

75 Біохімія крові
Кров – рідка сполучна тканина, що бере участь у забезпеченні безперервного зв’язку між органами та системами організму, обміні продуктами життєдіяльності організму з навколишнім середовищем. Кров містить рідку речовину – плазму та формені елементи – клітини крові (еритроцити, лейкоцити і тромбоцити). Кількість крові в організмі людини становить 4,5-5 л (1/13маси тіла).
Кров надходить в усі частини організму та виконує такі важливі функції:
транспортну – перенесення різних речовин між органа-ми і тканинами (кисню, оксиду вуглецю, поживних речовин, медіаторів, ферментів, електролітів, кінцевих продуктів обміну, гормонів тощо. Ці речовини транспортуються у вільному стані або в комплексі з білками;
поживну - кров забезпечує транспорт поживних речовин (вуглеводів, ліпідів, амінокислот та ін.) до тканин;
екскреторну – ця функція тісно пов’язана з транспорт-ною функцією; кров забезпечує виведення з тканин та органів кінцевих продуктів метаболізму (сечовини, сечової кислоти, аміаку тощо);
дихальну – ця функція пов’язана з транспортною функ-цією; кров забезпечує транспорт О2 та СО2 між тканинами та легенями;
регуляторну – кров бере участь у регуляції кислотно-основного стану організму, містить гормони та білки, які беруть участь у процесах координації біохімічних та фізіологічних процесів в організмі;
захисну – кров містить компоненти (лейкоцити, імуногло-буліни, система комплементу), які захищають організм від чужорідних агентів; система коагуляції захищає організм від втрати крові;
75.Гемоглобін: структура, властивості, похідні
Гемоглобін (Hb) – складний білок класу хромопротеїнів, який є гемопротеїном. Гемоглобін має дві основні фізіологічні функції:
1) дихальну – бере участь у транспорті кисню та вуглекислого газу;
2) забезпечує сталість рН (гемоглобінова буферна система є найбільш потужною системою підтримки рН крові).
Нb – це олігомерний білок, який складається з чотирьох субодиниць, або протомерів. Кожна субодиниця містить гем, що зв’язаний з білковою частиною через залишок гістидину. До складу молекули Hb входять по два поліпептидних ланцюги різних видів. Так, основний гемоглобін дорослої людини – HbА1 (96-99% усього гемоглобіну) - містить два α- та два β-ланцюги (α2β2). Також у крові міститься HbА2 (α2δ2), вміст якого стано-вить 2-3%, фетальний гемоглобін HbF (α2γ2), кількість якого – 2-3%. Частина HbА1 глікозильована – це глікозильований гемогло-бін HbА1с, який утворюється в результаті неферментативного глікування гемоглобіну залишками глюкози. Нормальна кон-центрація HbА1с – 4-7%.
У чоловіків концентрація гемоглобіну в нормі становить 130–160 г/л, у жінок – 120-150 г/л. Похідні гемоглобіну. До заліза, яке міститься в молекулі Hb, приєднується кисень – утворюється оксигемоглобін - HbO2 (ва-лентність заліза не змінюється, воно залишається двовалент-ним). У вигляді HbO2 транспортується значна частина кисню. Інтенсивність утворення HbО2 залежить від парціального тиску крові, значення рН, концентрації СО2 та вмісту 2,3-дифосфогліцерату (2,3-ДФГ). Різниця парціального тиску О2 між альвеолярним повітрям та міжклітинною рідиною, куди кисень потрапляє з крові, дорівнює 65 мм рт.ст. Ця значна різниця забезпечує перехід кисню із альвеол у кров і далі - в міжклі-тинну рідину. Крім того, функціонування цитохромоксидази дихального ланцюга призводить до безперервного використання кисню і зниження парціального тиску кисню в мітохондріях до 4 - 5 мм рт.ст. Таким чином, практично створюється «кисневий вакуум» у мітохондріях, що спрямовує потік кисню в клітини. Зв’язування гемоглобіну з різними лігандами, такими, як Н+ (при зниженні рН) та СО2 призводить до конформаційних змін у молекулі гемоглобіну і змінює спорідненість Hb до кисню. У тканинах СО2 витісняє О2 з гемоглобіну, в легенях, навпаки, кисень витісняє СО2 з крові в альвеолярне повітря. Це явище відоме під назвою ефект Бора. Цей ефект також бере участь у регуляції рН крові. У капілярах тканин відбувається приєднання протона до гемоглобіну, і, таким чином, це запобігає закиснен-ню середовища. Крім того, в тканинах збільшення кількості Н+ (при утворенні вугільної кислоти із СО2) знижує спорідненість Hb до кисню. У капілярах легень, навпаки, протон вивільняється, і О2 зв’язується з Hb.2,3-ДФГ – метаболіт, який утворюється з 1,3-ДФГ (про-міжний продукт гліколізу) та знижує спорідненість гемоглобіну до кисню, що сприяє вивільненню О2 у тканинах. Гемоглобін, який віддає кисень, має назву дезоксигемоглобін, або відновлений гемоглобін (НHb). Приєднання вуглекислого газу призводить до утворення карбгемоглобіну HbCO2 (СО2 з’єднується з N-кінцевими групами гемоглобіну). У складі цього похідного транспортується до 20% СО2. Молекула гемоглобіну може утворювати комплекси з іншими газами. Так, комплекс гемоглобіну з чадним газом – карбоксигемоглобін (HbCO) є міцною сполукою. Спорідненість Hb до СО у 200 разів вище, ніж до кисню, тому утворення кар-боксигемоглобіну блокує утворення оксигемоглобіну і транс-порт кисню. Саме тому навіть незначні кількості чадного газу в повітрі є небезпечними для життя. У крові людини, яка живе у місті, концентрація карбоксигемоглобіну становить менше ніж 2%. У крові людей, які палять, ця концентрація зростає до 10%. При деяких патологічних станах, наприклад при отруєн-нях потужними окисниками (перманганат калію, бертолетова сіль, сульфаніламідні препарати та ін.), залізо у складі гему окиснюється до тривалентного стану – утворюється метгемо-глобін MetHb. Цей похідний гемоглобіну не може зв’язувати кисень. У нормі в еритроцитах міститься до 2% метгемоглобіну, який утворюється в результаті аутоокиснення. Така незначна кількість не пригнічує газообміну. Накопиченню метгемогло-біну перешкоджає функціонування ферменту метгемоглобінре-дуктази, яка відновлює MetHb. Метгемоглобінемія (підвищення концентрації MetHb) може мати спадковий характер (при дефіциті метгемоглобін ре-дуктази) та розвиватися внаслідок надходження в організм значної кількості окисників – нітритів, аніліну, нітробензолу та ін. (розвивається гостра токсична метгемоглобінемія).При порушенні синтезу гемоглобіну виникають гемоглобінопатії і таласемії, які мають спадковий характер і належать до «молекулярних хвороб». Гемоглобінопатії є наслідком зміни кількісного або якісно-го амінокислотного складу поліпептидних ланцюгів гемогло-біну, тому вони належать до якісних гемоглобінопатій. Таласемії зумовлені порушенням швидкості синтезу поліпептидних ланцюгів гемоглобіну без зміни їх структури, тому вони ще мають назву кількісні гемоглобінопатії. Деколи спостерігається наявність цих двох патологій у одного хворого. Для гемоглобінопатій і таласемій характерний синдром еритропатій, який супроводжується: скороченням тривалості життя еритроцитів, підвищеним гемолізом, порушенням функцій еритроцитів. Відомо близько 300 аномальних гемогло-бінів, але не всі патології мають клінічні прояви. Перші аномальні гемоглобіни називали за літерами латинського алфавіту, але, оскільки існує велика кількість патологічних форм, до назв цих гемоглобінів почали включати назви місць їх відкриття (Москва, Boston) або назви шпиталів.