаяся в результате оплодотворения диплоидная клетка — зигота — представляет собой начальный этап развития нового организма.
Виды оплодотворения:
Внутреннее – яйцеклетка оплодотворяется вне половых путей самки.
Внешнее – яйцеклетка оплодотворяется в половых путях самки. В нём подразделяют:
Оплодотворение со спариванием – многие животные стремятся повысить эффективность внешнего оплодотворения за счёт его синхронизации во времени и пространстве.
Оплодотворение без спаривания.
Оплодотворение происходит в несколько фаз.
Фаза I – сближение. Оно обеспечивается совокупностью, повышающая вероятность сближения гамет (скоординированность к наступлению оплодотворении. у обоих полов, поведение полов, избыточная продукция сперматозоидов, крупные яйцеклетки). Способствуют сближению и взаимодействию половых клеток: гиногамоны – выделяемые яйцеклеткой вещества, активизирующие движение сперматозоидов, вызывают их склеивание при встрече с андрогенами (реакция аглютинизации). Её биологический смысл – предохранение яйцеклетки от избытка сперматозоидов.
Фаза II – в ней происходит акросомная реакция. Наружная мембрана сперматозоида разрывается, высвобождаются протеолитические ферменты и растворяют оболочку яйцеклетки. Плазматические мембраны сливаются, цитоплазмы соединяются. В цитоплазму яйцеклетки переходят ядро и центриоль сперматозоида. Хвостовая часть рассасывается. Затем яйцеклетка активируется, меняется её потенциал и отслаивается её желточная оболочка и образуется оболочка оплодотворения (кортикальная реакция). Активация заканчивается началом синтеза белка.
Фаза III – сингамия. В ней выделяют:
Стадия 2х пронуклеусов – мужское ядро набухает, принимает вид профазного, за это время удваивается ДНК и мужской пронуклеус получает гаплоидный набор редуплецированных хромосом (n2c). Яйцеклетка в момент встречи со сперматозоидом находится в стадии мейоза, заблокированной с помощью специального фактора. После встречи со сперматозоидом, яйцеклетка активируется и блок снимается. Ядро яйцеклетки, закончившее мейоз, превращается в женский пронуклеус, так же приобретая набор хромосом n2c.
Стадия синкариона – слияние ядерного материала и образование зиготы.
Первое митотическое деление зиготы приводит к образованию двух клеток зародыша (бластомеров) с набором хромосом 2n2c в каждом.
Партеногенез
Хотя оплодотворение представляет собой характерный признак полового размножения, дочерний организм иногда развивается из неоплодотворенной яйцеклетки. Это явление называют девственным развитием или партеногенезом. Источником наследственного материала для развития потомка в этом случае обычно служит ДНК яйцеклетки — гиногенез. Реже наблюдается андрогенез — развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской гаметы в случае андрогенеза погибает.
Обязательный партеногенез является измененной формой полового размножения в эволюции некоторых видов животных. У пчел, например, он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворенных яйцеклеток, а мужские (трутни) — партеногенетически. Партеногенез включен в жизненные циклы многих паразитов. Он обеспечивает рост численности особей в условиях, затрудняющих встречу партнеров противоположного пола. Имеются указания на возможность девственного развития у человека. В яичниках девушек, погибших при случайных обстоятельствах, в отсутствие предшествующего осеменения находили зародыши на ранних этапах дробления. Невозможность завершенного партеногенеза у людей в настоящее время доказана и связана с необходимостью наличия обоих геномов, мужского и женского. Наблюдения завершенного эмбриогенеза с партеногенетическим развитием в отношении человека отсутствуют. При партеногенезе, как и при типичном половом размножении, развиваются особи с диплоидными соматическими клетками. Восстановление диплоидного набора хромосом происходит обычно путем слияния ооцита и редукционного тельца во втором делении мейоза.
№4
Эмбриогенез
Дробление – это процесс, связанный с последними митотическими делениями заготы, заканчивающиеся образованием однослойного многоклеточного зародыша – бластулы, а образовавшиеся при этом клетки называют бластомерами. У большинства организмов стенки бластулы расходятся, образуя полость – бластоцель.
Периферийная часть зиготы превращается в сплошной слой клеток – бластодерму.
Дробление бывает:
Полным (гологластическим)
Неполным (мерабластическим)
Полное дробление, в свою очередь, бывает:
Равномерным (характерно для алецетальных и изолецитальных яиц. Заканчивается образованием бластулы)
Неравномерным (характерно для умереннолецитальных и телолецитальных яиц. Завершается образованием амфебластулы).
Неполное дробление бывает:
Дискоидальным (характерно для резколецитальных яиц. Завершается образованием дискобластулы). Бластоцель в виде цели.
Поверхносным (характерно для центролецитальных. В результате образуется перибластула, а бластоцель заполнена желтком).
Особенности дробления:
Отсутствие роста развивающегося организма.
Первое деление зиготы происходит синхронно, а затем длительность клеточных циклов меняется и деление идёт асинхронно.
На стадии дробления начинается дифференцировка клеточных систем. Клеточные ядра попадают в качественно и количественно различные области цитоплазмы и в клетках включается в работу разные наборы генов.
Геном зародыша неактивен (То есть нет транскрипции).
Отсутствие s-фазы (То есть клетки после деления почти сразу преступают к следующему делению).
№5
Образование зародышевых листков
Гаструляция – это процесс образования зародышевых листков. В результате перемещения эмбрионального материала, в начале образуется ранняя гаструла (когда зародыш состоит из эктодермы и энтодермы), а затем образуется средний зародышевый листок – мезодерма.
Способы образования ранней гаструлы:
Иммиграция (выселение) – часть клеток бластодермы уходит в бластоцель, образуя внутренний зародышевый листок.
Инвагинация – участок бластодермы впячивается внутрь, образуя энтодерму, которая выстилает полость первичной кишки.
Эпиболия (обрастание) – характерно для телолецитальных яиц. Образование гаструлы идёт здесь за счёт быстрого деления микромеров, которые обрастают вегетативный полюс яйца. Макромеры с большим содержанием желтка, не способны к перемещению и образуют энтодерму.
Деноминация (расслоение) – клетки бластодермы разделяются на экто- и энтодерму.
Формирование мезодермы:
Телопластический (характерен для первичноротых организмов). На границе между экто- и энтодермой по краям бластопора клетки начинают делиться и дают начало мезодерме.
Энтероцельный (характерен для вторичноротых организмов). Клетки мезодермы закладываются в виде карманов первичной кишки. Полости карманов соответствуют целомической полости. В мезодерме выделяют определённые участки, который дают начало тканям и органам.
Строение инвагинационной гаструлы:
Инвагинация проходит несколько иначе. На границе между анимальным и вегетативным полюсами в области серого серпа клетки сначала сильно вытягиваются внутрь, принимая вид «колбовидных», а затем тянут за собой клетки поверхностного слоя бластулы. Возникают серповидная бороздка и спинная губа бластопора.
Одновременно более мелкие клетки анимального полюса, делящиеся быстрее, начинают перемещаться в сторону вегетативного полюса. В области спинной губы они подворачиваются и впячиваются, а с боков и со стороны, противоположной серповидной бороздке, обрастают более крупные клетки. Затем процесс эпиболии приводит к тому, что образуются боковые и брюшная губы бластопора. Бластопор смыкается в кольцо, внутри которого некоторое время видны крупные светлые клетки вегетативного полюса в виде так называемой желточной пробки. Позднее они полностью погружаются внутрь, а бластопор сужается.
Гаструляция у человека:
Гаструляция у человека происходит в два этапа. Первый этап идет путем деляминации (расщепления), а второй - путем миграции.
В процессе первого этапа образуются два зародышевых листка (экто- и энтодерма), два провизорных органа (амнион и желточный мешок). Кроме того, непосредственно перед началом первого этапа происходит образование такого провизорного органа, как хорион. Формирование хориона - это второй этап в образовании плаценты.
Во время второго этапа образуются еще два зародышевых листка (мезодерма и мезенхима), провизорный орган - аллантоис и идет дальнейшее образование еще одного провизорного органа - плаценты: формируются третичные ворсинки хориона, которые в последующем соединяются с decidua basalis и формируют плаценту. Образуются осевые органы - хорда, нервная трубка, кишечная трубка, мезодерма.
№6
Гисто и органогенез
В самом начале формируются осевые органы.
Самое начало органогенеза называют нейруляцией. Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку. Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры — сомиты.
Нервная система позвоночных, включая человека, отличается устойчивостью основного плана строения на протяжении всей эволюционной истории подтипа. В формировании нервной трубки у всех хордовых много общего. Вначале неспециализированная спинная эктодерма, отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку, представленную нейроэпителиальными клетками цилиндрической формы.
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри — невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитохалазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу и образуют два главных потока. Клетки одного из них—поверхностного—включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении, образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма, занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой. Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов — склеротомы —превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образует миотомы, из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы, которые дают начало внутреннему слою кожи — дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела — целом. Внутренний листок, прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов нервная трубка — хорда — кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляции. Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов. Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка.
Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения; слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретирующие клетки передней и средней долей гипофиза, щитовидной и паращитовидной желез.
Мезодерма помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа.
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется и в итоге в его формировании принимают участие два или три зародышевых листка.
Провизорные органы
В строении и функциях провизорных органов различных амниот много общего. Характеризуя в самом общем виде провизорные органы зародышей высших позвоночных, называемые также зародышевыми оболочками, следует отметить, что все они развиваются из клеточного материала уже сформировавшихся зародышевых листков. Некоторые особенности имеются в развитии зародышевых оболочек плацентарных млекопитающих, о чем будет сказано ниже.
Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
Хорион (сероза) — самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция — дыхательный газообмен; у млекопитающих он выполняет гораздо более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, например гормонов.
Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мезодермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например в спланхноплевре желточного мешка, развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.
У млекопитающих нет запасов желтка и сохранение желточного мешка может быть связано с важными вторичными функциями. Энтодерма желточного мешка служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке.
Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя — сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
№7
Эмбриогенез
Врожденные пороки развития (ВПР) – это стойкие морфологические изменеия органа(ов) или организма, выходящее за пределы вариация их строения и сопровождающееся нарушением их функций.
ВПР бывают:
Наследственные.
Экзогенные.
Мультифакториальные.
Наиболее распространённые ВПР:
Агенезия (полное врождённое отсутствие органа).
Аплазия (врожденное отсутствие органа с сохранением его сосудистой ножки)
Атрезия и стеноза (полное отсутствие или сужение естественного канала или отверстия)
Отсутствие отдельных частей органа обозначают приставкой –олию
Врожденная гипоплазия (недоразвитие органа или системы)
Эктопия (смещение органа в нетипичное место)
Дополнительный орган означают приставкой –поли
Слияние конечностей или их частей приставками –син или –сим.
Неразделение органов или монозиготных близнецов.
Критический период эмбриогенеза – это период эмбриогенеза, отличающийся повышенной чувствительностью зародыша к повреждающему действию факторов внешней среды.
В медицине выделяют клиническую тератологию. Она изучает этиологию, патогенез, клинические проявления, методы диагностики, лечения и профилактики врожденных пороков развития.
Группу факторов, которые приводят к развитию тяжёлых анатомических аномалий, называют тератогенами. Выделяют:
физические тератогены (радиация, ультрафиолетовое излучение и т.д.)
химические тератогены (лекарства: противосудорожные агенты, хемотерапевтические препараты, анальгетики, антибиотики, транквилизаторы (например талидамит), наркотические вещетва и алкоголь).
биологические тератогены (вирусы (например краснуха, герпес, гепатит и т.д.), протозойные инвазии (например токсоплазма), бактерии).
№8
Постнатальное развитие человека
Сразу после рождения наступает период, называемый периодом новорожденности. Основанием для этого выделения служит тот факт, что в это время имеет место вскармливание ребенка молозивом в течение 8-10 дней. Новорожденные в начальном периоде приспособления к условиям внеутробной жизни разделяются по уровню зрелости на доношенных и недоношенных. Внутриутробное развитие доношенных детей длится 39-40 нед., недоношенных - 28-38 нед. При определении зрелости учитывают не только эти сроки, но и массу (вес) тела при рождении.
Доношенными считаются новорожденные с массой тела не менее 2500 г (при длине тела не менее 45 см), а недоношенными - новорожденные, имеющие массу тела меньше 2500 г. Кроме массы и длины, учитывают и другие размеры, например обхват груди в соотношении с длиной тела и обхват головы в соотношении с обхватом груди. Считается, что обхват груди на уровне сосков должен быть больше 0,5 длины тела на 9-10 см, а обхват головы - больше обхвата груди не более чем на 1-2 см.
Грудной период
Следующий период - грудной - продолжается до года. Начало этого периода связано с переходом к питанию "зрелым" молоком. Во время грудного периода наблюдается наибольшая интенсивность роста, по сравнению со всеми остальными периодами внеутробной жизни. Длина тела увеличивается от рождения до года в 1,5 раза, а масса тела утраивается. С 6 мес. начинают прорезываться молочные зубы. В грудном возрасте ярко выражена неравномерность в росте тела. В первом полугодии грудные дети растут быстрее, чем во втором. В каждом месяце первого года жизни появляются новые показатели развития. В первый месяц ребенок начинает улыбаться в ответ на обращение к нему взрослых, в 4 мес. настойчиво пытается встать на ножки (при поддержке), в 6 мес. пытается ползать на четвереньках, в 8 - делает попытки ходить, к году ребенок обычно ходит.
Период раннего детства
Период раннего детства длится от 1 года до 4 лет. В конце второго года жизни заканчивается прорезывание зубов. После 2 лет абсолютные и относительные величины годичных приростов размеров тела быстро уменьшаются.
Период первого детства
С 4 лет начинается период первого детства, который заканчивается в 7 лет. Начиная с 6 лет появляются первые постоянные зубы: первый моляр (большой коренной зуб) и медиальный резец на нижней челюсти.
Возраст от 1 года до 7 лет называют также периодом нейтрального детства, поскольку мальчики и девочки почти не отличаются друг от друга размерами и формой тела.