Найчастіше при гемоглобінопатіях спостерігаються гемогло-біни S, C, D, E, H.HbS – гемоглобін, в якому в 6-му положенні β-ланцюга глутамінова кислота (Глу) замінена на валін (Вал). Валін має неполярний радикал, тому така заміна призводить до зниження розчинності гемоглобіну. Внаслідок синтезу HbS змінюється структура еритроцитів – кристалізація гемоглобіну у вигляді тектоїдів супроводжується розтягненням оболонки і вони наби-рають форми серпа. Тому ця патологія має назву серпоподібно-клітинна анемія. У результаті спостерігаються підвищення в’язкості крові, зменшення швидкості кровотоку, зниження механічної резистентності еритроцитів - вони втрачають здат-ність проходити через дрібні капіляри. Такі еритроцити застря-ють у капілярах, руйнуються і утворюють тромби, наслідком чого є хронічна капіляропатія. HbC – гемоглобін, в якому в 6-му положенні β-ланцюга глу-тамінова кислота (Глу) замінена на лізин (Ліз). Цей гемоглобін також кристалізується в еритроцитах, які гемолізують, резуль-татом чого є розвиток анемії. Високий вміст HbC призводить до розвитку легкої форми гемолітичної анемії.HbD – гемоглобін, в 121-му положенні бета-ланцюга якого глутамінова кислота (Глу) замінена на глутамін (Глн). При висо-кому вмісті такого аномального гемоглобіну розвивається легка форма гемолітичної анемії.HbЕ – гемоглобін, в 26-му положенні β-ланцюга якого глу-тамінова кислота (Глу) замінена на лізин (Ліз). Внаслідок дефі-циту нормальних β-ланцюгів за симптоматикою ця гемогло-бінопатія подібна до β-таласемії і супроводжується розвитком мікроцитарної гіпохромної анемії та наявністю мішенеподібних еритроцитів. Бувають також більш тяжкі форми цієї патології, які супроводжуються вираженою спленомегалією.HbM – існує група гемоглобінів, у яких структурний дефект (амінокислотна заміна) перешкоджає відновленню метгемогло-біна до гемоглобіну. У таких гемоглобінах метгемоглобінредук-таза не може відновити тривалентне залізо до двовалентного стану, тому в еритроцитах спостерігається накопичення метгемоглобіну.Таласемії (греч. thalassa - море і гемо...) – це найбільш поширені спадкові захворювання людини. Залежно від того, порушення синтезу яких ланцюгів гемоглобіну спостерігається при цій патології, таласемії прийнято поділяти на дві групи: α- та β-таласемії. Альфа-таласемія обумовлена порушенням синтезу α-ланцюгів Hb (внаслідок делеції або інактивації одного з чотирьох генів альфа-ланцюгів глобіну). Бета-таласемія розвивається внаслідок порушення синтезу β-ланцюгів.При альфа-таласемії в організмі дорослої людини форму-ються бета4-тетрамери – це гемоглобін Н (HbH). Ці тетрамери нестабільні, крива дисоціації оксигемоглобіну не має S-подібної форми. У хворих спостерігається мікроцитарна гіпохромна анемія. Гемолітичні прояви захворювання обумовлені кількістю таких тетрамерів.При бета-таласемії надлишок α-ланцюгів гемоглобіну не утворює тетрамерів, α-ланцюги Нb зв’язуються з мембранами еритроцитів та пошкоджують їх. Гомо- та гетерозиготи мають різні за тяжкістю клінічні прояви. Варіації симптомів можуть бути від тяжкої анемії з клінічною симптоматикою, яка не дозволяє людині жити довше ніж до 20 років (анемія Кулі), до легкої мікроцитарної анемії.
76 Буферні системи крові.
У крові людини існують спеціальні системи, які сприяють підтримці сталості рН крові, - буферні системи. Вважають, що, завдяки існуванню саме буферних систем, рН крові – це найбільш стабільна константа внутрішнього середовища. У нормі відхилення від середнього значення 7,4 становить не більше ніж 0,04. Як правило, буферна система – це суміш сполук, які відіграють роль донора і акцептора протонів. Вивільнення та поглинання протонів із середовища сприяє стабілізації нормальних значень рН рідин організму (крові і тканин) в разі надходження кислот або лугів. Буферний розчин містить слабку кислоту та її сіль, яка утворена сильною основою (спряжена кислотно-основна пара). Така суміш має певну та відносно сталу концентрацію іонів водню, яка залежить від співвідношення кислоти та солі й іонної сили суміші. У крові функціонують такі буферні системи (в дужечках зазначена кислотно-основна пара кожної системи):
- гемоглобінова (відновлений Hb – калійна сіль Hb: ННb-КНbО2);
- гідрокарбонатна або бікарбонатна (вугільна кислота – двовуглекислий натрій: Н2СО3 — NаНСО3);
- фосфатна (одноосновний –двоосновний фосфорнокислий натрій: NaH2PO4- Na2HPO4);
- білкова.
Кожна з них має певну буферну ємність та їх дія взаємозв’язана.
Найбільш потужною буферною системою крові є гемоглобі-нова. Її буферна ємність – 73-76%. Відповідно для бікарбонатної – 17-27%, білкової – 2-5%, фосфатної – 1-2%. Головною фізико-хімічною системою регуляції плазми є бікарбонатна буферна система. Крім того, гемоглобінова, білкова та фосфатна буферні системи регулюють рН крові саме через стабілізацію цього буфера.Гемоглобінова буферна система Ця система представлена декількома підсистемами – HHb/HНbО2, KНb/HНb, KНbО2/HНbО2. Оксигемоглобін НbО2, який в крові найчастіше буває у вигляді калієвої солі, є більш сильною кислотою, ніж HHb. В еритроцитах периферичних тканин КНbО2 віддає кисень, який дифундує в тканини, та іон К+. Після цього Нb приєднує Н+, який вивільняється при дисоціації вугільної кислоти і перетворюється до ННb – більш слабкої кислоти. Іони калію з’єднуються з гідрокарбонатом, який утворився з СО2. Послідовність реакцій виглядає так:
СО2 + Н2О ↔ Н2СО3;
Н2СО3 ↔ Н+ + НСО3-;
НСО3- + К+ ↔ КНСО3;
КНb + Н2СО3 → КНСО3 + ННb.
У венозній крові, яка спрямовується до легень, містяться еритроцити з відновленим гемоглобіном. У легенях із ННb утворюється HНbО2. Крім того, в тканині легень внаслідок виділення великої кількості СО2 спостерігається надлишок аніонів НСО3-, тому є загроза зміщення рН у лужний бік. Це не відбувається тому, що аніони НСО3- транспортуються в еритроцити та перетворюються в основу КНСО3-. При взаємодії КНСО3- з HНbО2 формується KНbО2 та Н2СО3. Далі, вугільна кислота під дією карбоангідрази дисоціює на Н2О та СО2. Вуглекислота виділяється через легені. Таким чином, функціонування гемоглобінової буферної системи пов’язано з основною функцією гемоглобіну, а саме з транспортом кисню. Важливо те, що гемоглобінова буферна система взаємодіє з бікарбонатною системою, яка є основним лужним резервом крові. У тканинних капілярах взаємодія гемоглобіну з вуглецевою кислотою призводить до збереження гідрокарбонатів, тобто лужних резервів крові.
У легенях відбувається оксигенація гемоглобіну, що супроводжується одночасним від’єднанням СО2 від КНСО3, тому що оксигемоглобін є більш сильною кислотою. Іони К+ приєднуються до НbО2, в результаті чого утворюється калієва сіль оксигемоглобіну. Гідрокарбонат (НСО3-) приєднує протон, внаслідок чого утворюється вугільна кислота. Карбоангідраза розщеплює Н2СО3 з утворенням СО2, який переходить з еритроцитів у плазму, далі - в альвеоли легень та виділяється з організму. Виділення СО2 з організму запобігає зниженню рН крові. Приєднання кисню до Нb у легенях, віддача кисню в тканинах, проникнення всередину еритроцитів СО2 – це досить складний процес, у якому беруть участь аніони хлору, катіони калію та інші іони. Ступінь зв’язування гемоглобіну з киснем залежить від рН крові. Відомо, що при зниженні рН спорідненість Hb до кисню знижується і, тим самим, зменшується насиченість гемоглобіну киснем. З іншого боку, коли рН крові збільшується, спостерігається протилежний процес – спорідненість гемоглобіну до кисню та його насиченість киснем зростають. Описана закономірність має назву ефект Бора. Бікарбонатна (гідрокарбонатна) буферна система Ця система за своєю потужністю займає друге місце в сис-темі регуляції рН крові. Вона складається з вугільної кислоти Н2СО3 та бікарбонату натрію NaHCO3 (переважно в плазмі) або калію KHCO3 (в еритроцитах). Співвідношення вмісту кисло-ти та її солі становить 1/20.