Период второго детства
Период второго детства длится у мальчиков с 8 до 12 лет, у девочек - с 8 до 11 лет. В этот период выявляются половые различия в размерах и форме тела, а также начинается усиленный рост тела в длину. Темпы роста у девочек выше, чем у мальчиков, так как половое созревание у девочек начинается в среднем на два года раньше. Усиление секреции половых гормонов (особенно у девочек) обусловливает развитие вторичных половых признаков. Последовательность появления вторичных половых признаков довольно постоянна. У девочек вначале формируются молочные железы, затем появляются волосы на лобке, потом - в подмышечных впадинах. Матка и влагалище развиваются одновременно с формированием молочных желез. В гораздо меньшей степени процесс полового созревания выражен у мальчиков. Лишь к концу этого периода у них начинается ускоренный рост яичек, мошонки, а затем - полового члена.
Подростковый период
Следующий период - подростковый - называется также периодом полового созревания, или пубертатным периодом. Он продолжается у мальчиков с 13 до 16 лет, у девочек - с 12 до 15 лет. В это время наблюдается дальнейшее увеличение скоростей роста - пубертатный скачок, который касается всех размеров тела. Наибольшие прибавки в длине тела у девочек имеют место между 11 и 12 годами, по массе тела - между 12 и 13 годами. У мальчиков прибавка в длине наблюдается между 13 и 14 годами, а прибавка в массе тела - между 14 и 15 годами. Особенно велика скорость роста длины тела у мальчиков, в результате чего в 13,5-14 лет они обгоняют девочек по длине тела. В связи с повышением активности гипоталамо-гипофизарной системы формируются вторичные половые признаки. У девочек продолжается развитие молочных желез, наблюдается рост волос на лобке и в подмышечных впадинах. Наиболее четким показателем полового созревания женского организма является первая менструация.
В подростковый период происходит интенсивное половое созревание мальчиков. К 13 годам у них происходит изменение (мутация) голоса и появляются волосы на лобке, а в 14 лет появляются волосы в подмышечных впадинах. В 14-15 лет у мальчиков появляются первые поллюции (непроизвольные извержения спермы).
У мальчиков, по сравнению с девочками, более продолжителен пубертатный период и сильнее выражен пубертатный скачок роста.
Юношеский возраст
Юношеский возраст продолжается у юношей от 18 до 21 года, а у девушек - от 17 до 20 лет. В этот период в основном заканчиваются процесс роста и формирование организма и все основные размерные признаки тела достигают дефинитивной (окончательной) величины.
В юношеском возрасте завершается формирование половой системы, созревание репродуктивной функции. Окончательно устанавливаются овуляторные циклы у женщины, ритмичность секреции тестостерона и выработка зрелой спермы у мужчины.
Зрелый, пожилой, старческий возраст
В зрелом возрасте форма и строение тела изменяются мало. Между 30 и 50 годами длина тела остается постоянной, а потом начинает уменьшаться. В пожилом и старческом возрасте происходят постепенные инволютивные изменения организма.
№9
Старение
Старость представляет собой стадию индивидуального развития, по достижении которой в организме наблюдаются закономерные изменения в физическом состоянии, внешнем виде, эмоциональной сфере.
Старческие изменения становятся очевидными и нарастают в пострепродуктивном периоде онтогенеза. Однако начало угасания репродуктивной функции или даже ее полная утрата не могут служить нижней границей старости. Действительно, менопауза у женщин, заключающаяся в прекращении выхода зрелых яйцеклеток из яичника и соответственно прекращении месячных кровотечений, определяет окончание репродуктивного периода жизни. Вместе с тем к моменту достижения менопаузы большинство функций и внешних признаков далеко не достигают состояния, характерного для старых людей. С другой стороны, многие изменения, которые мы связываем со старостью, начинаются до снижения репродуктивной функции. Это относится как к физическим признакам (поседение волос, развитие дальнозоркости), так и к функциям различных органов. К примеру, у мужчин снижение выделения мужских половых гормонов половыми железами и повышение выделения гонадотропных гормонов гипофизом, что характерно для старого организма, начинается примерно от 25 лет.
Различают хронологический и биологический (физиологический) возраст. Согласно современной классификации, основанной на оценке многих средних показателей состояния организма, людей, хронологический возраст которых достиг 60—74 лет, называют пожилыми, 75—89 лет —старыми, свыше 90 лет —долгожителями. Точное определение биологического возраста затруднено тем, что отдельные признаки старости проявляются в разном хронологическом возрасте и характеризуются различной скоростью нарастания. Кроме того, возрастные изменения даже одного признака подвержены значительным половым и индивидуальным колебаниям.
Рассмотрим такой признак, как упругость (эластичность) кожи. Один и тот же биологический возраст достигается в этом случае женщиной примерно в 30 лет, а мужчиной — в 80. Именно поэтому прежде всего женщинам необходим грамотный и постоянный уход за кожей. С целью определения биологического возраста, что необходимо для суждения о скорости старения, используют батареи тестов, проводя совокупную оценку одновременно многих признаков, закономерно изменяющихся в процессе жизни.
Основу таких батарей составляют сложные функциональные показатели, состояние которых зависит от согласованной деятельности нескольких систем организма. Простые тесты обычно бывают менее информативными. Например, скорость распространения нервного импульса, которая зависит от состояния нервного волокна, снижается в возрастном интервале 20—90 лет на 10%, тогда как жизненная емкость легких, определяемая координированной работой дыхательной, нервной и мышечной систем —на 50%.
Состояние старости достигается благодаря изменениям, составляющим содержание процесса старения. Этот процесс захватывает все уровни структурной организации особи — молекулярный, субклеточный, клеточный, тканевой, органный. Суммарный результат многочисленных частных проявлений старения на уровне целостного организма заключается в нарастающем с возрастом снижении жизнеспособности особи, уменьшении эффективности приспособительных, гомеостатических механизмов. Показано, например, что молодые крысы после погружения в ледяную воду на 3 мин восстанавливают температуру тела примерно за 1 ч. Животным среднего возраста на это требуется 1,5 ч, а старым — около 2 ч.
В целом старение приводит к прогрессивному повышению вероятности смерти. Таким образом, биологический смысл старения заключается в том, что оно делает неизбежной смерть организма. Последняя же представляет собой универсальный способ ограничить участие многоклеточного организма в размножении. Без смерти не было бы смены поколений — одного из главных условий эволюционного процесса.
Возрастные изменения в процессе старения не во всех случаях заключаются в снижении приспособляемости организма. У человека и высших позвоночных в процессе жизни приобретается опыт, вырабатывается умение избегать потенциально опасных ситуаций. Интересна в этом плане и система иммунитета. Хотя ее эффективность после достижения организмом состояния зрелости в целом снижается, благодаря «иммунологической памяти» по отношению к некоторым инфекциям старые особи могут оказаться более защищенными, чем молодые.
Изменения в органах и системах организма в процессе старения
Как правило, после 40—50 лет у человека возникают стойкие внешние проявления старения, в частности кожных покровов. Появляются морщины, образующиеся из-за потери подкожной жировой ткани, пигментные пятна, бородавки. Кожа становится сухой и шершавой в связи с уменьшением количества потовых желез, теряется ее эластичность, она становится дряблой.
Признаки старения сердечно-сосудистой системы становятся заметными обычно в возрасте после 40 лет. Закономерные изменения наблюдаются в стенках сосудов: в них откладываются липиды, прежде всего холестерин, что наряду с другими структурными превращениями снижает эластичность и искажает ответы на различные стимулы, регулирующие кровообращение. Типичным является разрастание в стенках сосудов и сердца соединительной ткани, замещающей рабочую мышечную ткань. В результате снижается эффективность работы сердца. В итоге нарушается кровоснабжение тканей и органов. Так, кровоток по сосудам головного мозга 75-летнего человека по сравнению с 30-летним уменьшен на 20%.
В основе функциональных расстройств дыхательной системы лежит разрушение межальвеолярных перегородок, что сокращает дыхательную поверхность, разрастание в легких соединительной ткани, снижает эффективность аэрогематического обмена кислорода. В итоге с возрастом падает жизненная емкость легких, которая к 75 годам достигает всего 56% от уровня в возрасте 30 лет.
Легко заметным изменением в системе пищеварения является потеря зубов. Падает эффективность функционирования пищеварительных желез, нарушения двигательной (моторной) функции кишечника нередко приводят к привычным запорам.
В процессе старения страдает функция мочевыделительной системы, снижается интенсивность фильтрации в почечных клубочках (на 31% в 75-летнем возрасте по сравнению с 30-летним), так же как и обратное всасывание веществ из фильтрата в почечных канальцах. Ухудшение функции мочевыделения объясняется гибелью с возрастом значительного количества нефронов (до 44% от уровня 30-летнего возраста), представляющих собой структурно-функциональные единицы почек.
Специального внимания заслуживают изменения в процессе старения со стороны мышечной системы и скелета. Снижается сила сокращений поперечно-полосатой мускулатуры, быстрее развивается утомление, наблюдается атрофия мышц. Характерная для стареющих людей перестройка костей заключается в разрежении их вещества (старческий остеопороз), что приводит к снижению прочности.
В процессе старения организма существенные изменения происходят в репродуктивной системе. При этом они затрагивают обе основные функции главных органов названной системы — половых желез: выработку гамет и образование половых гормонов. У женщин овогенез прекращается по достижении ими менопаузы. Образование функционально полноценных сперматозоидов в мужском организме возможно, по-видимому, даже в преклонном возрасте.
Изменение гормонального профиля людей в связи с угасанием репродуктивной функции носит сложный характер. Распространенным является мнение о прогрессивном снижении с возрастом концентрации у мужчин тестостерона, а у женщин эстрадиола и прогестерона — главных мужского и женских половых гормонов. Напомним, что оба типа гормонов образуются организмами обоих полов, только в разном количестве. Указанные сдвиги сопровождаются повышением секреции эстрадиола и прогестерона у мужчин и тестостерона у женщин. Вместе с тем содержание фолликулостимулирующего гормона у 80—90-летних женщин выше в 14 раз, а лютеинизирующего гормона — в 5 раз, чем у 20—30-летних. Резко нарушено у старых людей соотношение названных гормонов гипофиза, что является важной причиной нарушения репродуктивной функции в целом. Картина усложняется также тем, что в процессе старения изменяется ответ ткани на половые гормоны в связи с сокращением количества клеточных рецепторов к ним.
Изменения в процессе старения функций эндокринной системы носят объективно сложный характер. В качестве примера рассмотрим изменения в стареющем организме функции щитовидной железы. Обнаружено, что к старости падает содержание в крови трииодтиронина и тироксина, в связи с изменением белков плазмы крови ухудшается перенос гормонов к тканям, в клетках уменьшается количество рецепторов, узнающих гормоны, а чувствительность рецепторов повышается. Вместе с тем в крови сохраняется достаточно высокая концентрация тиреотропного гормона гипофиза; чувствительность к нему клеток щитовидной железы возрастает. Из приведенной картины видно, что в отдельных звеньях цепи регуляции жизненно важных функций гормонами щитовидной железы возрастные изменения не одинаковы по масштабу, а иногда и разнонаправлены. В таком случае важен общий результат, степень выраженности которого подвержена индивидуальным колебаниям.
Наряду с эндокринной системой нервной системе принадлежит важнейшая роль в регуляции, координации и интеграции разнообразных проявлений жизнедеятельности. Изменения нервной системы в процессе старения включают нарастающую гибель нейронов, масштабы которой колеблются от отдела к отделу и составляют от 15—20 до 70—75%. С другой стороны, сохранившиеся клетки увеличивают свои размеры, образуют дополнительные разветвления окончаний отростков в тканях-мишенях, что имеет приспособительное, заместительное значение. Общий итог, однако, заключается в том, что после 40—50 лет в тканях образуется недостаток нервных влияний, что приводит к нарушению регуляций функций. Это справедливо, к примеру, в отношении иннервации сердца и сосудов, а также поперечно-полосатой скелетной мускулатуры симпатической нервной системой.
Функциональные нарушения в процессе старения в центральной нервной системе зависят как от уменьшения количества нервных клеток, так и от снижения синтеза медиаторов, ослабления связей между отдельньми мозговыми структурами, уменьшения скорости проведения нервных импульсов по волокнам и через синапсы. В стареющем организме с большим трудом вырабатываются условные рефлексы, приобретаются новые навыки. Изменения ряда функций носят более сложный характер. Так, интеллектуальные способности людей в возрасте 55—70 лет сохраняются такими же, какими они были в 20—30-летнем возрасте, а в возрасте 55—60 лет наблюдается второй пик творческой деятельности.
Одна из черт процесса старения заключается в снижении надежности механизмов регуляции, направленных на поддержание постоянства жизненно важных параметров внутренней среды организма — гомеостаза. Причину этого видят в функциональных изменениях стареющего гипоталамуса, с которым связывают действия своеобразных биологических часов старения всего организма. К гипоталамической области головного мозга человека относятся 32 пары ядер, участвующих в регуляции важнейших вегетативных функций. В целом процесс старения гипоталамических структур характеризуется неравномерностью и разнонаправленностью, что типично для других областей нервной системы.
В старости наблюдается снижение функций практически всех органов чувств. Уменьшается способность глаза к аккомодации, так как слабеют глазодвигательные мышцы и изменяется вещество хрусталика. Это ведет к старческой дальнозоркости. У старых людей хрусталик нередко теряет прозрачность — развивается катаракта. Падает острота зрения. Чувствительность органа слуха снижается, причем особенно к высокочастотным звуковым колебаниям. Старый человек хуже различает запах и вкус. К 75 годам люди сохраняют примерно 36% от количества вкусовых луковиц, которое имеется в возрасте 30 лет. У них нарушается чувство равновесия.
С
возрастом заметно изменяются функции
иммунной
системы.
За гуморальный иммунитет ответственны
В-лимфоциты, вырабатывающие антитела
к носителям чужеродной биологической
информации — антигенам. Т-лимфоциты
ответственны за клеточный иммунитет,
например за отторжение трансплантата,
хотя могут участвовать в реакциях
гуморального иммунитета. К старости
наблюдается ослабление реакций как
гуморального, так и клеточного иммунитета.
Становление функции иммунной системы
во многом связано с активностью тимуса,
которая прекращается в связи с инволюцией
железы по достижении половозрелого
возраста (рис. 8.30). Нарушение естественного
развития иммунной системы в эксперименте
путем удаления вилочковой железы
(например, у мышей) сокращает
продолжительность жизни животных.