При надходженні у кров кислоти, яка більш сильна, ніж вугільна, в реакцію вступає бікарбонат. Утворюється нейтральна сіль та слабкодисоційована вугільна кислота. Вугільна кислота під дією карбоангідрази розщеплюється до Н2О та СО2, останній виділяється легенями:
1) Na+ + HCO3- + H+ + X-→ NaX + H2CO3
(X – аніон кислоти);
2) H2CO3→ H2O + CO2.
У разі, якщо в кров надходить луг, в реакцію вступає вугільна кислота і утворюються гідрокарбонат і вода:
НСО3 +Na+ →NaHCO3.
Надлишок бікарбонату виділяється нирками. Фосфатна буферна система
Вищезазначена буферна система складається з дигідрофос-фату (NаН2РО4) та гідрофосфату (Nа2НРО4) натрію. Перший з них має властивості слабкої кислоти та взаємодіє з лужними речовинами, які надходять у кров. Другий компонент має влас-тивості слабкого лугу та реагує з більш сильними кислотами. Ця буферна система здатна стримувати зміни рН в інтервалі 6,2 – 8,2, таким чином, може забезпечити значну частку буферної ємності крові. На жаль ємність цього буфера невелика у зв’язку з низьким вмістом фосфатів у крові. При підвищенні кислот у крові відбувається така реакція:
2Na+ + HPO42- + H+ + X-→NaX + Na+ + H2PO4-.
Надлишок гідрофосфату натрію виділяється із сечею. При підвищенні кількості лугів процес, що відбувається, ілюструє така реакція:
Na+ + H2PO4- + Na+ + OH- → H2O + 2Na+ + HPO42- .
Між неорганічними буферними системами існують певний зв’язок та динамічна рівновага. Так, наприклад, фосфатна буферна система може підвищувати резерви бікарбонатної за рахунок відновлення гідрокарбонату натрію:
2Na+ + HPO42- + H2CO3 ↔ NaHCO3 +Na+ + H2PO4- .
У результаті ліквідується надлишок H2CO3 в плазмі та відновлюється постійне співвідношення концентрацій вугільної кислоти та бікарбонату натрію (1/20).
Білкова (протеїнова) буферна система
Ця система здійснює підтримку рН за рахунок амфотерних властивостей білків плазми, які містять вільні -СООН та -NH2. У лужних умовах вони поводять себе як кислоти (донори протонів), а в кислих - надають лужних властивостей білкам, які поводять себе як акцептори протонів.
Вище було зазначено, що нормальне значення такого показника, як рН крові, становить 7,4 ± 0,04 (або 7,36-7,44). Було встановлено, що це значення може бути суттєво змінено без змін метаболізму, які б були несумісні з життям. Лише при значеннях рН менше ніж 6,8 та більше ніж 8 в організмі відбуваються незворотні зміни обміну речовин.
Для характеристики регуляторних фізико-хімічних, фізіологічних та інших механізмів, які забезпечують сталість рН введений термін «кислотно-основний стан» організму (КОС). У регуляції КОС, крім буферних систем організму, беруть участь легені, нирки, печінка та кишечник. Через легені, як ми з’ясували, видаляються леткі кислоти, насамперед вугільна. Із сечею видаляються нелеткі кислоти (кислі фосфати) та сполуки лужного характеру (гідрокарбонати, іони амонію та ін.). Роль печінки у процесі підтримки сталості КОС організму полягає у тому, що в цьому органі відбуваються процеси повного катаболізму недоокиснених продуктів, що утворюються при метаболізмі. У разі патологічних змін у паренхімі печінки у кров надходять речовини, які здатні зменшувати значення рН. Участь ШКТ у регуляції КОС активно проявляється при втраті іонів водню, хлору, калію при блюванні та бікарбонатів - при діареї. Для характеристики КОС організму використовують мікро-метод Аструпа, що базується на принципі Зигарда-Андерсена. В основу методу (згідно з рівнянням Гендерсона-Гессельбаха) покладена залежність між рН та рСО2. Метод полягає у визначенні рН плазми крові до і після еквілібризації проби крові двома газовими сумішами, що містять 4% та 8% СО2. Дані, які отримують у ході експерименту, використовують для визна-чення рСО2, SB (СБ – стандартний бікарбонат) та ВЕ (base excess або надлишок основ) за стандартною нанограмою. Нанограма являє собою залежність між рН та рСО2, тобто є графічним зображенням буферної лінії крові.Оцінку КОС можна зробити, як мінімум, за трьома основними показниками рН, рСО2 та ВЕ (або SB): рН – негативний десятковий логарифм концентрації іонів водню; рСО2 – парціальний тиск СО2 у плазмі, мм Hg; SB (стандартний бікарбонат) – вміст іона бікарбонату в плаз-мі крові, яка повністю насичена киснем та приведена до стан-дартних умов. Це основний показник, який безпосередньо відображає нирковий компонент регуляції КОС; ВЕ (надлишок основ, Basis excess) – відображає кількість невикористаних буферів крові, головним чином бікарбонатного та гемоглобінового. Це різниця між фактичною та необхідною величинами суми всіх аніонів нерозбавленої крові (ВВ – buffer base) при метаболічному алкалозі. Крім того, для характеристики КОС використовують такі показники: ВВ (buffer base) – сума всіх аніонів нерозбавленої крові (фактично, SB + аніонний запас білкового буфера). BD (base deficit) – різниця між фактичним та необхідним значенням ВВ при метаболічному ацидозі. АВ (актуальний бікарбонат крові) – концентрація гідрокарбонатів (НСО3-) при фізіологічних конкретних умовах. Нижче наведені нормальні значення основних показників КОС організму:Ацидоз та алкалоз – порушення КОС організму, коли регуляторні системи (у тому числі й буферні) не можуть підтримувати сталість рН на постійному рівні, і саме тому в крові накопичується значна кількість кислот або лугів. При ацидозі в крові з’являється відносний або абсолютний надлишок кислот, при цьому спостерігається зниження рН крові (рН<7,37). Алкалоз характеризується підвищенням вмісту лугів у крові і підвищенням рН (рН>7,43). Кожний з цих двох типів змін КОС поділяють на декілька видів залежно від причин порушення сталості рН.
Виділяють дихальний (респіраторний, газовий) та метаболіч-ний (негазовий, недихальний, нереспіраторний) ацидоз/алкалоз.Нижче наведена класифікація порушень КОС та причини, які їх викликають.
Ацидоз:Газовий,Негазовий:
а) метаболічний: кетоацидоз, лактоацидоз, ацидоз при накопиченні органічних та неорганічних кислот (при запаленнях, травмах);
б) видільний: затримка кислот (дифузний нефрит, уремія), втрата лугів (нирковий);
в) екзогенний: довготривале вживання кислої їжі, приймання ліків, отруєння кислотами;
г) комбіновані форми.
3. Мішаний (1+2): при асфіксії, серцево-судинній недостат-ності.