Напротив, пересадка тимуса и костного
мозга от молодой мыши к старой приводит
к омоложению иммунной системы 19-месячной
мыши до уровня 4-месячной. Некоторые
такие животные жили на
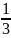 дольше своего обычного срока.
дольше своего обычного срока.
В стареющем организме клетки иммунной системы ошибочно вырабатывают антитела против собственных клеток и белков. Таким образом, старение сопровождается нарастанием аутоиммунных реакций.
Изменения в процессе старения органов кроветворной системы заключаются в снижении продукции эритроцитов. С возрастом способность костного мозга к восстановлению массы эритроцитов падает почти в 6 раз. Восполнение утраченных вследствие потери 30—40% общего объема крови эритроцитов происходит у 2-месячных крыс по 4,8%, а у 25-месячных —по 0,8% в 1 сут.
Из изложенного в данном разделе следует, что процесс старения распространяется практически на все структуры и функции организма. При этом некоторые показатели (упругость кожи, поседение волос) изменяются с возрастом довольно монотонно. Другие (артериальное давление, жизненная емкость легких) изменения носят пороговый характер, начиная регистрироваться примерно в возрасте 35—45 лет.
Клиническая смерть — это последний этап умирания. По определению академика Неговского — «клиническая смерть уже не является жизнью, но ещё не является смертью. Это возникновение нового качества — перерыв непрерывности. В биологическом смысле это состояние напоминает анабиоз, хотя и не идентично этому понятию». Клиническая смерть является обратимым состоянием и сам по себе факт прекращения дыхания или кровообращения не является доказательством наступления смерти.
К признакам клинической смерти можно отнести:
Отсутствие дыхания.
Отсутствие сердцебиения.
Генерализованная бледность или генерализованный цианоз.
Отсутствие реакции зрачков на свет.
Биологическая смерть (или истинная смерть) представляет собой необратимое прекращение физиологических процессов в клетках и тканях. Под необратимым прекращением обычно понимается «необратимое в рамках современных медицинских технологий» прекращение процессов. Со временем меняются возможности медицины по реанимации умерших пациентов, вследствие чего, граница смерти отодвигается в будущее. С точки зрения учёных – сторонников крионики и наномедицины, большинство умирающих сейчас людей могут быть в будущем оживлены, если сейчас сохранить структуру их мозга.
К ранним признакам биологической смерти относятся:
Отсутствие реакции глаза на раздражение (надавливание)
Помутнение роговицы, образование треугольников высыхания (пятен Лярше).
Появление симптома «кошачьего глаза»: при боковом сдавлении глазного яблока зрачок трансформируется в вертикальную веретенообразную щель.
№10
Проблемы регенерации и трансплантации
Регенерация (от лат. regeneratio — возрождение) — процесс восстановления организмом утраченных или поврежденных структур. Регенерация поддерживает строение и функции организма, его целостность.
Различают два вида регенерации: физиологическую и репаративную.
Восстановление органов, тканей, клеток или внутриклеточных структур после разрушения их в процессе жизнедеятельности организма называют физиологической регенерацией. Восстановление структур после травмы или действия других повреждающих факторов называют репаративной регенерацией.
Физиологическая регенерация представляет собой процесс обновления функционирующих структур организма. Благодаря физиологической регенерации поддерживается структурный гомеостаз и обеспечивается возможность постоянного выполнения органами их функций. С общебиологической точки зрения, физиологическая регенерация, как и обмен веществ, является проявлением такого важнейшего свойства жизни, как самообновление.
На внутриклеточном уровне – восстановление субклеточных структур в клетках всех органов и тканей (что очень важно для нервной ткани). На клеточном уровне обновление эпидермиса кожи и так далее.
Обновление производных эпидермиса (волосы и ногти) происходит за счёт пролиферативной регенерации – то есть восполнение клеток за счёт их деления.
Физиологическая регенерация присуща всем организмам.
Репаративная регенерация – регенерация после повреждения ткани или органа.
Она очень разнообразна по факторам, вызывающим повреждения, по объемам повреждения, по способам восстановления. Механическая травма, например оперативное вмешательство, действие ядовитых веществ, ожоги, обморожения, лучевые воздействия, голодание, другие болезнетворные агенты,— все это повреждающие факторы. Наиболее широко изучена регенерация после механической травмы.
Существует несколько разновидностей или способов репаративной регенерации. К ним относят эпиморфоз, морфаллаксис, заживление эпителиальных ран, регенерационную гипертрофию, компенсаторную гипертрофию.
Виды репаративной регенерации:
Эпителизация – заживление ран с нарушенным эпителиальным покровом.
Эпиморфоз – отрастание нового органа от ампутированной поверхности. В ней подразделяют:
Регрессивная фаза начинается с заживления раны, во время которого происходят следующие основные события: остановка кровотечения, сокращение мягких тканей культи конечности, образование над раневой поверхностью сгустка фибрина и миграция эпидермиса, покрывающего ампутационную поверхность. Затем начинается разрушение остеоцитов на дистальном конце кости и других клеток. Одновременно в разрушенные мягкие ткани проникают клетки, участвующие в воспалительном процессе, наблюдается фагоцитоз и местный отек. Затем вместо образования плотного сплетения волокон соединительной ткани, как это происходит при заживлении ран у млекопитающих, в области под раневым эпидермисом утрачиваются дифференцированные ткани. Характерна остеокластическая эрозия кости, что является гистологическим признаком дедифференцировки. Раневой эпидермис, уже пронизанный регенерирующими нервными волокнами, начинает быстро утолщаться. Промежутки между тканями все более заполняются мезенхимоподобными клетками. Скопление мезенхимных клеток под раневым эпидермисом является главным показателем формирования регенерационной бластемы. Клетки бластемы выглядят одинаково, но именно в этот момент закладываются основные черты регенерирующей конечности.
Затем начинается прогрессивная фаза, для которой наиболее характерны процессы роста и морфогенеза. Длина и масса регенерационной бластемы быстро увеличиваются. Рост бластемы происходит на фоне идущего полным ходом формирования черт конечности, т.е. ее морфогенеза. Когда форма конечности в общих чертах уже сложилась, регенерат все еще меньше нормальной конечности. Чем крупнее животное, тем больше эта разница в размерах. Для завершения морфогенеза требуется время, по истечении которого регенерат достигает размеров нормальной конечности.
При эпиморфозе не всегда образуется точная копия удалённой структуры – такая регенерация называется атипичной. В ней разделяют:
Гиперморфоз – регенерация с частичным замещением ампутированной конечности.
Гетероморфоз – появление иной структуры на местеутраченой (появление конечности вместо глаз у членестоногих).
Избыточная регенерация – образование дополнительных структур (появление у планарии второй головы)
Морфолаксис – регенерация путём перестройки регенерирующего участка (Примером служит регенерация гидры из кольца, вырезанного из середины ее тела, или восстановление планарии из одной десятой или двадцатой ее части)
Регенерационная гипертрофия – увеличение размеров остатка органа без восстановления исходной формы (печень у млекопитающих).
Компенсаторная гипертрофия – измененя в одном из органов при нарушении в другом, относящимся к той же системе (например гипертрофия одно из почек при удалении другой).
Трансплантация
Трансплантация — в медицине пересадка какого-либо органа или ткани, например почки, сердца, печени, лёгкого, костного мозга, стволовых гемопоэтических клеток.
Различают следующие виды трансплантации:
Аутотрансплантация, или аутологичная трансплантация — реципиент трансплантата является его донором для самого себя. Например, аутотрансплантация кожи с неповреждённых участков на обожжённые широко применяется при тяжёлых ожогах. Аутотрансплантация костного мозга или гемопоэтических стволовых клеток после высокодозной противоопухолевой химиотерапии широко применяется при лейкозах, лимфомах и химиочувствительных злокачественных опухолях.
Гомотрансплантация, или гомологичная трансплантация — донором трансплантата является 100% генетически и иммунологически идентичный реципиенту однояйцевый близнец реципиента. Вероятно, в будущем, когда научатся клонировать отдельные ткани и выращивать в лабораторных условиях целые органы человеческого тела с заданными иммунологическими характеристиками, все 100% трансплантаций органов будут гомологичными.
Аллотрансплантация, или гетерологичная трансплантация — донором трансплантата является генетически и иммунологически другой человеческий организм.
Ксенотрансплантация, или межвидовая трансплантация — трансплантация органов от животного другого биологического вида, чем человек.
№11
Современный дарвинизм
Популяция - элементарная единица эволюции. Современная эволюционная биология в качестве элементарной единицы эволюции рассматривает популяцию. Популяцией называют сообщество особей одного вида, занимающих определенную территорию и связанных друг с другом родственными узами.
Ч.Дарвин считал наследственную изменчивость особей, борьбу за существование и естественный отбор главными движущими силами (факторами) процесса эволюции. В настоящее время исследования в области эволюционной биологии подтвердили справедливость этого утверждения и выявили ряд других факторов, которые играют важную роль в процессе эволюции.
Вы знаете, что эволюция – это наследственное изменение свойств и признаков живых организмов в ряду поколений. Это означает, что отдельные особи не могут эволюционировать. Каждая особь развивается на основе генотипа, унаследованного от родителей. Генотип определяет особенности ее развития, ее взаимоотношения с внешней средой, в том числе и возможность адаптивных модификаций в ответ на изменение внешних условий. Но как бы ни менялась особь, ее генотип остается неизменным. Таким образом, элементарной единицей эволюции является не особь, а популяция. Совокупность генотипов всех особей в популяции называют генофондом. В ходе эволюции меняется набор генотипов в генофонде популяций. Одни генотипы распространяются, а другие становятся редкими и постепенно исчезают. Эффективность размножения и распространения в популяции каждого конкретного генотипа зависит от того, насколько фенотип особи, созданной на его основе, соответствует тем условиям, которые существуют в то время и в том месте, где живет эта особь. Если особь доживает до размножения и производит потомков, то она передает им полностью или частично тот генотип, который позволила ей это сделать, и в следующем поколении носителей этого «удачного» генотипа становится больше. Мы можем сказать, что ее генотип распространяется в генофонде популяции. Если особь погибает до размножения или не оставляет потомков, то вместе с ее смертью пресекается распространение и ее генотипа. В следующем поколении уже будет относительно меньше носителей этого генотипа, не подходящего к тем условиям, в которых живет популяция. Однако условия жизни изменяются постоянно и непредсказуемо. Изменяются климат, ландшафт, характеристики других видов (хищников, жертв, паразитов, конкурентов), с которыми взаимодействуют особи данной популяции, изменяется численность и плотность самой популяции. Соответственно изменяется и полезность фенотипов. То, что было полезным в предыдущем поколении, может оказаться вредным в последующем и наоборот. Условия жизни меняются не только во времени, но и в пространстве. Каждый вид занимает определенную территорию, которая называется ареалом. Иногда ареал вида ограничивается небольшим островком, а иногда охватывает целые континенты. Условия жизни особей из разных частей ареала широко распространенных видов сильно различаются. Генотипы, которые полезны, например, на севере ареала, могут оказаться вредными на юге. То, что хорошо в долине, плохо в горах, и наоборот. В каждой популяции отбираются те генотипы, которые обеспечивают наилучшую адаптацию их носителей к местным условиям. Частота генотипов, которые обеспечивают выживание в долинах, увеличивается в долинных популяциях и уменьшается в горных. Формируются генетические различия между популяциями. Однако между популяциями одного вида постоянно происходит обмен особями и, следовательно, генетическими программами. Миграции животных, перенос пыльцы растений, спор грибов и микроорганизмов ведет к постоянному перемешиванию генетического состава популяций, к уменьшению различий между популяциями и к увеличению разнообразия внутри популяций. Не остаются постоянными и сами генотипы. Отдельные их элементы– гены – также меняются со временем. Разные мутации в разных генах возникают у разных особей, меняя при этом генотипы потомков этих особей. Все организмы с половым размножением передают потомкам свои генотипы не полностью, а частично - каждый потомок получает половину генов от матери и половину от отца и оказывается носителем уникальной комбинацией аллелей, полученных от родителей. Каждая особь имеет уникальный генотип, который лишь частично передается (или не передается вовсе) ее потомкам. Таким образом, мы можем описать процесс эволюции как изменение частот разных аллелей в популяциях. Естественно, это будет неполное и сильно упрощенное описание эволюции, но такой подход позволит нам яснее представить, какие факторы и в какой степени определяют эволюционный процесс.
Мутационный процесс
Изменения наследственного материала половых клеток в виде генных, хромосомных и геномных мутаций происходят постоянно. Особое место принадлежит генным мутациям. Они приводят к возникновению серий аллелей и, таким образом, к разнообразию содержания биологической информации.
Вклад мутационного процесса в видообразование носит двоякий характер. Изменяя частоту одного аллеля по отношению к другому, он оказывает на генофонд популяции прямое действие. Еще большее значение имеет формирование за счет мутантных аллелей резерва наследственной изменчивости. Это создает условия для варьирования аллельного состава генотипов организмов в последовательных поколениях путем комбинативной изменчивости. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет исходный элементарный эволюционный материал. В процессе видообразования он используется как основа действия других элементарных эволюционных факторов.
Хотя отдельная мутация — событие редкое, общее число мутаций значительно. Допустим, что некая мутация возникает с частотой 1 на 100 000 гамет, количество локусов в геноме составляет 10 000, численность особей в одном поколении равна 10 000, а каждая особь производит 1000 гамет. При таких условиях по всем локусам за поколение в генофонде вида произойдет 106 мутаций. За среднее время существования вида, равное нескольким десяткам тысяч поколений, количество мутаций составит 1010. Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие. В силу рецессивности мутантные аллели обычно присутствуют в генофондах "популяций в гетерозиготных по соответствующему локусу генотипах.
Благодаря этому достигается тройственный положительный результат:
Исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном.
Сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем.
Накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса).
Таким образом создается резерв наследственной изменчивости популяции.
Доля полезных мутаций мала, однако их абсолютное количество в пересчете на поколение или период существования вида может быть большим. Допустим, что одна полезная мутация приходится на 1 млн. вредных. Тогда в рассматриваемом выше примере среди 106 мутаций за одно поколение 104 будет полезной. За время существования вида его генофонд обогатится 104 полезными мутациями.
Мутационный процесс, выполняя роль элементарного эволюционного фактора, происходит постоянно на протяжении всего периода существования жизни, а отдельные мутации возникают многократно у разных организмов. Генофонды популяций испытывают непрерывное давление мутационного процесса. Это обеспечивает накопление мутаций, несмотря на высокую вероятность потери в ряду поколений единичной мутации.
Популяционные волны
Популяционными волнами или волнами жизни (С.С. Четвериков) называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причины колебаний часто имеют экологическую природу. Так, размеры популяций «жертвы» (зайца) растут при снижении давления на них со стороны популяций «хищника» (рыси, лисицы, волка). Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что, в свою очередь, интенсифицирует истребление жертвы.
Вспышки численности организмов некоторых видов, наблюдавшиеся в ряде регионов мира, были обусловлены деятельностью человека. В XIX—XX вв. это относится к популяциям кроликов в Австралии, домовых воробьев в Северной Америке, канадской элодеи в Евразии. В настоящее время существенно возросли размеры популяций домовой мухи, находящей прекрасную кормовую базу в виде разлагающихся пищевых отбросов вблизи поселений человека. Напротив, численность 12 популяций домовых воробьев в городах падает вследствие прекращения широкого использования лошадей. Масштабы колебаний численности организмов разных видов варьируют. Для одной из зауральских популяций майских жуков отмечены изменения количества особей в 106 раз.
Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов. Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. В условиях возросшей численности интенсифицируются межпопуляционные миграции особей, что также способствует перераспределению аллелей. Рост количества организмов обычно сопровождается расширением занимаемой территории.
На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида и оказываются в необычных условиях существования. В таком случае они испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование.
При спаде численности наблюдается распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом потеряны. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает. На спаде волны жизни часть популяций, как правило, небольших по размерам, остается за пределами обычного ареала вида. Чаще они, испытывая действие необычных условий жизни, вымирают. Реже, при благоприятном генетическом составе, такие популяции переживают период спада численности. Будучи изолированными от основной массы вида, существуя в необычной среде, они нередко являются родоначальниками новых видов.
Популяционные волны — это эффективный фактор преодоления генетической инертности природных популяций. Вместе с тем их действие на генофонды не является направленным. В силу этого они, так же как и мутационный процесс, подготавливают эволюционный материал к действию других элементарных эволюционных факторов.
Изоляция
Ограничение свободы скрещиваний (панмиксии) организмов называют изоляцией.
Виды изоляции:
Биологическая изоляция возникает вследствие внутривидовых различий организмов и имеет несколько форм.
К экологической изоляции приводят особенности окраски покровов или состава пищи, размножение в разные сезоны, у паразитов — использование в качестве хозяина организмов разных видов. Так, в Молдове есть две несмешивающиеся популяции мышей — желтогорлая лесная мышь и степная. Фактором разделения их является состав пищи. Разобщение популяций способствовало выявлению и усилению особенностей фенотипа степных мышей. Они мельче и имеют иную форму черепа. В описанном примере экологическая изоляция дополняется территориальной. Сезонные расы, выделяемые по месту и времени икрометания, описаны у лососевых, осетровых, карповых рыб.
Длительная экологическая изоляция способствует дивергенции популяций вплоть до образования новых видов. Так, предполагают, что человеческая и свиная аскариды, морфологически очень близкие, произошли от общего предка. Их расхождению, согласно одной из гипотез, способствовал запрет на употребление человеком в пищу свиного мяса, который по религиозным соображениям распространялся длительное время на значительные массы людей.
Этологическая (поведенческая) изоляция существует благодаря особенностям ритуала ухаживания, окраски, запахов, «пения» самок и самцов из разных популяций. Так, подвиды щеглов — седоголовый Carduelis carduelis carduelis и черноголовый С. с. brevirostis — имеют выраженные отметины на голове. Серые вороны Corvus corone cornix из крымской и североукраинской популяций, внешне неразличимые, отличаются карканьем.
При физической (механической) изоляции препятствием к скрещиванию являются различия в структуре органов размножения или просто разница в размерах тела. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей.
Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания.
Генетическая (репродуктивная) изоляция создает более жесткие, иногда непреодолимые барьеры скрещиваниям. Она заключается в несовместимости гамет, гибели зигот непосредственно после оплодотворения, стерильности или малой жизнеспособности гибридов.
Естественный отбор и его формы
Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции.
Виды естественного отбора:
В зависимости от результата различают стабилизирующую, движущую и дизруптивную формы естественного отбора.
Стабилизирующий отбор сохраняет в популяции средний вариант фенотипа или признака. Он устраняет из репродуктивного процесса фенотипы, уклоняющиеся от сложившейся адаптивной «нормы», приводит к преимущественному размножению типичных организмов. Так, сотрудник одного из университетов США подобрал после снегопада и сильного ветра 136 оглушенных воробьев Passer domesticus. Из них 72 выживших воробья имели крылья средней длины, тогда как 64 погибшие птицы были либо длиннокрылыми, либо короткокрылыми. Стабилизирующая форма соответствует консервативной роли естественного отбора. При относительном постоянстве условий среды благодаря этой форме сохраняются результаты предшествующих этапов эволюции.
Движущий (направленный) отбор обусловливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде. После того как новое значение признака придет в оптимальное соответствие условиям среды, движущая форма отбора сменяется стабилизирующей. Примером такого отбора является замещение в популяции гавани Плимут (Англия) крабов Carcinus maenas с широким головогрудным щитком животными с узким щитком в связи с увеличением количества ила. Направленный отбор составляет основу искусственного отбора. Так, в одном эксперименте на протяжении ряда поколений из популяции шестинедельных мышей отбирали для скрещивания наиболее тяжелых и наиболее легких животных. Избирательное воспроизведение по признаку массы тела привело к образованию двух самостоятельных популяций, соответственно с возрастающей и убывающей массой тела. По окончании опыта, занявшего 11 поколений, ни одна из этих популяций не вернулась к первоначальной массе.
Дизруптивный (разрывающий) отбор сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков. Так, в зависимости от преобладающего цвета почвы улитки Cepaea nemoralis имеют раковины коричневой, желтой, розовой окраски. Дизруптивная форма отбора «разрывает» популяцию по определенному признаку на несколько групп. Она поддерживает в популяции состояние генетического полиморфизма.
№12
Биологический вид – качественный этап эволюции
Видом называют совокупность особей, сходных по основным морфологическим и функциональным признакам, кариотипу, поведенческим реакциям, имеющих общее происхождение, заселяющих определенную территорию (ареал), в природных условиях скрещивающихся исключительно между собой и при этом производящих плодовитое потомство.
Видовая принадлежность особи определяется по соответствию ее перечисленным критериям: морфологическому, физиолого-биохимическому, цитогенетическому, этологическому, экологическому и др. Наиболее важные признаки вида — его генетическая (репродуктивная) изоляция, заключающаяся в нескрещиваемости особей данного вида с представителями других видов, а также генетическая устойчивость в природных условиях, приводящая к независимости эволюционной судьбы.
Со времен К. Линнея вид является основной единицей систематики. Особое положение вида среди других систематических единиц (таксонов) обусловливается тем, что это та группировка, в которой отдельные особи существуют реально. В составе вида в природных условиях особь рождается, достигает половой зрелости и выполняет свою главную биологическую функцию: участвуя в репродукции, обеспечивает продолжение рода. В отличие от вида таксоны надвидового ранга, такие, как род, отряд, семейство, класс, тип, не являются ареной реальной жизни организмов. Выделение их в естественной системе органического мира отражает результаты предшествующих этапов исторического развития живой природы. Распределение организмов по надвидовым таксонам указывает на степень их филогенетического родства.
Важнейшим фактором объединения организмов в виды служит половой процесс. Представители одного вида, скрещиваясь друг с другом, обмениваются наследственным материалом. Это ведет к перекомбинации в каждом поколении генов (аллелей), составляющих генотипы отдельных особей. В результате достигаются нивелировка различий между организмами внутри вида и длительное сохранение основных морфологических, физиологических и прочих признаков, отличающих один вид от другого. Благодаря половому процессу происходит также объединение генов (аллелей), распределенных по генотипам разных особей, в общий генофонд (аллелофонд) вида. Этот генофонд заключает в себе весь объем наследственной информации, которым располагает вид на определенном этапе его существования.
Определение вида, приведенное выше, не может быть применено к размножающимся бесполым путем агамным (некоторые микроорганизмы, синезеленые водоросли), самооплодотворяющимся и строго партеногетическим организмам. Группировки таких организмов, эквивалентные виду, выделяют по сходству фенотипов, общности ареала, близости генотипов по происхождению. Практическое использование понятия «вид» даже у организмов с половым размножением нередко затруднено. Это обусловлено динамичностью видов, проявляющейся во внутривидовой изменчивости, «размытости» границ ареала, образовании и распаде внутривидовых группировок различного объема и состава (популяций, рас, подвидов). Динамичность видов является следствием действия элементарных эволюционных факторов.
Видообразование
Видообразование – это процесс образования видов, осуществляющийся в результате взаимодействия элементарных эволюционных факторов.
Видообразование в типичных случаях заключается в разделении первоначально единого вида на два или более новых. Это связано с возникновением межпопуляционных изоляционных барьеров и углублением различий между генофондами популяций под действием естественного отбора вплоть до генетической изоляции. Такой процесс, ведущий к увеличению количества видов, называют дивергентным или истинным видообразованием. Выделяют также филетическое видообразование. Оно заключается в постепенном превращении во времени одного вида в другой. Этот способ наблюдается, если изменения условий захватывают весь ареал. Известны примеры видообразования путем гибридизации.
Существуют аллопатрический и симпатрический пути образования видов.
При аллопатрическом видообразовании, называемом также географическим, препятствия к скрещиванию первично обусловлены пространственным разобщением популяций. Генетическая изоляция развивается вторично. Так, некогда в Австралии существовал один вид попугайчиков рода Pachycephala. В условиях засушливого периода единый ареал разделился на западную и восточную зоны. Со временем особи двух популяций приобрели морфофизиологические различия, которые сделали невозможным скрещивание, когда ареал вновь стал общим. Произошло образование из одного предкового вида двух новых.
При симпатрическом видообразовании новый вид образуется внутри ареала исходного вида. С самого начала изоляция является генетической. Такое положение создается в результате полиплоидии вследствие нарушений нормального хода мейоза, при крупных хромосомных перестройках или межвидовой гибридизации. Аллопатрическое видообразование происходит медленно и дает виды, как правило, отличающиеся по морфофизиологическому критерию от вида-родоначальника. Симпатрический путь относительно быстрый и дает виды, близкие к исходному по морфофизиологическим показателям.
Большинство видов, особенно животных, возникают аллопатрическим путем. Симпатрическое видообразование на основе полиплоидии характерно для растений. Так, разные виды пшеницы составляют ряд с наборами 14, 28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, культурного — 52. Культурная слива возникла путем гибридизации терна с алычой. Примером гибридогенного вида является рябинокизильник, распространенный в лесах центральной Сибири. Симпатрический путь видообразования у паразитов часто связан с освоением популяцией новых хозяев. Анализ генного состава и межхромосомных различий между человеком и человекообразными обезьянами дает повод предположить, что разделение этих двух ветвей могло идти симпатрическим путем.
№13
Популяционная структура вида
Видом называют совокупность особей, сходных по основным морфологическим и функциональным признакам, кариотипу, поведенческим реакциям, имеющих общее происхождение, заселяющих определенную территорию (ареал), в природных условиях скрещивающихся исключительно между собой и при этом производящих плодовитое потомство.
Видовая принадлежность особи определяется по соответствию ее перечисленным критериям: морфологическому, физиолого-биохимическому, цитогенетическому, этологическому, экологическому и др. Наиболее важные признаки вида — его генетическая (репродуктивная) изоляция, заключающаяся в нескрещиваемости особей данного вида с представителями других видов, а также генетическая устойчивость в природных условиях, приводящая к независимости эволюционной судьбы.
Со времен К. Линнея вид является основной единицей систематики. Особое положение вида среди других систематических единиц (таксонов) обусловливается тем, что это та группировка, в которой отдельные особи существуют реально. В составе вида в природных условиях особь рождается, достигает половой зрелости и выполняет свою главную биологическую функцию: участвуя в репродукции, обеспечивает продолжение рода. В отличие от вида таксоны надвидового ранга, такие, как род, отряд, семейство, класс, тип, не являются ареной реальной жизни организмов. Выделение их в естественной системе органического мира отражает результаты предшествующих этапов исторического развития живой природы. Распределение организмов по надвидовым таксонам указывает на степень их филогенетического родства.
Важнейшим фактором объединения организмов в виды служит половой процесс. Представители одного вида, скрещиваясь друг с другом, обмениваются наследственным материалом. Это ведет к перекомбинации в каждом поколении генов (аллелей), составляющих генотипы отдельных особей. В результате достигаются нивелировка различий между организмами внутри вида и длительное сохранение основных морфологических, физиологических и прочих признаков, отличающих один вид от другого. Благодаря половому процессу происходит также объединение генов (аллелей), распределенных по генотипам разных особей, в общий генофонд (аллелофонд) вида. Этот генофонд заключает в себе весь объем наследственной информации, которым располагает вид на определенном этапе его существования.
Определение вида, приведенное выше, не может быть применено к размножающимся бесполым путем агамным (некоторые микроорганизмы, синезеленые водоросли), самооплодотворяющимся и строго партеногетическим организмам. Группировки таких организмов, эквивалентные виду, выделяют по сходству фенотипов, общности ареала, близости генотипов по происхождению. Практическое использование понятия «вид» даже у организмов с половым размножением нередко затруднено. Это обусловлено динамичностью видов, проявляющейся во внутривидовой изменчивости, «размытости» границ ареала, образовании и распаде внутривидовых группировок различного объема и состава (популяций, рас, подвидов). Динамичность видов является следствием действия элементарных эволюционных факторов.
Закон Харди-Вайнберга
В пределах генофонда популяции доля генотипов, содержащих разные аллели одного гена; при соблюдении некоторых условий из поколения в поколение не изменяется. Эти условия описываются основным законом популяционной генетики, сформулированным в 1908 г. английским математиком Дж. Харди и немецким врачом-генетиком Г. Вайнбергом. «В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствие мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение».
Допустим, что в генофонде популяции, удовлетворяющей описанным условиям, некий ген представлен аллелями А1 и А2, обнаруживаемыми с частотой р и q. Так как других аллелей в данном генофонде не встречается, то р+q = 1. При этом q = 1—р.