Алкалоз:1Газовий:а) гіпервентиляція;б) підсилене виділення СО2 як наслідок інших причин.2. Негазовий:а) видільний: затримка лугів, втрата кислот, гіпохлоремічний (метаболічний);б) екзогенний: довготривале вживання лужної їжі, введення ліків (бікарбонатів та інших лужних сполук).3. Мішані форми (1 + 2).Негазовий (метаболічний) ацидоз є найбільш поширеною формою порушення КОС
Стани, які призводять до виникнення негазового ацидозу.Підвищення введення іонів Н+ (екзогенний ацидоз) – введення може бути у вигляді кислот або деяких кислих солей (СаСl2, NH4Cl).Підвищення утворення Н+ (власне «метаболічний» ацидоз) – утворення великої кількості кислих продуктів метаболізму (кетонових тіл, лактату, інших органічних кислот). Кетоацидоз виникає у хворих на цукровий діабет, тиреотоксикоз, при лікувальному голодуванні тощо. Лактоацидоз є результатом тканинної гіпоксії при недостатній функції сердцево-судинної системи, ефірній анестезії, шоку, ускладнених пневмоніях тощо. Мішані форми метаболічного ацидозу виникають при сепсисі, опіках, гострій печінковій недостатності, рахіті (у дітей) тощо.Зниження виділення іонів Н+ із сечею – це спостерігається при олігурії або анурії, нирковій недостатності (яка супроводжується накопиченням залишків сірчаної та фосфорної кислот), захворюванні нирок зі зниженою клубочковою фільтрацією.Підвищення втрати бікарбонатів через кишечник (при діареї), із сечею (видільний ацидоз) – при тубулярному нефриті, під впливом інгібіторів карбоангідрази (діуретиків діакарбу, діамоксу), які порушують процеси «реабсорбції» бікарбонату. Газовий (респіраторний) ацидоз - цей вид порушення КОС виникає як результат накопичення вуглекислого газу в організмі (гіповентиляція легень, набряки легень, порушення механіки дихання тощо).Негазовий (метаболічний) алкалоз – розвивається як наслідок введення великих доз гідрокарбонату натрію (при корекції метаболічного ацидозу), втрати великої кількості кислот та хлоридів (багаторазовому блюванні), некоректній гормональній терапії, яка не враховує втрати калію (при синдромі Іценко-Кушинга, Конна), тощо.
Газовий (респіраторний) алкалоз – є результатом гіпервен-тиляції легень, виникає при гематологічних та серцево-судинних захворюваннях, стимуляції дихального центру при органічних ураженнях ЦНС, сепсисі, невротичній задусі, яка викликана стресом, біллю, сильним плачем у дітей тощо.
771.1. Альбуміни
Частка альбумінів – 40-50 г/л ( 52-65% загального білка).
Вони зв’язують вільні жирні кислоти і транспортують їх. Таке зв’язування забезпечує зниження концентрації фізіологічно активних вільних жирних кислот у 10 000 разів. Тому зниження кількості альбумінів може бути однією з причин розвитку жирової інфільтрації печінки.
Альбуміни також мають різні специфічні центри для зв’язування з гормонами (тиреоїдними, стероїдними, інсуліном). Встановлено, що насичення альбумінів одним гормоном не пригнічує зв’язування з іншим. Кількість альбумінів регулює вміст вільних гормонів, які є активними. Таким чином, регулюється ступінь активності деяких гормонів. Зниження кількості альбумінів призводить до серйозних метаболічних та фізіологічних розладів, пов’язаних із зростанням гормональної активності;,мають буферні властивості, які зумовлені наявністю вільних аміно- та карбоксильних груп у структурі білка;,можуть виконувати резервну та пластичну функції. Встановлено, що при аліментарній недостатності білка альбуміни можуть бути використані тканинами як пластичний матеріал для побудови власних білків. Глобуліни
Це гетерогенна суміш білкових молекул, в якій виділяють α-, β- та γ- глобуліни. Кожна з цих фракцій містить специфічні білки, які виконують певні біохімічні функціїДо α1-глобулінів належать α1-антитрипсин, α1-кислий гліко-протеїн, альфа-фетопротеїн та інші.α1-Антитрипсин (2-2,5 г/л) – це глікопротеїн (природний інгібітор протеїназ). Його біологічна функція реалізується шляхом зв’язування з ферментами трипсином, хімотрипсином, плазміном, тромбіном, що призводить до пригнічення активн-ості цих протеїназ. Антитрипсин належить до білків гострої фази. У гострій фазі захворювання в печінці спостерігається підвищення синтезу цього білкаСпадкові порушення синтезу α1-антитрипсину призводять до розвитку емфіземи легень у людей віком 20-40 років та неонатального гепатиту, наслідком якого може бути цироз печінки. Причиною емфіземи є відсутність механізмів інгібу-вання еластази (також протеїнази), яка бере участь у деструктив-них процесах у легенях.
α1-Кислий глікопротеїн (орозомукоїд, AGP - a1-acid glycopro-tein) – високомолекулярний білок, фізіологічна роль якого до кінця ще не встановлена, але існує точка зору про наявність у нього імуномодуляторних властивостей – AGP може зв’язувати-ся з ендогенними та екзогенними медіаторами запалення. Орозомукоїд може захищати організм в умовах підвищеної продукції цитокінів запалення (наприклад, при ендотоксичному шоці). У період розвитку запальної реакції концентрація цього білка зростає у 2- 4 рази, тому AGP також належить до білків гострої фази. Альфа-фетопротеїн (АФП, a-Fetoprotein) – глікопротеїн сироватки крові, який починає вироблятися на 5-му тижні розвитку плода. За будовою АФП подібний до альбумінів і виконує аналогічну функцію в організмі плода. В акушерстві та гінекології цей білок є одним із показників стану розвитку плода та наявності спадкової патології. Визначення концентрації АФП у крові матері разом з рівнем β-хоріонічного гонадотропіну (ХГЧ) та вільного естрадіолу належить до «потрійного тесту» діагностики риску відхилень у розвитку плода. В онкології АФП використовують як один з ембріональних імунологічних маркерів злоякісних пухлин (онкофетальний антиген). У дорослих цей білок визначається в крові у високих концентраціях у разі доброякісних або злоякісних проліфера-тивних процесів у клітинах, в яких він виробляється в ембріо-нальному періоді. Крім того, визначення концентрації АФП призначають для виявлення метастазування пухлини в печінку, оцінки терапії злоякісних пухлин, скринінгу груп .До α2-глобулінів належать гаптоглобін, α2-макроглобулін, церулоплазмін, С-реактивний білок. Гаптоглобін (Нр) – глікопротеїн, біологічною функцією якого є зв’язування вільного гемоглобіну (Hb), що вивільняється при внутрішньосудинному гемолізі. Таким чином, Нр перешкоджає втраті організмом гемоглобіну тому, що комплекс Нр-Hb не може пройти крізь ниркові клубочки. Комплекс Нр-Hb за декіль-ка хвилин видаляється клітинами РЕС. Залізо, яке вивільняється, надходить у кров, зв’язується трансферином та транспортується до клітин кісткового мозку для синтезу Hb.
Цей білок синтезується в печінці і належить до білків гострої фази. Його кількість зростає і є наслідком стимуляції синтезу інтерлейкінами при всіх ексудативно-запальних процесах. Вміст Нр свідчить про стан сполучної тканини – кількість цього білка зростає при деструктивних змінах у тканині та знижується при терапії глюкокортикоїдами. Збільшення концентрації гаптогло-біну можливе при нефротичному синдромі, пухлинах, холестазі, колагенозах, лімфогранулематозі тощо. Зниження концентрації цього білка спостерігається при гострих і хронічних захворю-ваннях печінки, при усіх видах гемолізу, дефіциті глюкозо-6-фосфатдегідрогенази тощо. α2-Макроглобулін (АМГ, α2-сіромукоїд) – це глікопротеїн, білок гострої фази з великою молекулярною масою, який синтезується в підшлунковій залозі, є інгібітором протеаз (як і антитрипсин, але з більш широким спектром активності). Він інактивує плазмін, а також знижує активність тромбіну. Встановлено, що АМГ транспортує цитокіни (інтерлейкіни, інтерферони, стимуліни, фактори некрозу пухлин, інгібіни, фактори росту). Крім того, він має антиоксидантні властивості, зв’язує інсулін, у дітей підвищується при нефротичному синдромі. Церулоплазмін – мідьвмісний білок, який містить по 8 іонів Сu+ та Cu2+. Він є головним мідьвмісним металопротеїном крові (містить 95% міді сироватки крові та 3% міді організму). Цей білок виконує декілька функцій. По-перше, церулоплазмін зв’язує і забезпечує транспорт міді у крові. По-друге, він має ферментативні властивості – є оксидазою аскорбінової кис-лоти, адреналіну, норадреналіну, ДОФА, серотоніну, інактивує активні форми кисню (антиоксидантна властивість). Також він може каталізувати окиснення Fe2+ до Fe3+, саме тому його ще називають фероксидазою. Недостатність церулоплазміну є результатом пригнічення його синтезу в печінці та спостерігається при хворобі Вільсона-Коновалова або тяжких ураженнях печінки. Дефіцит цього білка може бути також наслідком його втрати організмом (нефротичний синдром).С-Реактивний білок (CRP) – отримав назву завдяки здатності вступати в реакцію преципітації з С-полісахаридом пневмококів (цей важливий механізм раннього захисту організму від інфекції). Цей білок належить до білків гострої фази, є чутливим індикатором ураження тканин при запаленні, некрозі, травмі.
CRP синтезується переважно у печінці. Швидке підвищення концентрації С-реактивного білка (в сотні разів) спостерігається при запальних процесах різної етіології та локалізації, інфекційних захворюваннях, травмах, пухлинах, які супровод-жуються некрозом та запаленням. Після інфаркту міокарда зростання кількості цього білка спостерігається вже на 2-й день захворювання (при стенокардії збільшення вмісту CRP не спостерігається). С-реактивний білок також виявляється в крові у гострій фазі ревматизму, при колагенозах, захворюваннях на рак, бактеріальній та вірусній інфекції. β-Глобуліни
Трансферин (Tf) – це глікопротеїн плазми, який синтезується в печінці і є основним транспортером іонів заліза. Синтез Tf в печінці залежить від функціонального стану органа, потреб та резервів заліза в організмі. У нормі приблизно 30% цього білка зв’язано із залізом; кожна молекула білка містить 2 атоми металу. 1 г трансферину зв’язує 1,25 мг заліза. Трансферин має велику спорідненість до заліза, тому теоретично в 1 літрі крові міститься лише 1 атом вільного заліза. Вміст Tf у жінок на 10% вищий ніж у чоловіків. Концентрація його знижується у людей похилого віку. При дефіциті заліза в разі приймання естрогенів концентрація трансферину у крові також зростаєГемопексин (Р-глобулін) – білок, який зв’язує вільний гем гемопротеїнів (гемоглобіну, міоглобіну, каталази) та транспотує його в клітини РЕС печінки для реутилізації. Це перешкод-жає виведенню гему нирками і втраті заліза із сечею. β2-Мікроглобулін (beta2-МГ) – низькомолекулярний білок поверхневих антигенів клітинних ядер. Це компонент білків першого класу головного комплексу гістосумісності. Вeta2-МГ має антиоксидантні властивості. У крові його концентрація 2,4 мг/л, із сечею виділяється приблизно 130 мкг/л. Підвищення вмісту цього білка спостерігається при злоякісних захворюваннях, ревматоїдному артриті, інфаркті міокарда, опіках, аутоімунних захворюваннях, порушеннях клітинного імунітету (наприклад, СНІД), трансплантації органів тощо. У сечі концентрація beta2-МГ підвищується при діабетичній нефропатії, інтоксикації важкими металами (наприклад, кадмієм). γ-Глобуліни Ця фракція містить основну масу антитіл (імуноглобулінів), що забезпечують гуморальну захисну реакцію організму1.3. Деякі «індикаторні» білки крові
Тропонін Т належить до скорочувальних білків м’язів. У крові навіть при важкому фізичному навантаженні не спостерігається суттєвих змін його концентрації. Тому тропонін Т розглядають виключно як
кардіоспецифічний маркер. При інфаркті міокарда зростання концентрації цього білка спостерігається через 3-4 години після больового нападу. Максимальне зростання концентрації - через 3-4 доби. Специфічність методів визначення концентрації тропоніну Т – 901-100%, що значно вище, ніж визначення інших маркерів, таких, як креатинфосфокіназа (КФК), лактатдегідрогеназа (ЛДГ), міоглобін. Крім того, концентрація тропоніну Т після початку інфаркту міокарда зростає значно більше, ніж активність названих ферментів (КФК та ЛДГ). Також зростання концентрації тропоніну Т спостерігається при гострій алкогольній інтоксикації (але не хронічній).
78 Ферменти крові
Ферменти сироватки (плазми) крові умовно поділяють на три групи:
індикаторні (клітинні, маркерні) ферменти – локалізо-вані в клітинах тканин, потрапляють у кров у результаті фізіологічного старіння та руйнації клітин або в результаті підвищення проникливості клітинних мембран. У крові міс-титься декілька десятків індикаторних ферментів. У нормі клітинні ферменти у крові мають невелику активність та не виконують специфічних функцій. При надходженні у кров вони інактивуються протеазами сироватки і тканин. Актив-ність цих ферментів зростає при ураженні органів, коли спостерігається потужна руйнація клітинних мембран. Фер-менти цієї групи поділяють на неспецифічні та органо-специфічні.
Неспецифічні індикаторні ферменти каталізують уні-версальні реакції метаболізму та локалізовані в більшості органів і тканин. Органоспецифічні ферменти містяться ли-ше в тих органах і тканинах, де відбуваються специфічні реакції, властиві лише для клітин цього органа. Саме тому підвищення активності цих ферментів у крові свідчить про органну локалізацію патологічного процесу; секреторні (плазмоспецифічні) ферменти – синтезуються в печінці, виділяються у кров, де виконують певні фізіологічні функції (ферменти системи згортання крові, фібринолізу, холінестераза, церулоплазмін, протеази ренін-ангіотензинової та калікреїнової систем тощо);екскреторні ферменти – синтезуються в печінці, підшлунковій залозі, слизовій оболонці кишечника. Поява цих ферментів у крові пов’язана з природною руйнацією клітинних структур, у яких вони утворюються (лужна фосфатаза, лейцинамінопептидаза, ентерокіназа, ГГТП, трипсин, ліпаза та ін.). Виділяють декілька типів зміни активності ферментів у крові: гіперферментемія, гіпоферментемія та дисферментемія.
Гіперферментемія – підвищення активності ферментів у сироватці крові. Це може бути наслідком: 1) надходження ферментів у кров з пошкоджених органів і тканин; 2) підвищен-ня каталітичної активності ферментів як безпосередньо у пошкодженому органі, так і при надходженні в кров’яне русло. Гіпоферментемія – зниження активності ферментів крові, що є результатом пригнічення синтезу ферментів у тканинах. Цей вид зміни активності ферментів характерний лише для окремих ферментів, наприклад, для холінестерази. Дисферментемія – цей тип характеризує появу деяких ферментів у крові, активність яких у нормі відсутня. Такі зміни можуть бути характерні для деяких органоспецифічних ферментів, наприклад, сорбітолдегідрогенази, фруктозомоно-фосфатальдолази та ін.Креатинфосфокіназа (КФК, креатинкіназа – КК) у крові людини може бути наявна в різних формах: КФК-ММ, КФК-МВ, КФК-ВВ. ММ - це м’язовий ізофермент, ВВ - мозковий, МВ – має високу активність у міокарді. Крім того, в сироватці крові мястяться макро-КФК типу 1 (зв’язані з імуноглобуліном), макро-КФК типу 2 (мітохондріальна форма).
Оскільки ізоферменти КФК мають велику активність переважно в скелетних м’язах, міокарді і ЦНС, визначення загальної активності КФК та ізоферментів має значення для діагностики захворювань ЦНС, міопатій та інфаркту міокарда.Зростання активності КФК при інфаркті міокарда починається через 2 – 3 години після початку нападу, максимум спостерігається через 13 – 30 годин (у 5 – 10 разів перевищує норму). Особливе значення має визначення активності МВ-КФК. Якщо активність МВ-КФК становить більше ніж 5% від загальної активності КФК, ураження міокарда можна вважати доведеним. У гострому періоді інфаркту міокарда співвідно-шення МВ-КФК/загальна КФК підвищується від 3 до 40 %. На жаль, у нашій країні визначення ізоферментного спектра можливе не в усіх клініко-діагностичних лабораторіях.Крім того, високе діагностичне значення має визначення співвідношення активностей КФК/АсАТ, яке використовується для диференціальної діагностики інфаркту міокарда та уражень скелетних м’язів. При ураженні скелетних м’язів це співвідношення дорівнює приблизно 27 (13-56); про патологію кардіоміоцитів свідчить значення близько 5 (2 – 9).