Соответственно особи данной популяции образуют р гамет с аллелем А1 и q гамет с аллелем А2. Если скрещивания происходят случайным образом, то доля половых клеток, соединяющихся с гаметами А1, равна р, а доля половых клеток, соединяющихся с гаметами A2, — q. Возникающее в результате описанного цикла размножения поколение F1 образовано генотипами AlA1, A1A2, A2A2, количество которых соотносится как (р + q) (р + q) = р2 + 2pq + q2. По достижении половой зрелости особи AlAi и АгА2 образуют по одному типу гамет — A1 или A2 — с частотой, пропорциональной числу организмов указанных генотипов (р и q). Особи A1A2 образуют оба типа гамет с равной частотой 2pq/2.
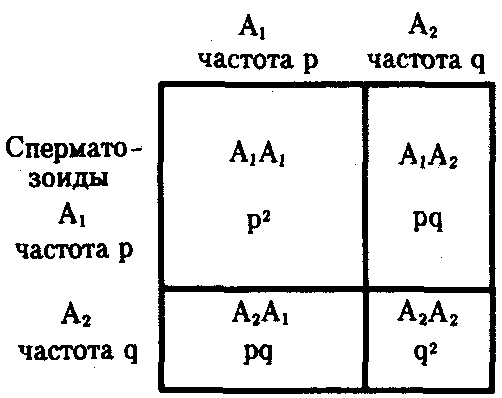
Таким образом, доля гамет A1 в поколении F1 составит р2 + 2pq/2 = р2 + р(1—р) = p, а доля гамет А2 будет равна q2 + 2pq/2 = q2 + + q(l-q) = q.
Так как частоты гамет с разными аллелями в поколении fi в сравнении с родительским поколением не изменены, поколение F2 будет представлено организмами с генотипами AlA1, A1A2 и А2А2 в том же соотношении р2 + 2pq + q2. Благодаря этому очередной цикл размножения произойдет при наличии р гамет A1 и q гамет А2. Аналогичные расчеты можно провести для локусов с любым числом аллелей. В основе сохранения частот аллелей лежат статистические закономерности случайных событий в больших выборках.
Уравнение Харди—Вайнберга в том виде, в котором оно рассмотрено выше, справедливо для аутосомных генов. Для генов, сцепленных с полом, равновесные частоты генотипов AlA1, A1A2 и А2А2 совпадают с таковыми для аутосомных генов: р2 + 2pq + q2. Для самцов (в случае гетерогаметного пола) в силу их гемизиготности возможны лишь два генотипа A1— или А2 —, которые воспроизводятся с частотой, равной частоте соответствующих аллелей у самок в предшествующем поколении: р и q. Из этого следует, что фенотипы, определяемые рецессивными аллелями сцепленных с хромосомой Х генов, у самцов встречаются чаще, чем у самок.
Так, при частоте аллеля гемофилии, равной 0,0001, это заболевание у мужчин данной популяции наблюдается в 10 000 раз чаще, чем у женщин (1 на 10 тыс. у первых и 1 на 100 млн. у вторых).
Еще одно следствие общего порядка заключается в том, что в случае неравенства частоты аллеля у самцов и самок разность между частотами в следующем поколении уменьшается вдвое, причем меняется знак этой разницы. Обычно требуется несколько поколений для того, чтобы возникло равновесное состояние частот у обоих полов. Указанное состояние для аутосомных генов достигается за одно поколение.
Закон Харди — Вайнберга описывает условия генетической стабильности популяции. Популяцию, генофонд которой не изменяется в ряду поколений, называют менделевской. Генетическая стабильность менделевских популяций ставит их вне процесса эволюции, так как в таких условиях приостанавливается действие естественного отбора. Выделение менделевских популяций имеет чисто теоретическое значение. В природе эти популяции не встречаются. В законе Харди — Вайнберга перечислены условия, закономерно изменяющие генофонды популяций. К указанному результату приводят, например, факторы, ограничивающие свободное скрещивание (панмиксию), такие, как конечная численность организмов в популяции, изоляционные барьеры, препятствующие случайному подбору брачных пар. Генетическая инертность преодолевается также благодаря мутациям, притоку в популяцию или оттоку из нее особей с определенными генотипами, отбору.
№14
Популяционная структура человечества
Размножение человека осуществляется половым путем, а репродуктивные ареалы в той или иной степени ограничены определенной группой населения. Это позволяет выделить в человечестве сообщества, аналогичные популяциям в биологическом понимании этого термина. В антропогенетике популяцией называют группу людей, занимающих общую территорию и свободно вступающих в брак. Изоляционные барьеры, препятствующие заключению брачных союзов, нередко носят выраженный социальный характер (например, различия в вероисповедании). Благодаря этому в формировании популяций людей главную роль играет не общность территории, а социальные факторы.
Размер, уровень рождаемости и смертности, возрастной состав, экономическое состояние, уклад жизни являются демографическими показателями популяций людей. Генетически они характеризуются генофондами (аллелофондами). Демографические показатели оказывают серьезное воздействие на состояние генофондов человеческих популяций, главным образом через структуру браков. Большое значение в определении структуры браков имеет размер группы.
Популяции из 1500—4000 человек называют демами, популяции численностью до 1500 человек — изолятами. Для демов и изолятов типичен относительно низкий естественный прирост населения — соответственно порядка 20% и не более 25% за поколение. Частота внутригрупповых браков в них составляет 80—90% и свыше 90%, а приток лиц из других групп сохраняется на уровне 1—2% и менее 1%. В силу высокой частоты внутригрупповых браков члены изолятов, просуществовавших четыре поколения (примерно 100 лет) и более, являются не менее чем троюродными братьями и сестрами (сибсами).
В больших по размерам популяциях распределение аллелей отдельных генов в генотипах индивидуумов последовательных поколений подчиняется закону Харди — Вайнберга. Это используют в медико-генетической практике для расчета доли гетерозигот — носителей определенного рецессивного аллеля. Так, в Швеции в 1965—1974 гг. страдающие фенилкетонурией встречались с частотой примерно 1 : 40000. Исходя из закона Харди — Вайнберга, по локусу, представленному двумя аллелями, три возможных генотипа (AlA1, A1A2 и А2А2) распределяются с частотой р2, 2pq, q2. Следовательно, q2 = 1/40000, a q = 1/200. Частота доминантного аллеля нормального обмена фенилаланина р=1—q=l— 1/200 = 199/200. Тогда частота гетерозигот 2pq = 2 х (1/200) • (199/200) = 2 • (199/40000). При найденных частотах доминантного и рецессивного аллелей популяция численностью 40 000 человек содержит одного больного фенилкетонурией (А2А2) и 400 носителей неблагоприятного аллеля в гетерозиготном состоянии (A1A2). Остальные члены популяции гомозиготны по благоприятному доминантному аллелю (AlA1). Ниже приведены данные о частоте гетерозиготного носительства и соответствующей ей частоте рецессивных гомозигот с фенотипическим проявлением определенного аллеля.
Встречаемость гомозигот (в пересчете на число членов популяции) |
Встречаемость гетерозигот (в пересчете на число членов популяции) |
1 : 10 |
1 : 2,3 |
1 : 100 |
1 : 5.6 |
1 : 1000 |
1 : 16 |
1 : 10 000 |
1 : 51 |
1 : 100 000 |
1 : 159 |
1 : 1 000 000 |
1 : 501 |
Даже по редким рецессивным аллелям количество гетерозигот оказывается достаточно высоким, чтобы это учитывалось при медико-генетическом консультировании вступающих в брак.
Факторы, влияющие на генетическую конституцию людей
Мутационный процесс у человека сходен с таковым у других организмов по всем основным показателям — средней частоте мутирования на локус или геном за поколение, генетико-физиологическим характеристикам мутаций, наличию антимутационных барьеров. Это совпадение неслучайно. Основные характеристики спонтанного мутагенеза формировались на начальных этапах эволюции жизни под действием таких постоянных факторов, как ультрафиолетовое и иные виды излучения, температура, определенная химическая среда.
Хотя оценка частоты возникновения мутаций у людей встречает серьезные трудности, некоторые подходы к получению таких данных имеются. Согласно одному из них, максимальная определяемая вероятность новой мутации составляла 2,24 · 10-5 на один локус в поколении.
В настоящее время давление мутационного процесса на генофонд человечества, по-видимому, усиливается благодаря росту индуцированных мутаций. Их причиной нередко служат факторы, возникающие в связи с производственной деятельностью человека в условиях научно-технической революции, например ионизирующее излучение. Подсчет прироста количества мутаций сверх фоновых значений встречается с теми же трудностями, о которых шла речь выше. Согласно ориентировочным данным, доза в 1 Гр (грей), получаемая при низком уровне радиации мужчинами, индуцирует от 1000 до 2000 мутаций с серьезными фенотипическими последствиями на каждый миллион живых новорожденных. У женщин эта цифра ниже — 900.
Мутагенные факторы индуцируют мутации как в половых, так и в соматических клетках. В последнем случае результат может состоять в повышении частоты определенных заболеваний, прежде всего злокачественных опухолей. В отношении ионизирующих излучений, в частности, это лейкозы. Далее идут рак молочной железы и щитовидной железы.
Дрейф генов и особенности генофондов изолятов
Предположительно человеческие популяции в палеолите состояли из нескольких сотен индивидуумов. Всего одно-два столетия тому назад люди жили преимущественно поселениями в 25—35 домов. Вплоть до самого последнего времени количество индивидуумов в отдельных популяциях, непосредственно участвующих в размножении, редко превышало 400—3500 человек. Причины географического, экономического, расового, религиозного, культурного порядка ограничивали брачные связи масштабами определенного района, племени, поселения, секты. Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала благоприятные условия для дрейфа генов.
Генетико-автоматические процессы, или дрейф генов, приводят к сглаживанию изменчивости внутри группы и появлению случайных, не связанных с отбором различий между изолятами. Именно это выявили наблюдения за особенностями фенотипов малочисленных групп населения в условиях, например, географической изоляции. Так, среди жителей Памира резус-отрицательные индивидуумы встречаются в 2—3 раза реже, чем в Европе. В большинстве кишлаков такие люди составляют 3—5% популяции. В некоторых изолированных селениях, однако, их насчитывается до 15%, т.е. примерно как в европейской популяции.
В крови человека имеются гаптоглобины, которые связывают свободный гемоглобин после разрушения эритроцитов, чем предотвращают его выведение из организма. Синтез гаптоглобина Нр1-1 контролируется геном Нр1. Частота этого гена у представителей двух соседних племен на Севере Южной Америки составляет 0,205 и 0,895, отличаясь более чем в 4 раза.
Примером действия дрейфа генов в человеческих популяциях служит эффект родоначальника. Он возникает, когда несколько семей порывают с родительской популяцией и создают новую на другой территории. Такая популяция обычно поддерживает высокий уровень брачной изоляции. Это способствует случайному закреплению в ее генофонде одних аллелей и утрате других. В результате частота очень редкого аллеля может стать значительной.
Так, члены секты амишей в округе Ланкастер штата Пенсильвания, насчитывающей к середине девятнадцатого века примерно 8000 человек, почти все произошли от трех супружеских пар, иммигрировавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев особой формы карликовости с многопалостью, которая наследуется по аутосомно-рецессивному типу. Эта аномалия не зарегистирирована среди амишей штатов Огайо и Индиана. В мировой медицинской литературе описано едва ли 50 таких случаев. Очевидно, среди членов первых трех семей, основавших популяцию, находился носитель соответствующего рецессивного мутантного аллеля — «родоначальник» соответствующего фенотипа.
Последствия дрейфа генов, представляющие интерес для медицины, заключаются в неравномерном распределении по группам населения Земного шара некоторых наследственных заболеваний. Так, изоляцией и дрейфом генов объясняется, по-видимому, относительно высокая частота церебромакулярной дегенерации в Квебеке и Ньюфаундленде, детского цестиноза во Франции, алкаптонурии в Чехии, одного из типов порфирии среди европеоидного населения в Южной Америке, адреногенитального синдрома у эскимосов. Эти же факторы могли быть причиной низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
Изменение генетического состава популяции вследствие генетико-автоматических процессов приводит к гомозиготизации индивидуумов. При этом чаще фенотипические последствия оказываются неблагоприятными. Вместе с тем следует помнить, что возможно образование и благоприятных комбинаций аллелей. В качестве примера рассмотрим родословные Тутанхамона и Клеопатры VII, в которых близкородственные браки были правилом на протяжении многих поколений.
Тутанхамон умер в возрасте 18 лет. Анализ его изображения в детском возрасте и подписи к этому изображению позволяют предположить, что он страдал генетическим заболеванием — целиакией, которая проявляется в изменении слизистой оболочки кишечника, исключающем всасывание клейковины. Тутанхамон родился от брака Аменофиса III и Синтамоне, которая была дочерью Аменофиса III. Таким образом, мать фараона была его сводной сестрой. В могильном склепе Тутанхамона обнаружены мумии двух, по всей видимости мертворожденных, детей от брака с Анкесенамон, его племянницей. Первая жена фараона была или его сестрой, или дочерью. Брат Тутанхамона Аменофис IV предположительно страдал болезнью Фрелиха и умер в 25—26 лет. Его дети от браков с Нефертити и Анкесенамон (его дочерью) были бесплодны. С другой стороны, известная своим умом и красотой Клеопатра VII была рождена в браке сына Птоломея Х и его родной сестры, которому предшествовали кровнородственные браки на протяжении по крайней мере шести поколений.
№15
Учение академика А.Н.Северцова о направлениях эволюционного процесса
Рассматривая эволюцию отдельных таксонов, можно убедиться в том, что некоторые из них находятся в состоянии расцвета, в то время как другие вымирают. Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса, критериями которого являются:
увеличение количества представителей соответствующей группы;
расширение ареала распространения;
активизация видообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д.
Явление, противоположное биологическому прогрессу, — биологический регресс — свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию. Так же как и биологический прогресс, состояние регресса может длиться очень долго. Оставшиеся немногочисленные представители некогда биологически прогрессивной группы, найдя соответствующую их организации экологическую нишу, могут продолжать существовать на протяжении сотен миллионов лет. Они называются реликтами, обладают консервативностью организации и крайне медленно эволюционируют. Примерами реликтов являются современная кистеперая рыба — латимерия, ближайшие родственники которой жили в девонском периоде более 200 млн. лет назад, а также дерево гинкго, непосредственные предки которого существовали на Земле уже в первой половине мезозойской эры.