Визначення активності амінотрансфераз АлАТ і АсАТ в сироватці крові має важливе значення також для діагностики захворювань печінки. Значне підвищення активності цих ферментів спостерігається при гострому вірусному гепатиті. Встановлено, що при цій патології домінує активність АлАТ. При внутрішньопечінковому холестазі, цирозі печінки, метаста-зах карциноми активність АсАТ у крові підвищена і більша, ніж АлАТ.При захворюваннях печінки у крові також зростає актив-ність глутаматдегідрогенази, органоспецифічних ферментів - цистази, урокінази, сорбітолдегідрогенази, орнітинкарбомоїл-трансферази, глутаматдегідрогенази. Підвищення активності лужної фосфатази спостерігається при двох видах патологій: захворюваннях кісток, які супроводжуються проліферацією остеобластів та хворобах з явищами холестазу. Крім того, визначення активності лужної фосфатази має велике значення для діагностики рахіту у дітей. Підвищення активності кислої фосфатази спостерігається при новоутвореннях у передміхуровій залозі, тромбоемболіях, гемолітичних станах, ураженнях кісток метастазами тощо. У крові людини також міститься альфа-амілаза, яка має різне походження, тобто двох типів: панкреатична (Р) та сли- ни (S). У практично здорової людини активність цих двох ізоен-зимних типів приблизно однакова. Вищеназваний фермент зв’язаний з білками плазми крові. Активність альфа-амілази значно зростає при захворюваннях підшлункової залози (гострому панкреатиті); незначне підвищення активності спосте-рігається при холециститі, захворюваннях нирок, простатиті, черепно-мозкових травмах, пневмонії, діабетичному кето-ацидозі, в післяопераційний період тощо.
79 Калікреїн-кінінова система
У крові містяться низькомолекулярні пептиди – кініни. Вони беруть участь у регуляції тонусу судин (розширюють їх), знижують кров’яний тиск, регулюють процеси мікроциркуляції, беруть участь у запальних та алергічних реакціях. Найважливішими кінінами є брадикінін та калідин. Ці пептиди синтезуються з неактивних попередників, що постійно наявні в плазмі крові та міжтканинній рідині. Неактивні їх попередники – кініногени – білки, які синтезуються в печінці та зв’язуються в крові з α2 – глобулінами. Перетворення кініногенів у кініни здійснюють специфічні ферменти – калікреїни. Калікреїни також знаходяться в неактивній формі – прекалікреїни – й активуються за участі фактора XII системи згортання крові. Кініни мають коротко-тривалу дію – період напіврозпаду цих молекул становить 20 – 30 с. Цей факт пояснюється високою активністю кініназ – ферментів, які їх розщеплюють. У зв’язку з тим, що кініни відіграють значну роль у патогенезі запалення, в клініці широкого використання набули лікарські препарати – інгібітори кініноутворення (контрикал, гордокс та інші). З легенів та слинних залоз бика виділений інгібітор калекреїнів – трасилол. Він також є інгібітором трипсину, тому його використовують у лікуванні гострих панкреатитів.
80 Небілкові азотовмісні компоненти крові
У плазмі крові азотисті продукти білкового обміну містяться в концентрації 14 – 28 ммоль/л, але при деяких фізіологічних станах (наприклад, споживання їжі, що збагачена азотовмісними сполуками) зростання концентрації до 35 ммоль/л не є ознакою патології.
Концентрацію цих азотовмісних сполук визначають після осадження білків крові трихлороцтовою (ТХО), фосфорно-молібденовою або фосфорновольфрамовою кислотами. Саме тому фракцію цих сполук називають залишковим азотом (rest nitrogen) крові.
До rest азоту крові входять сполуки, які є проміжними продуктами білкового обміну, а саме:сечовина (46 - 60% від загальної кількості залишкового азоту); амінокислоти ( до 25%); сечова кислота (4%);- креатинін (2,5 – 7,5%); креатин (5%),а також індикан, аміак, пептиди, нуклеотиди, нуклеозиди, білірубін, холін, гістамін та ін.
Сечовина (3,33-8,32 ммоль/л) утворюється в печінці як кінцевий продукт знешкодження аміаку. Ця сполука малотоксична, але при підвищенні концентрації сечовини, яка є осмотично активною речовиною, виникають набряки тканин паренхіматозних органів, міокарда, ЦНС. Підвищення концен-трації сечовини, яке супроводжується клінічним синдромом інтоксикації, має назву уремія. У клініці найбільше значення має визначення концентрації сечовини для діагностики захворювань нирок. Крім того, для диференціальної діагностики захворювань нирок та дистрофічних уражень паренхіми печінки використовують коефіцієнт Urea ratio.Креатин та креатинін – важливі компоненти залишкового азоту, які є необхідними складовими функціонування біоенер-гетичних молекулярних систем в організмі. З креатину в клітинах організму утворюється важливий макроерг – креатин-фосфат, який є депо метаболічної енергії. При ферментативному розщепленні креатинфосфату утворюється креатин, який знову підлягає рефосфорилюванню. Підвищення концентрації креатиніну може бути результатом підсиленого утворення (продукційна креатинінемія) або затримки цієї сполуки в організмі (ретенційна креатинінемія). Сечова кислота є кінцевим продуктом розпаду пуринових нуклеотидів, які входять до складу нуклеопротеїнів тканин організму та містяться в їжі. У нормі її концентрація в крові становить 0,12 – 0,38 мкмоль/л (для чоловіків) та 0,12 - 0,46 мкмоль/л (для жінок). Сечова кислота – це слабкокислий продукт, тому після секреції у кров вона утворює солі з лужними катіонами (98% з натрієм) - урати. У крові ці солі зв’язуються альфа-глобулінами, виводяться нирками (2/3) та через тонкий кишечник (1/3).
Підвищення вмісту сечової кислоти в крові також викликають: їжа, що збагачена пуринами (печінка, нирки, червоне вино), довготривале голодування, алкоголь, приймання діуретиків.Гіпоурикемія – зменшення концентрації сечової кислоти у крові, спостерігається при хворобі Вільсона-Коновалова, лімфогранулематозі, після приймання піперезину, саліцилатів, кортикотропіну. Індикан (калієва або натрієва сіль індоксилсірчаної кислоти) утворюється в печінці як результат знешкодження одного з продуктів гниття білків у кишечнику – індолу. Гіперіндиканемія спостерігається у хворих на початковій стадії розвитку ниркової недостатності, при гломерулонефриті, при підсиленні процесів гниття білків у кишечнику (защемленій грижі, запорі, кишковій непрохідності), пухлинах, абсцесах, туберкульозному ураженні легень, обтурації жовчних протоків тощо. Аміак – кінцевий продукт білкового обміну, який також є компонентом залишкового азоту. Його джерела – це процеси дезамінування, які відбуваються в тканинах, та катаболізм азотовмісних сполук у кишечнику під дією гнилісних бактерій. Концентрація аміаку в плазмі крові здорових людей дуже низька і становить 17-78 мкмоль/л. Підвищення концентрації аміаку в крові спостерігається при печінковій комі, дії гепатотропних отрут (чотирихлористого вуглецю, етіоніну та ін.), частковому видаленні печінки, білковому голодуванні, тепловому ударі, спадкових ензимопатіях орнітинового циклу, гострій печінковій недостатності, жировій інфільтрації печінки, гострій нирковій недостатності, кишковому дисбактеріозі тощо. Середньомолекулярні пептиди – це пептиди з молекул яр-ною масою 300-5000 Да, які також не осаджуються ТХО і входять до фракції залишкового азоту крові. Останнім часом цікавість до вивчення та визначення цих середніх молекул зросла у зв’язку з тим, що вони були охарактеризовані як універ-сальні фактори інтоксикації. Їх також називають молекулами середньої маси, або середніми молекулами. До них належать гормони, нейропептиди, медіатори імунної відповіді, деякі вітаміни, нуклеотиди, продукти розпаду фібрину та інші, деякі з них на цей час не ідентифіковані. Склад середньомолекулярних пептидів може бути різним, він залежить від виду патології та характеру ускладнень. Підвищення концентрації середніх молекул у крові спостерігається при всіх патологічних станах, які супроводжуються інтоксикацією (гнійних перитонітах, черепно-мозкових травмах, захворюваннях печінки, запаленні легень, гострій та хронічній нирковій недостатності). Для боротьби з інтоксикацією проводять гемо- та ентеросорбцію (у великих клініках використовують гемодіаліз), що знижує концентрацію пептидів та неполярних амінокислот у крові. При деяких патологічних станах залишковий азот крові зростає більше ніж на 28-35 ммоль/л. Такий стан називають азотемія або гіперазотемія. Гіперазотемія може бути абсолютною або відносною. Абсолютна гіперазотемія виникає внаслідок накопичення компонентів залишкового азоту. Причиною цієї азотемії може бути підсилення продукції відповідних азотовмісних компонентів або затримка (ретенція) кінцевих продуктів азотистого обміну. Тому абсолютна гіперазотемія може бути двох видів: ретенційна – розвивається при недостатньому виведенні азотовмісних сполук з організму. Цей вид азотемії поділяють на:
ниркову - спостерігається внаслідок порушення екскре-торної функції нирок (фракція залишкового азоту збільшується насамперед за рахунок зростання концентрації сечовини); позаниркову – розвивається при недостатності кровообігу, зниженні кров’яного тиску;
продукційна – є результатом надходження у кров надлишку азотовмісних продуктів, які утворюються в результаті підсиленого розпаду тканинних білків. Відносна гіперазотемія спостерігається при станах, які супроводжуються порушеннями водневого балансу (профузних проносах, підсиленому потовиділенні тощо).
81 ФУНКЦІОНАЛЬНІ ТА БІОХІМІЧНІ ВЛАСТИВОСТІ
СИСТЕМИ ГЕМОСТАЗУ
Система гемостазу складається з ферментів та додаткових біохімічних факторів плазми крові, тромбоцитів та інтими кровоносних судин. В умовах нормальної життєдіяльності утворення тромбів відбувається за умов пошкодження судинної стінки і має захисне значення, протидіючи витіканню рідкої крові з гемоциркуляторного русла. Внутрішньосудинне утворення тромбів (тромбоз) є патологічним процесом і має місце при тяжких порушеннях у регуляції системи гемостазу. За своєю структурою тромб — це утворення, що складається із сітки ниток нерозчинного білка фібрину та іммобілізованих у фібриновій сітці формених елементів крові. Розрізняють: білі тромби, утворені, переважно, скупченнями тромбоцитів; червоні тромби, до складу яких входить значна кількість еритроцитів.
Підтримання рідкого стану крові всередині кровоносних судин та протидія внутрішньосудинному згортанню крові забезпечуються антизгортальною системою крові, що складається з численних антикоагулянтів. Фібринові згустки, що можуть утворюватися в судинах, руйнуються ферментами фібринолітичної системи.
Із фізіологічної точки зору розрізняють:
1. Судинно-тромбоцитарний гемостаз, який є первинною реакцією на пошкодження судин мікроциркуляторного русла, що забезпечується судинною стінкою та тромбоцитами. Судинно-тромбоцитарному гемостазу належить провідна роль на початкових етапах зупинки кровотечі в зоні мікроциркуляторних судин. Цей тип гемостазу включає в себе: – рефлекторний спазм судин, що стимулюється судинозвужувальними сполуками, які виділяються з тромбоцитів (серотонін, адреналін, норадреналін)– адгезію тромбоцитів у зоні ушкодження судини. Поверхнею, до якої прикріплюються тромбоцити, є колагенова сітка ендотелію та субендотеліальної базальної мембрани. Відбувається взаємодія негативних ділянок на поверхні тромбоцитів+-групами лізилових залишків колагенових фібрил; – агрегацію тромбоцитів, що призводить до утворення тромбоцитарного тромбу.
Розрізняють фазу зворотної агрегації, яка стимулюється АДФ, та фазу незворотної агрегації, індукторами якої є тромбін, колаген та Са2+. Протидіють агрегації тромбоцитів та, відповідно, утворенню тромбу цАМФ та інгібітори циклооксигенази 2. Коагуляційний гемостаз, кінцевим результатом активації якого є утворення фібринового згустка, який іммобілізує формені елементи та утворює міцний тромб на поверхні ендотелію. Цей процес є основним типом гемостазу при кровотечах і реалізується складним ферментативним механізмом згортальної системи крові.
82
ЗГОРТАЛЬНА СИСТЕМА КРОВІ: КОМПОНЕНТИ,
МЕХАНІЗМИ АКТИВАЦІЇ
До складу згортальної системи крові входять ферментативні та неферментативні білки плазми, тканин, надмолекулярні комплекси, іони кальцію.Процес згортання крові та утворення кров’яного згустка становлять каскад послідовних ферментативних реакцій, що каталізуються спеціалізованими білками — факторами згортання. У каскадній системі згортання крові кожний білковий фактор спричиняє активації наступного компонента каскаду за послідовне лавиноподібне посилення процесу та реалізує швидку захисну реакцію
на травму судини. Поряд із зазначеними основними факторами коагуляції, що містяться переважно в плазмі крові, існують тромбоцитарні фактори коагуляції, які беруть участь на різних етапах судинно-тромбоцитарного, коагуляційного гемостазу та фібринолізу. Тромбоцитарні фактори коагуляції звичайно позначають латинською літерою Р (від англ. platelets — пластинки (кров’яні)) із цифровими індексами: P1-P11. Механізми активації та функціонування каскадної системи згортання крові
Каскадний ланцюг згортання крові може включатися за рахунок активації двох альтернативних механізмів: внутрішнього та зовнішнього шляхів (механізмів) коагуляції, що розрізняються початковими реакціями та конвергують в єдиний загальний шлях коагуляції, який починається з активації фактора Х. Центральною молекулярною подією в реалізації процесу коагуляції крові є утворення активного фактора Х (фактора Стюарта-Провера). Фактор Х — це Са2+-залежний
глікопротеїн, що синтезується в печінці при участі вітаміну К. Він скла- дається з легких та важких поліпептидних ланцюгів, що з’єднані дисуль-
фідними місточками, й активується шляхом обмеженого протеолізу. Активований фактор Х (ф.Ха) є сериновою протеїназою, що перетворює
протромбін (ф.II) в активний тромбін (ф.IIа), необхідний для трансфор-
мації фібриногену (ф.I) в фібрин —основу фібринового згустка або
тромбу. Взаємозв’язок між внутрішнім, зовнішнім та загальним кінцевим
шляхами в процесі згортання крові Внутрішній шлях коагуляції
1. Активація фактора ХII (ф. Хагемана) — відбувається при взаємодії крові з поверхнею за умов протеолітичної дії калікреїну, який відщеплює від ф.ХII пептидний фрагмент із молекулярною масою 28 кД, утворюючи активний фактор ХIIа. 2. Активація фактора ХI — відбувається під впливом фактора ХIIа, який утворює з фактора ХI фактор ХIа (активний тромбопластин плазми). 3. Активація фактора IХ (ф. Кристмаса) — відбувається під впливом фактора ХIа, який відщеплює від фактора IХ пептидний фрагмент із м.м. 9 кД, утворюючи активну серинову u1087 протеїназу — фактор IХа. Процес потребує присутності іонів Са2+.