А.Н.Северцов связывал морфофизиологический прогресс с повышением дифференциации организма и интенсификацией функций (в частности, с повышением интенсивности обменных процессов и энергии жизнедеятельности организма). Позднее различными учеными к этим двум критериям арогенеза были добавлены другие. Наиболее важными из них являются: усовершенствование интеграции организма; рационализация его устройства, т. е. упорядочение организации, ведущее к оптимизации функционирования; повышение уровня гомеостаза, т. е. способности поддерживать постоянство внутренней среды организма; возрастание объема информации, извлекаемой организмом из внешней среды, и совершенствование ее обработки и использования. Другие критерии арогенеза, предлагавшиеся разными учеными, либо являются частными производными от перечисленных основных его критериев, либо не всегда коррелируют с морфофизиологическим прогрессом.
Конкретные проявления морфофизиологического прогресса в разных филогенетических стволах существенно различаются в соответствии с исторически сложившимися особенностями организации разных групп и различным характером взаимоотношений с внешней средой. Поэтому часто оказывается трудно сравнивать уровни морфофизиологического прогресса, достигнутые разными группами. Ч.Дарвин полагал даже, что «попытки сравнивать в отношении высоты организации представителей различных типов совершенно безнадежны; кто решит, кто выше, каракатица или пчела». Действительно, необходимые для подобных сравнений методы количественной оценки высоты организации еще не разработаны. Однако на качественном уровне сравнение по основным критериям арогенеза все же возможно: едва ли может возникнуть сомнение, что общий уровень организации насекомых выше, чем, например, у многоножек, и у любых членистоногих он выше, чем у кольчатых червей, а у последних — выше, чем у плоских червей, и т. п.
При этом важно подчеркнуть, что лишь совокупность названных основных критериев, не менее четырех из них, характеризует общий морфофизиологический прогресс (арогенез), т. е. повышение организации в филогенезе данной группы организмов. Арогенез происходил, например, в филогенезе позвоночных, членистоногих, моллюсков; среди таксонов меньшего ранга общий арогенез характерен для млекопитающих, птиц, насекомых, головоногих моллюсков.
Так же, Северцов предлагал различать:
Ароморфоз — прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов.
Идиоадаптация (алломорфоз) - малозначимые эволюционные изменения, связанные с мелкими приспособлениями к конкретным условиям окружающей среды.
Общую дегенерацию - регрессивные изменения (катаморфоз).
В качестве примеров ароморфозов или ароморфных изменений можно взять преобразования кровеносной системы у позвоночных (формирование четырёхкамерного сердца и разделение большого и малого кругов кровообращения), концентрацию нервной системы с образованием нервных узлов (ганглиев) и т. п. Н. А. Северцов полагал, что ароморфозы были нечастыми эволюционными событиями, и большая часть изменений должна квалифицироваться как идиоадаптации (в том числе, например, специализация конечностей наземных позвоночных, включая преобразование их в крылья).
С точки зрения современных представлений о ходе эволюционного процесса, нет никаких оснований полагать, что имеют место качественно отличные друг от друга изменения, которые Н. А. Северцов называл ароморфозами и идиоадаптациями.
Вместе с тем, некоторые биологи в России продолжают держаться взглядов Н. А. Северцова. Те, кто считает это различение продуктивным, полагают, что идиоадаптации выражаются в изменениях формы тела, избыточной степени развития или редукции отдельных органов. Ароморфозы же, в большинстве случаев, связывают с появлением новых структур, качественными изменениями в эмбриональном развитии. Ароморфоз позиционирует как главный скоростной двигатель прогресса. При ароморфозе животные начинают эволюционировать в ускоренном темпе. Эволюционное значение ароморфоза — сохранение в ходе дальнейшей эволюции, что и приводит к возникновению крупных систематических групп: типов, отделов, классов, некоторых отрядов (у млекопитающих).
№16
Индивидуальное и историческое развитие
Геккель в 1866 г. сформулировал биогенетический закон: «Онтогенез – есть краткое проявление филогенеза». Что подтверждает наличие некоторых признаков у зародышей в процессе онтогенеза:
Признаки взрослых предков, которые проявляются у потомков (палингенезы).
Признаки, являющиеся новообразующимися для приспособления к эмбриональным и личиночным стадиям (ценогенезы).
Так же, Геккель предложил изучение филогенеза различных органов используя сравнительно-анатомический, палеонтологический и эмбриологический методы, чтобы судить об эволюционном развитии в целом. Этот метод является основным в эволюционной морфологии и называется методом тройного параллелизма («Триада Геккеля»).
Сущность взглядов А. Н. Северцова: "Филогенетическое изменение строения взрослых органов происходит путем изменения эмбрионального развития этих органов. Филогенез является, таким образом, функцией онтогенеза". К сожалению, до сих пор остается неясным, "как" все происходит, а "период" определен слишком в общем виде. Северцов выделил такие виды филэмбриогенезов: архаллаксис, девиация, анаболия.
1. Анаболии, или надставки, возникают после того, как орган практически завершил свое развитие (т.е. изменение происходит на конечных стадиях эмбриогенеза), и выражаются в добавлении дополнительных стадий, изменяющих конечный результат.
К анаболиям относят такие явления, как приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, неотличимый от других рыб, а также появление изгибов позвоночника, сращение швов в мозговом черепе, окончательное перераспределение кровеносных сосудов в организме млекопитающих и человека.
2. Девиации — уклонения, возникающие в процессе морфогенеза органа (в эмбриональном периоде изменение происходит на средних стадиях эмбриогенеза). Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих . В развитии легких у млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее морфогенез идет по-новому.
3. Архаллаксисы — изменения, которые могут выражаться в изменении некоторых процессов дифференцировки начальной массы, сдвигах места закладки органов (гетеротопия) или времени закладки органов (гетерохромия). В эмбриональном периоде изменение происходит на ранних стадиях. Классическим примером архаллаксиса является развитие волос у млекопитающих, закладка которых наступает на очень ранних стадиях развития и с самого начала отличается от закладок других придатков кожи позвоночных.
По типу архаллаксиса возникают хорда у примитивных бесчерепных, хрящевой позвоночник у хрящевых рыб, развиваются нефроны вторичной почки у пресмыкающихся.
№17
Органический мир как результат процесса эволюции
№18
Общие закономерности филогенеза
Эволюционная морфология как наука объединяет данные сравнительной анатомии, палеонтологии и эмбриологии. Она изучает организации живого, её место в эволюционном ряду, устанавливает родственные взаимосвязи между разными формами животного мира. Она изучает пути происхождения форм животного мира, восстанавливает промежуточные формы и открывает новые.
Методы изучения филогенеза предложил Геккель: изучать филогенез различных органов используя сравнительно-анатомический, палеонтологический и эмбриологический методы, чтобы судить об эволюционном развитии в целом. Этот метод является основным в эволюционной морфологии и называется методом тройного параллелизма («Триада Геккеля»).
Основные понятия
Если у двух организмов, находящихся на разных уровнях организации, обнаруживаются органы, которые построены по единому плану, расположены в одинаковом месте и развиваются сходным образом из одинаковых эмбриональных зачатков, то это свидетельствует о родстве данных организмов. Такие органы называют гомологичными. Гомологичные органы часто выполняют одну и ту же функцию (например, сердце рыбы, земноводного, пресмыкающегося и млекопитающего), но в процессе эволюции функции могут и меняться (например, передних конечностей рыб и земноводных, пресмыкающихся и птиц).
При обитании неродственных организмов в одинаковых средах у них могут возникать сходные приспособления, которые проявляются в возникновении аналогичных органов. Аналогичные органы выполняют одинаковые функции, строение же их, местоположение и развитие резко различны. Примерами таких органов являются крылья насекомых и птиц, конечности и челюстной аппарат членистоногих и позвоночных.
Гомойология - приобретённое под влиянием сходных условий жизни морфологического или физиологического сходство органов или признаков у животных далёких систематических групп (например, гребень грудной кости у птиц и крота, ласты ихтиозавра, плезиозавра и кита). Внешне такие органы сходны с истинными гомологичными органами, хотя в действительности это аналогичные органы.
Гомодинамия - сходство органов или частей, расположенных последовательно на продольной оси тела животного. Термин гомодинамия предложен немецким биологом Э. Геккелем в 1866. Примеры гомодинамии: позвонки, ребра, передние и задние конечности позвоночных, сегменты тела членистоногих со всеми их частями, в том числе конечностями. Если гомодинамные органы приспособлены к выполнению различных функций, они могут иметь неодинаковое строение (например, крылья и ноги птиц).
Главные принципы эволюции
Изменчивость и наследственность – в следствии изменчивости признаков и свойств даже в потомстве одной пары родителей почти не встречается одинаковых особей. При благоприятных условиях эти различия могут не играть существенной роли, при неблагоприятных – каждое мельчайшее событие может стать решающим в том, останется ли этот организм в живых и даст потомство или же будет уничтожен.
Размножение в геометрической прогрессии – потенциально вид в каждом поколении производит гораздо больше особей, чем их может выжить до взрослого состояния на занимаемой территории. Следовательно, значительная часть родившихся гибнет в «борьбе за жизнь». Отсюда вытекает следующий принцип.
Борьба за существование. Дарвин выделял 3 формы борьбы за существование:
Внутривидовая борьба – взаимоотношения между особями разного пола, между различными поколениями, борьба в стае и т.д.
Межвидовая борьба – конкуренция в местах обитания, размножения, взаимоотношения из-за пищи и т.д.
Борьба с неблагоприятными условиями – возникает в зависимости от климатических условий, влияющих на жизнедеятельность организма.
Естественный отбор – это (по Дарвину) совокупность происходящих в природе событий, обеспечивающих выживание наиболее приспособленных, и преимущественное оставление ими потомства.
Мультифункциональность органов и структур, принцип, согласно которому каждый орган или структура выполняет в организме более чем одну функцию одновременно или в разные периоды его индивидуального развития, или онтогенеза. Так, крыло летучей мыши не только служит для полёта, но и помогает при ловле добычи (действуя по принципу сачка), осуществляет терморегуляцию тела, кожная перепонка его участвует в продукции витамина D и т. д. Мультифункциональность характерна также для органоидов тканевых клеток и органелл простейших. Принцип мультифункциональности лежит в основе всех изменений органов и структур в процессе эволюции.
Количественные функциональные изменения органов
1. Расширение функций. Например, уши у слона служит дополнительно органом терморегуляции; кровеносная система выполняет функцию терморегуляции и защитную функцию.
2. Сужение функций. Например, конечности лошади утратили лазающую и хватательные функции. Сужение функций часто связано с их иммобилизацией – утрате функций в связи с редукцией органа.
3. Интенсификация функций. Например, увеличение переднего мозга привело к формированию второй сигнальной системы; развитие шерстного покрова обеспечило и терморегуляцию, и защиту от физико-химических повреждений. Интенсификация функций часто связана с их активацией – преобразованием пассивного органа в активный. Примеры: втяжные когти кошачьих, подвижные челюсти змей, использование метаболической воды обитателями степей и пустынь.
№19
Способы морфофункционального преобразования органов
Один из основных принципов эволюции органов — принцип расширения и смены функций. Расширение функций сопровождает обычно профессивное развитие органа, который по мере дифференциации выполняет все новые функции. Так, парные плавники рыб, возникшие как пассивные органы, поддерживающие тело в воде в горизонтальном положении, с приобретением собственной мускулатуры и прогрессивным расчленением становятся еще и активными рулями глубины и поступательного движения. У придонных рыб они обеспечивают также их передвижение по дну. С переходом позвоночных на сушу к перечисленным функциям конечностей добавились хождение по Земле, лазание, бегание и др.
Расширение функций сопровождается специализацией, благодаря которой главной функцией становится одна из бывших ранее второстепенными. Бывшая главной функция преобразуется во второстепенную и может впоследствии даже исчезнуть. Орган при этом меняется таким образом, что его строение становится максимально соответствующим выполнению главной функции. Так, переход предков ластоногих и китообразных к водному образу жизни привел к преобразованию их парных конечностей в ласты, практически утратившие способность обеспечивать передвижение по суше. Жизнь ленивцев, представителей отряда неполнозубых, на деревьях привела к формированию у них крючкообразных конечностей, с помощью которых возможно лишь медленное перемещение по веткам в подвешенном состоянии с почти полной утратой способности движения по земле.
Нередко функции, выполняемые органами, могут измениться кардинально. Так, плавательный пузырь рыб, будучи гидростатическим органом, у кистеперых рыб становится дополнительным органом дыхания, а у земноводных он преобразуется в легкое, и основной функцией его становится дыхательная. У пресмыкающихся и млекопитающих, ведущих наземный образ жизни, легкие выполняют только дыхательную функцию, но первичная функция плавательного пузыря сохраняется за легкими у крокодилов, ластоногих и китообразных, ведущих водный образ жизни, а также у наземных форм во время плавания.
В других случаях видоизменения органов в связи со сменой их функций столь велики, что выполнение ими функций, бывших ранее главными, становится невозможным. Так, передние жаберные дуги предков хрящевых рыб преобразовались в челюсти, а у наземных позвоночных они стали выполнять функции звукопроводящего аппарата, превратившись в слуховые косточки. Участие их в пищеварении и дыхании стало невозможным.
В прогрессивной эволюции органов очень важным является принцип активации функций. Он наиболее часто реализуется на начальных этапах эволюции органов в том случае, когда малоактивный орган начинает активно выполнять функции, существенно при этом преобразуясь.
Так, крайне малоподвижные парные плавники хрящевых рыб становятся активными органами движения уже у костистых.
Более часто в филогенезе наблюдается интенсификация функций, являющаяся следующим этапом эволюции органов после активации. Благодаря этому орган обычно увеличивается в размерах, претерпевает внутреннюю дифференцировку, гистологическое строение его усложняется, нередко наблюдается многократное повторение одноименных структурных элементов, или полимеризация структуры. Примером является усложнение структуры легких в ряду наземных позвоночных за счет ветвления бронхов, появления ацинусов и альвеол на фоне постоянной интенсификации его функций.
Высокая степень дифференцировки может сопровождаться уменьшением количества одинаковых органов, выполняющих одну и ту же функцию, или их олигомеризацией. Это явление наблюдается, к примеру, в эволюции артериальных жаберных дуг, которые закладываются у хрящевых рыб в количестве 6—7 пар, у костных рыб их становится 4 пары, а у млекопитающих и человека сохраняются в дефинитивном состоянии лишь части 3, 4 и 6-й пар.
В ходе эволюции часто наблюдается субституция – замещение одного органа другим или передача функций от одного органа к другому. Различают субституцию органов и субституцию функций.
Субституция органов, или гомотопная субституция – замещение в ходе эволюции одного органа другим, занимающим сходное положение в организме и выполняющим биологически равноценную функцию. В этом случае происходит редукция замещаемого органа и прогрессивное развитие замещающего. Так, у хордовых осевой скелет – хорда – замещается сначала хрящевым, затем костным позвоночником. В ряде случаев субституция приводит к появлению аналогичных органов, например, у растений листья (фотосинтезирующие органы) замещаются филлодиями (уплощенными черешками) или филлокладиями (уплощенными стеблями). Термин «субституция органов» введён Н. Клейненбергом (1886).
Субституция функций, или гетеротопная субституция – утрата в ходе эволюции одной из функций (при этом выполнявший её орган редуцируется) и замещение её другой, биологически равноценной (выполняемой другим органом). Так, функция перемещения тела в пространстве при помощи ног (хождение) у змей замещена перемещением при помощи изгибаний позвоночника (ползание); дыхание с помощью жабр (извлечение кислорода из воды) у наземных позвоночных замещено газообменом в лёгких. Термин «субституция функций» введён А. Н. Северцовым (1931).
Субституция тесно связана с принципом компенсации и с редукцией органов. Например, у птиц редукция зубов связана с развитием мускулистого желудка.
В противоположность интенсификации и активации ослабление функций ведет в филогенезе к упрощению строения органа и его редукции, вплоть до полного исчезновения.
Гетеробатмия - неодинаковый уровень развития и специализации различных органов, достигнутый в результате относит, независимости в развитии разных частей организма в процессе эволюции.
№20
Антропогенез (Положение человека в системе животного мира)
Положение человека в эволюционной лестнице:
царство Животные
тип Хордовые
класс Млекопитающие
отряд Приматы
подотряд Гаплориновые
инфроотряд Узконосые обезьяны
надсемейство Гоминоиды
семейство Гоминиды
род Собственно Люди
Вид Неоантроп
подвид Современный человек
Биосоциальная природа человека
В процессе антропогенеза (эволюционно-исторического процесса формирования человека) произошло возникновение на нашей планете высшей формы существования материи - социальной. Появление человека является результатом развития жизни в одной из ее эволюционных ветвей - царстве животных. Биологический вид Человек разумный (Homo sapiens) представляет собой уникальную жизненную форму, соединяющую в себе биологическую и социальную сущности. Это соединение обусловлено длительным процессом биологической эволюции и исторического развития человечества. Социальность не противопоставляет людей остальной природе. Соответственно своей биологической организации человек безусловно принадлежит живой природе, царству животных. Жизнедеятельность человеческого организма основывается на фундаментальных биологических механизмах, которые составляют биологической наследство человека. Именно поэтому он включен в систему природы, которая складывалась на протяжении большей части истории развития планеты независимо от социального фактора и сама же породила этот фактор. Человек составляет своеобразный и неотъемлемый компонент биосферы. Особенность биологической сущности человека заключается в том, что она проявляется в условиях действия законов высшей, социальной формы движения материи. Из социальной сущности людей вытекают закономерности и направления исторического развития человечества. Биологические процессы с необходимостью происходят в организме человека и им принадлежит фундаментальная роль в обеспечении важнейших сторон жизнеспособности и развития. Тем не менее, в популяциях людей эти процессы не приводят к результатам, обычным для остального мира живых существ. Так, естественный отбор - движущий фактор эволюции живых организмов - утратил свое значение (например, в видообразовании) в развитии человека, уступив ведущую роль социальным факторам. Процесс индивидуального развития человека базируется на информации двух видов. Первый вид представляет собой биологически целесообразную информацию, которая отбиралась и сохранялась в процессе эволюции предковых форм и зафиксирована в виде генетической информации в ДНК (универсальный для всех живых организмов механизм кодирования, хранения, реализации и передачи из поколения в поколение информации). Благодаря ей, в индивидуальном развитии человека складывается уникальный комплекс структурных и общения в социуме. Данная особенность человека определяется понятием «социальной наследственности», присущей исключительно человеческому обществу. функциональных признаков, отличающих его от других живых организмов. Второй вид информации представлен суммой знаний, умений, которые создаются, сохраняются и используются поколениями людей в ходе развития человеческого общества. Освоение этой информации индивидуумом происходит в процессе его воспитания, обучения и общения в социуме. Данная особенность человека определяется понятием «социальной наследственности», присущей исключительно человеческому обществу. функциональных признаков, отличающих его от других живых организмов. Второй вид информации представлен суммой знаний, умений, которые создаются, сохраняются и используются поколениями людей в ходе развития человеческого общества. Освоение этой информации индивидуумом происходит в процессе его воспитания, обучения и общения в социуме. Данная особенность человека определяется понятием «социальной наследственности», присущей исключительно человеческому обществу.
№21
Антропогенез (Человек – продукт эволюции)
Сравнительная морфология даёт много фактов, свидетельствующих о происхождении человека от высших животных:
сходство его эмбрионального развития с развитием человекообразных обезьян
наличие рудиментов (зубы мудрости, аппендикс, мышцы уха и др.)
возможность атавизмов (чрезмерное развитие волосяного покрова, дополнительные соски и др.)
у всех приматов 1ый палец руки противопоставлен другим, имеются ногти, пара молочных желёз, хорошо развитые ключицы.
Доказательства из области сравнительной физиологии:
Кровь человека и шимпанзе близки по своим физиологическим и биохимическим показателям.
Одинаковый порядок прорезывания зубов.
Позднее половое созревание (у обезьян в 8-12 лет).
Одинаково высоко развиты формы заботы о потомстве.
Доказательства из области сравнительной патологии и паразитологии:
Возможность развития одинаковых форм инфекционных и вирусных заболеваний (грипп и др.)
Наличие общих видов паразитов.
Цитогенетические и биохимические доказательства:
99% белков идентичны.
Кариотип отличается по количеству на 1ну пару.
13 пара хромосом идентична. Метод дифференцированного окрашивания указывает на наличие ряда хромосомных мутаций, которые в кариотипе человека изменили лишь местоположение большинства хромосомных участков.
Становление человека как вида происходило в 4ре этапа:
Предшественники человека – проантропы.
Древнейшие люди – архантропы.
Древние люди – палеоантропы.
Человека современного типа – неоантроп.
Филогенез семейства гоминид представляет собой разветвленное дерево, где первая ветвь достигла человека.
Это группа родственных видов возникла в конце мезозоя – начале палеозоя (примерно 70-40 млн. лет назад). Эволюция приматов привела в появлению ветви древнейших человекоподобных обезьян (парапитеков).
д ревнейшие
человекоподобные обезьяны (парапитеки)
ревнейшие
человекоподобные обезьяны (парапитеки)
м алые человекоподобные обезьяны (гиббоны) дреопитеки (древесные обезьяны)
С охранили древесные формы жизни Освоили открытые места обитания
Освобождение рук от участия в передвижении
I этап. Предшественники человека.
В основании теории развития линии приматов, приведших к образованию человека, находятся австралопитеки. Их останки были обнаружены в Южной Африке в 30х годах в пустыне Калахари. Их возраст примерно от 1 до 4 млн. лет. Они зарактеризуются:
широкая зубная дуга и длинная с крупными коренными и редуцированными клыками челюсть
лицо прогнатное
объём мозга 350-400 мл.
масса мозга 450-550 г.
рост 100-160 см.
масса тела 20-50 кг.
Модели: по вертикальному строению, строению таза и суставов ног можно судить о двуножном передвижении гоминид. Жили группами (были кочевниками) по 20-30 особей. Предполагали, что у них была достаточно сложная система взаимоотношений, иерархией, воспитанием детей и попыткой изъясняться с помощью голоса. В качестве орудий применялись палки и камни.
Среди прогрессирующих австралопитеков выделяют человек умелый, появившийся около 2х млн. лет назад в Африке. Отличается большой массой головного мозга (до 650 г), на внутренней поверхности черепа обнаружены следы нейрофизиологических изменений, которые свидетельствуют о выраженной асимметрии больших полушарий и развитии 2х речевых центров.
II этап. Древнейшие люди.
Представители – «человек выпрямленный». Благодаря ярко выраженной социальности этот вид быстро расселился по всей Африке и Евразии. По сравнению с человеком умелым имел более крупное телосложение (рост 160-170 см.), имел большую ёмкость черепа (около 1030 см3), массу мозга (800-1000 г.). Становится возможным речь. Переход к прямохождению сопровождается ароморфозом. Освобождается передняя конечность и превращается в орган труда. Ведут стадный образ жизни. Появляются каменные орудия труда. На этой стадии начинается дифференцировка гоминид на внутривидовые формы. Из них наиболее известны:
питекантроп (о. Ява)
синантроп (китай)
гейдергбергский человек (Германия)
атлантроп (Алжир)
олдовайский питекантроп (Центральная Африка).
III этап. Древние люди.
Становится наиболее распространённым видом около 100-30 тысяч лет назад. По месту 1ого описания их назвали Неандертальцами. Населяли приледниковую зону Европы. Их облик свидетельствует о приспособленности к суровому климату (широкие плечи при малорослости (160см.), сильноразвитая мускулатура, массивный скелет, череп удлиненной формы с массивным надглазничным валиком и низким сводом). Масса мозга примерно 1500 г., причём сильнее всего развиты получили логические участки мозга. Использовались орудия труда, шкуры для одежды, высокий уровень охоты, возникает культура захоронений.
Около 40-50 тысяч лет назад эволюционные преобразования привели к появлению человека разумного.
IV этап. Современные люди.
Ископаемые останки были найдены во Франции в гроте Кроманьён, откуда и получили название Кроманьёльцы.
Вопрос о формирования человека как вида не решён. С одной стороны по теории моноцентризма – неоантроп возник в единой области о 1ой группы палеоантропов. Полицентристы считают, что каждая раса приобрела черты современного человека независимо от других.
В итоге была предложена теория широкого моноцентризма (профессор Рогинский):
«Современный человек сформировался в какой-то области старого света и распространился к периферии, смешиваясь с местными формами других людей».
Кроманьёльцы характеризуются:
значительное развитие всех отделов головного мозга (речь, трудовая деятельность)
Становление человека разумного связанно с формированием устойчивого морфологического тапа.
быстрый рост культуры.
№22
Антропогенез, учение о расах
В человеческом обществе популяцией называют группы людей, занимающую общую территорию и свободно вступающую в брак.
Популяции человека различаются как по численности, так и по характеру организации:
Изоляты – это простые сообщества до 1500 человек.
Демы – это сообщества от 150о до 4 тысяч человек.
Высшие популяционные комплексы - от 4 тысяч человек.
Расы – это исторически сложившиеся группы популяций людей разной численностью, характеризующиеся сходством морфологических и физиологических свойств, а так же общностью занимаемой ими территории.
Адаптационный тип – это карта биологических реакций на комплекс условий среды обитания, обеспечивающая состояние равновесия популяции со средой. Находят выражения в морфофункциональных особенностях популяций и не зависят от расы и этнической принадлежности.
Расы
На протяжении длительного времени в антропологии господствовали представления о значимости расовой дифференцировки человечества и о большой роли естественного отбора в формировании основных расовых признаков. Применение методов молекулярной антропологии в значительной степени изменило представление о расах и расогенезе.
Морфологические и в меньшей степени физиологические признаки дают возможность выделить внутри человечества три основные большие расы: европеидную, австрало-негроидную и монголоидную.
Европеоиды имеют светлую или смуглую кожу, прямые или волнистые волосы, узкий выступающий нос, тонкие губы и развитый волосяной покров на лице и теле. У монголоидов кожа также может быть как светлой, так и темной, волосы обычно прямые, жесткие, темно пигментированные, косой разрез глаз и эпикант («третье веко»). Негроиды характеризуются темной кожей, курчавыми или волнистыми волосами, толстыми губами и широким, слегка выступающим носом. Имеются отличия рас и по некоторым физиологическим и биохимическим показателям: интенсивность потоотделения с единицы площади кожи у негроидов выше, чем у европеоидов, средние показатели уровня холестерина в плазме крови наиболее велики у европеоидов.
В рамках каждой большой расы выделяются отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми расами. Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и на основе популяционной концепции. В соответствии с первым подходом три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга. Несмотря на то что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой — об условности членения человечества даже на большие расы.
На основании исследований распределения различных групп крови и белков в популяциях человека произведено сравнение доли каждого из четырех уровней меж- и внутрипопуляционных различий в общем объеме генетического полиморфизма человека по этим признакам.
Таким образом, от тотального генетического полиморфизма человечества расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдельных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем к другому немцу, живущему в соседней квартире. Изучение геногеографии популяций человека показало, что географическое распределение частот генов групп крови системы АВ0, MN, Lutheran, Duny, Diego и др., а также различных форм ферментов и иммуноглобулинов не соответствует ареалам расселения ни одной из рас. Так, по группам крови АВ0 и MN жители Европы оказываются ближе к африканцам, в то время как по системе иммуноглобулинов они ближе к монголоидам Азии. Сходные результаты получены и в отношении распределения в популяциях вариантов митохондриальной ДНК.
Эти данные свидетельствуют о том, что биохимический полиморфизм человека эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики являются не более чем отдельными проявлениями общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда.
На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов (0,17—0,22), и еще более мала по сравнению с межвидовым расстоянием (0,5—0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу.
№23
Человек и биосфера
№24
Экология человека
Проблема приспособления человека к социальным условиям существования также находится в центре внимания многих естественных и общественных наук, изучающих жизнедеятельность человека во всех формах ее проявления.
В.П. Казначеев определяет популяцию как «сложную социально-биологическую общность людей». Основой существования человеческой популяции является социально-трудовая активность людей в отличие от инстинктивно-поведенческого способа существования популяций животных. Биологическое приспособление животных организмов к изменениям условий развивающейся по законам природы биосферы опосредуется действующими в социуме закономерностями, в результате биологическое содержание приспособительных механизмов проявляется в «снятом» виде. В первую очередь это проявляется в том, что биологические механизмы адаптивно-компенсаторных процессов трансформируются в социально-биологические.
Изначально возникшая в результате биологических эволюционных процессов в биосфере человеческая популяция в процессе социального развития выделилась из биосферы как наиболее активный ее элемент, преобразующий биосферу в ноосферу, дальнейшая эволюция которой подчиняется закономерностям, определяющим социальное развитие общества.
С появлением человека биологическая эволюция перешла на высший уровень эволюции – социальный. Ведущую роль в развитии заняло общество мыслящих людей вместо популяции биологических организмов. Это явилось следствием того, что информационные взаимодействия животных со средой, обеспечивающие ранее пассивное приспособление организмов к изменениям среды на основе опережающего отражения, сменились принципиально новым уровнем информационных процессов, в которых генетическая информация опосредовалась социальной информацией – трудовым опытом людей. В результате человек приобрел возможность активного воздействия на среду своего существования – биосферу. Возникновение второй сигнальной системы – речи – привело к тому, что накопление концентрации энергии живым веществом биосферы за счет обмена организмов со средой веществом дополнилось накоплением социальной информации, позволившим регулировать обменные процессы между обществом и природой.
По мере накопления информации о закономерностях природных процессов человек получил возможность технически использовать природные ресурсы, осуществлять физические и химические методы превращения энергии природных процессов в энергию, потребную для прогрессивного развития человеческого общества. В биосфере происходит становление ее нового компонента – техносферы. Основной характеристикой техносферы как этапа перехода биосферы в ноосферу можно считать возрастающее антропогенное воздействие в глобальных масштабах на процессы, происходящие в биосфере.
Уровни экологических связей человека:
Индивидуальный – это уровень, показывающий отношение каждого человека к экологии окружающей среды (количество выбрасываемого мусора на 1 человека и так далее).
Групповой – уровень отношения группы или нескольких групп людей к окружающей среде (отношение одной или нескольких семей к окружающей среде).
Глобальный – суммарное действие человечества на окружающую среду в целом.
№25
Экологическая дифференциация человечества
Адаптационный тип – это карта биологических реакций на комплекс условий среды обитания, обеспечивающая состояние равновесия популяции со средой. Находят выражения в морфофункциональных особенностях популяций и не зависят от расы и этнической принадлежности.
Среда обитания может быть представлена 10ю основными экосистемами:
Экваториальные леса
Тропические лека и кустарники.
Тропические степи (саванны)
Пустыни и засушливые области.
Леса умеренного пояса.
Леса и кустарники средиземноморья.
Степи умеренного пояса.
Северные леса (тайга).
Полярные области и тундра.
Горные районы.
Каждая из экосистем неодинакова и зависит от особенностей абиотический и биотических компонентов в каждой. Максимальная численность человека представлена во 2 и 5. Разнообразие этих систем привело к адаптивным типам.
Характеристика нескольких типов:
Тропические широты – удлиненная форма тела, пигментная кожа, повышенное количество потовых желёз, специфичный водно-солевой обмен, сниженный процесс метаболизма и холестерина.
Горные районы – крупные размеры тела, мощно развитый скелет и в частности грудная клетка, гипертрофия красного мозга, повышенный уровень эритропоэза, увеличенный жизненный объём лёгких и теплопродуктивность, повышенное содержание гемоглобина.
Арктические зоны – небольшой рост, масса тела увеличена, увеличенные размеры талии и ягодиц, повышенное жироотложение, основной обмен веществ повышен на 16%, сдвиг лейкоцитарной формулы в сторону нейтрофилов, повышено содержание иммуноглобулина.
Умеренный пояс – по морфологическим и функциональным свойствам занимают среднее положение между жителями Арктики и тропиков.
№26
Филогенез кровеносной системы позвоночных
У ланцетника кровеносная система наиболее проста. Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями. На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд — печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, — в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника.
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд — артериальный конус, разветвляющийся последовательно на три пары сосудов:
кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами — передними полыми венами.
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой — легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных.
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены — рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими.
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным. Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком. Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфогенезе сердца большое значение имеют онтогенетические корреляции. Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
Филогенез артериальных жаберных дуг:
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных. В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального, или боталлова протока, представляющего собой часть корня спинной аорты между 4-й и'6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке. Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов — отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
№27
Филогенез пищеварительной системы
Уникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными. Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum.
Ротовая полость
Рассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания.
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть. Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.
Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб. У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости. Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом.
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом.
У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе. Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.
язык, который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого. Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе.
Слюнные железы у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей—и токсины. У млекопитающих слюнные железы многочисленны: это и мелкие — зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные — подъязычные, подчелюстные и околоушные. Подъязычная и подчелюстная железы — результат дифференцировки подъязычной железы пресмыкающихся, околоушная — новое приобретение млекопитающих.
Средняя и задняя кишка
Кишечная трубка претерпевает в ходе филогенеза следующие прогрессивные преобразования: увеличение общей длины, дифференцировку на отделы и образование крупных многоклеточных желез.
Если длина этого отдела пищеварительной трубки ланцетника составляет примерно 1/3 длины тела, то у млекопитающих возможно превышение ее длины по отношению к телу в 10 раз и более. Этим достигается удлинение времени контакта пищевых частиц с ферментами пищеварения и увеличение всасывающей поверхности. На органном уровне это сопровождается появлением складчатости слизистой оболочки кишки, возникновением ворсинок и крипт. Кишечник ланцетника абсолютно не дифференцирован. В средней его части расположен печеночный вырост — простое слепое выпячивание брюшной стенки кишки, выстланное железистыми клетками. Функционально этот вырост представляет собой зачаточную печень. Он является единственной многоклеточной железой пищеварительной системы ланцетника.
У рыб за глоткой следует короткий пищевод, затем желудок, слабо от него отграниченный. В кишечнике выделяются тонкий и толстый отделы, последний открывается анусом во внешнюю среду. В отличие от ланцетника у рыб хорошо развита печень, снабженная желчным пузырем. Поджелудочная железа у различных рыб построена по-разному. В некоторых случаях она представлена отдельными мелкими дольками в стенке кишечника, в мезентерии, рассеяна в ткани печени. Нередко уже у рыб она представляет собой компактный орган, включающий в себя как экзокринную часть, которая ответственна за синтез пищеварительных ферментов, так и эндокринную, которая выделяет гормоны, регулирующие углеводный обмен. Таким образом, у рыб наблюдаются все основные стадии эволюции сложной многоклеточной железы, структура и функции которой впоследствии принципиально практически не изменяются.
Единственное отличие пищеварительной трубки земноводных от трубки рыб — ее удлинение и впадение толстой кишки в клоаку.
Существенной особенностью пищеварительной трубки пресмыкающихся является возникновение в ней слепой кишки. У большинства из них она зачаточна, но у некоторых развита весьма существенно. Слепая кишка — важное эволюционное приобретение пресмыкающихся, позволяющее им расширить рацион питания и использовать растительную пищу, переваривание которой сложно и требует участия симбиотических простейших и бактерий. Слепая кишка особенно богата микрофлорой, под действием которой осуществляются процессы брожения, позволяющие наиболее полно использовать пластические вещества растительного происхождения.
У млекопитающих за счет дальнейшей дифференцировки пищеварительного тракта, и в частности больших размеров слепой кишки, резко увеличивается разнообразие используемых продуктов питания и возникает пищевая специализация на травоядность и плотоядность; всеядность встречается редко, в частности в отряде приматов.
Задняя кишка плацентарных млекопитающих дифференцирована, клоака отсутствует и прямая кишка заканчивается анусом.
В онтогенезе человека рекапитулируют основные стадии филогенеза кишечной трубки и ее производных. Из пороков развития, связанных с задержкой эмбриогенеза, известны гипоплазии всей пищеварительной системы, укорочение кишки и недоразвитие любых ее отделов, а также печени и поджелудочной железы. В зависимости от степени недоразвития тяжесть аномалии может быть как незначительной, так и несовместимой с жизнью.
Клинический интерес представляет гетеротопия тканей поджелудочной железы в стенке тонкого кишечника или желудка. Иногда гетеротопированные фрагменты могут симулировать опухолевую трансформацию слизистой оболочки. По данным патологоанатомов, частота этой аномалии — от 0,6 до 5%. Механизм гетеротопии — нарушение клеточной миграции зачатков железы из стенки кишечной трубки. С недоразвитием и задержкой дифференцировки связана и такая аномалия, как персистирование клоаки, при которой мочеполовые пути и прямая кишка объединены. При нормальном развитии после 8-й недели эмбриогенеза клоака должна полностью дифференцироваться на прямую кишку, мочевыделительные и половые протоки.
№28
Филогенез выделительной истемы
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом. В эмбриогенезе закладка мочеполовой системы — нефрогонотом — формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целомических животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых.
Эволюция почки
Органами выделения позвоночных являются почки — парные компактные органы, структурная единица которых представлена нефроном. В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос; первичная почка—туловищная, или мезонефрос, и вторичная почка - тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки.
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её.
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле.
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки.
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития.
Эволюция половых желёз
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте, семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
№29
Филогенез дыхательной системы
Дыхание – это совокупность процессов, обеспечивающих потребление организмом кислорода и выделение углекислого газа.
В зависимости от преобладания при дыхании поднятия ребер или уплощения диафрагмы различают типы дыхания: грудной (реберный) и брюшной (диафрагмальный) тип дыхания. Например у мужчин брюшной тип дыхания, а у женщин – грудной.
В процессе дыхания различают 3 этапа: внешнее (лёгочное) дыхание, заключающееся в обмене газов в лёгких между организмом и средой, транспорт газов кровью и тканевое дыхание, состоящее из газообмена в тканях и биологического окисления в митохондриях.
Эволюция органов дыхания
Жабры — наиболее ранние специализированные органы дыхания, появляющиеся среди хордовых впервые у рыб. Они представляют собой тонкие складки слизистой оболочки глотки, лежащие на жаберных дугах, снабжаемые венозной кровью через жаберные артерии и распадающиеся здесь на капилляры.
Позади последней жаберной дуги у кистеперых рыб за счет выпячивания вентральной стороны глотки формируется парное образование — плавательный пузырь, который выполняет в первую очередь гидростатические функции, уравновешивая тело рыбы в толще воды. Связь между ним и глоткой, имеющаяся в эмбриогенезе, не прерывается и позже, поэтому воздух при попадании в глотку может беспрепятственно проходить в плавательный пузырь. Эта морфофизиологи-ческая особенность при изменении условий существования кистеперых рыб явилась предпосылкой преобразования плавательного пузыря в легкие.
У земноводных, связанных по происхождению с кистеперыми рыбами, в личиночном состоянии функционируют жабры, а во взрослом — органами дыхания становятся легкие. В связи с отсутствием грудной клетки и диафрагмы воздух в них попадает из ротовой полости за счет глотательных движений, осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг, следующих за подъязычной дугой, частично редуцируясь, входит в состав хрящей гортани, которая, появляясь впервые у земноводных, является первым органом, относящимся к нижним дыхательным путям. Легкие начинаются непосредственно от гортани. Они крупноячеисты и имеют относительно малую дыхательную поверхность, в связи с чем газообмен в большей степени осуществляется через кожные покровы.
Пресмыкающиеся, вышедшие на сушу окончательно, имеют как верхние дыхательные пути (не полностью отграниченную от ротовой полости полость носа), так и нижние — гортань, трахею и бронхи. Их легкие мелкоячеисты, содержат многочисленные внутренние перегородки и обладают большой дыхательной поверхностью. Впервые появляется диафрагма, которая в дыхании принимает скорее пассивное участие, так как она либо лишена мышечных элементов, либо разделяет грудную и брюшную полости не полностью. Механизм дыхания основан на сокращении межреберных мышц, приводящих в движение грудную клетку.
У млекопитающих дыхательные пути, как и у предков, выстланы мерцательным эпителием. Они полностью отделены от пищеварительной системы и только перекрещиваются с ней в глотке. Бронхи многократно разветвляются, вплоть до бронхиол, ведущих в альвеолы — легочные пузырьки, имеющие в сумме огромную площадь поверхности (у человека до 90 м ). Основной мышцей, изменяющей объем грудной полости, становится диафрагма.
В эмбриогенезе человека отражается первоначальное единство пищеварительной и дыхательной систем. На этом основано формирование большой группы врожденных пороков развития пищевода и трахеи типа эзофаготрахеальных свищей. Нарушением альвеолярной дифференцировки легочной ткани могут быть объяснены дизонтогенетические бронхолегочные кисты — округлые полости в легких, отграниченные от окружающих тканей примитивно построенной стенкой недифференцированного бронха, а также кистозная гипоплазия (недоразвитие) легких . При этой патологии недоразвитыми оказываются целые доли легкого, которые представляют собой многочисленные полости, связанные с крупными бронхами и имеющие малую поверхность. Газообмен в таких участках легких резко ослаблен. Широко известны также гипоплазии диафрагмы от небольших дефектов в ее куполе до полной аплазии. Последняя аномалия несовместима с жизнью и встречается обычно вместе с другими множественными пороками развития.
№30
Филогенез опорно-двигательного аппарата
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов. Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка.
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй — опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся , поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12—13 пар ребер.
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida — дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа.
В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними. Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным.
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих.
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой' и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности — грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными. Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль — стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
В результате из передних отделов складок развились грудные, а из задних — брюшные плавники. Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками. За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7—12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители — стегоцефалы — обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб.
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3—4 ряда, в пястье располагается 7—5 костей, а далее также радиально лежат фаланги 7—5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух — у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду — и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно. Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять.
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3—4-го шейных позвонков, а нижние — на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой — с конечностями, переместившимися на новое место.
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия, или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения — развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1—2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки. Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения — не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят:
изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе;
изменения позвоночного столба — его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении;
изменения черепа — резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка;
специализация верхних (передних) конечностей как органа труда;
появление подбородочного выступа в связи с развитием членораздельной речи.
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
№31
Филогенез нервной системы
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе. Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок. Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря:
передний (I)
средний (II
задний (III).
Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon.
Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon.
Задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека.
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз, или платиневрию,— отсутствие замыкания нервной трубки. Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфогенеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
№32
Филогенез покровов тела
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью. Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других—сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос, покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам. Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна.
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.