4. Активація фактора Х — при функціонуванні “внутрішнього шляху” коагуляції активація фактора Х відбувається за рахунок протеолітичної дії фактора IХа, який відщеплює від фактора Х пептидні фрагменти з утворенням активних форм серинової протеїнази — фактора Ха, — яка складається з двох молекулярних форм (Ха-та Ха-). Процес перебігає при участі білка-модифікатора — фактора VIII (антигемофільного глобуліну А) — та іонів Са2+. 5. Активація фактора II (протромбіну) — перетворення протромбіну в тромбін (фактор IIа) відбувається під впливом протеїнази Ха за умов присутності іонів Са2+ та активної форми проакцелерину (фактора Vа). Активація протромбіну відбувається на поверхні тромбоцитів за участю фосфоліпідів тромбоцитарних мемб-
ран; присутність акцелерину (фактора Vа) тромбоцитів необхідна для зв’язування фактора Ха, вона збільшує швидкість реакції в десятки тисяч разів. Результатом процесу є утворення тромбіну — основного ферменту згортальної системи крові, який є сериновою протеїназою з м.м. 34 кД, що має трипсиноподібну активність. Протромбін людини складається з двох ланцюгів — легкого та; пептидні ланцюги в молекулі тромбіну
зв’язані з глюкозаміном та сіаловою кислотою. 6. Перетворення фібриногену в фібрин — заключний етап коагуляційного кас-
каду. Фібриноген — це глікопротеїн із м.м. 340 кД, що складається з шести поліпептидних ланцюгів (два А-ланцюги, два В-ланцюги та два -ланцюги; структура молекули фібриногену — (А)2 (B)22). Тромбін розщеплює чотири пептидні зв’язки типу –Arg–Gly– в молекулі фібриногену, що призводить до утворення вільних пептидів фібрину-мономера. дією ферменту трансглутамінази (фактора ХIII) відбувається зшивання окремих молекул фібрину-мономера з утворенням фібрину-полімеру. Така стабілізація фібринової сітки отримала назву ретракції згустка крові. Зовнішній шлях коагуляції активується за умов пошкодження кровоносних судин та оточуючих тканин і надходження в кров ліпопротеїнового тканинного фактора, який у сучасній літературі позначається як фактор III (тканинний тромбопластин).
Фактор III, що діє як білок-модифікатор, спричиняє активацію фактора VII (проконвертину), перетворюючи останній в активну протеїназу — фактор VIIа (конвертин). Конвертин разом з іонами Са2+ активує фактор Х, тобто запускає загальний шлях коагуляції за розглянутою вище схемою: Після утворення тканинного тромбопластину, що є швидкість-лімітуючим фактором згортання крові за механізмом зовнішнього шляху, подальша коагуляція відбувається надзвичайно швидко і кров’яний згусток утворюється протягом декількох секунд.
Роль вітаміну К у реакціях каскаду коагуляції
Вітамін К — жиророзчинний вітамін, що існує у вигляді двох вітамерів (К1 та К2), є необхідним кофактором реакцій коагуляції. При його відсутності порушується формування функціонально активних факторів згортальної системи —фф. II, VII, IX та X. Біохімічний механізм дії вітаміну К полягає в його участі у функціонуванні ферментної системи,
яка перетворює глутамінову кислоту пептидних ланцюгів зазначених факторів коагуляцію в -карбоксиглутамінову кислоту. Зокрема, в молекулі протромбіну відбувається -карбоксилювання
-Карбоксилювання білкових факторів коагуляції збільшує спорідненість їх молекул з іонами Са2+, які необхідні для зв’язування білків із мембранними фосфоліпідами та запуску каскаду коагуляції. Авітаміноз К супроводжується підвищеною кровоточивістю; декальцинована кров не згортається. Спадкові порушення процесу згортання крові
Порушення у функціонуванні системи згортання крові — коагулопатії можуть розвиватися внаслідок генетичного дефекту в синтезі плазмових чи тромбоцитарних факторів коагуляції і клінічно характеризуються зниженням згортальної активності крові, схильністю до кровотеч. Найбільш поширеними спадковими коагулопатіями є:
1. Гемофілії — коагулопатії, що виникають внаслідок спадкової відсутності одного або декількох факторів згортальної системи плазми. Гемофілії проявляються значними кровотечами, які з’являються навіть при незначних пошкодженнях кровоносних судин і є небезпечними для життя. Виділяють: – гемофілію А (хворобу Віллебранда) — розвивається за умов нестачі фактора VIII, синтез якого пов’язаний із Х-хромосомою; наслідується за рецесивним типом, проявляючись в осіб чоловічої статі;
– гемофілію В (хворобу Кристмаса) — розвивається за умов порушення синтезу фактора IX; – гемофілію С — розвивається при порушенні синтезу фактора XI, клінічно характеризується менш вираженими кровотечами, ніж гемофілії А та В. 2. А-(гіпо-)фібриногенемії — характеризуються повною або частковою відсутністю в плазмі фібриногену. Патологія наслідується як автосомна рецесивна хвороба, при якій спостерігаються тяжкі кровотечі внаслідок повної відсутності
здатності крові до коагуляції. 3. Дисфібриногенемії — коагулопатії, які виникають при амінокислотних замінах у первинній структурі молекул фібриногену. Аномальні молекули фібриногену мають змінену конформацію, що утруднює нормальний процес перетворення фібриногену у фібрин.
83.АНТИЗГОРТАЛЬНА СИСТЕМА КРОВІ
Існування в нормальних фізіологічних умовах рідкого стану крові зумовлене наявністю антизгортальної системи, що складається з антикоагулянтів — сполук, які протидіють внутрішньосудинній активації системи коагуляції. Антикоагулянти виконують функції інгібіторів певних білкових факторів згортання крові. Зниження вмісту цих інгібіторів у плазмі крові людини спричиняє підвищення схильності крові до згортання і може призводити до тромбозу. Антитромбіни — білки крові, що гальмують каталітичну активність тромбіну.
Найбільш потужним інгібітором тромбіну є антитромбін III — білок, антикоагулянтна активність якого значно зростає в присутності гепарину. Антитромбі III здатний зв’язуватися із сериновими протеїназами ферментного каскаду коагуляції крові, блокуючи, крім тромбіну, активність таких факторів, як IXа, Xа, XIа, XIIа.
1 — інгібітор протеїназ (1 — інгібітор трипсину) — глікопротеїн 1-глобулінової фракції плазми крові. Інгібітор має широкий спектр антипротеїназної дії, гальмуючи активність багатьох серинових протеїназ, зокрема, тромбіну факторів Ха та ХIа системи згортання крові.
2 — макроглобулін — глікопротеїн 2-глобулінової фракції плазми крові, що є інгібітором протеїназ із широкою субстратною специфічністю, блокуючи серинові, тіолові, карбокси- та металопротеїнази. Концентрація 2-макроглобуліну в плазмі крові людини (до 2,5 г/л) найвища, порівняно з іншими протеїназними інгібіторами. 2-Макроглобулін є інгібітором тромбіну, активність якого, на відміну від антитромбіну III, не залежить від дії гепарину. На долю цього інгібітора припадає до 25 % антитромбінової активності плазми крові. Гепарин — гетерополісахарид (глікозамінглікан), що є потужним природним антикоагулянтом. Його молекула побудована з дисахаридних фрагментів, що повторюються і складаються із залишків сульфатованої D-глюкуронової або
L-ідуронової кислоти та N-ацетилглюкозаміну; гепарин існує у формі поодиноких полісахаридних ланцюгів або у вигляді протеогліканів, тобто білків, які зв’язані з декількома u1075 глікозамінглікановими ланцюгами.
Гепарин синтезується тучними клітинами (гепариноцитами), що розташовані в печінці, легенях та впродовж стінок кровоносних судин. Механізм антикоагулянтної дії гепарину полягає в активації антитромбіну III: взаємодія з гепарином спричиняє конформаційну перебудову антитромбіну III, в результаті якої в останнього з’являється можливість зв’язуватися із сериновими протеїназами коагуляційного каскаду, блокуючи їх каталітичні активності.Кумарини — антикоагулянти природного (рослинного) та синтетичного походження, антагоністи вітаміну К. Вони є антикоагулянтами непрямої дії, протидіють утворенню біохімічно активних (-карбоксиглутамінованих) факторів коагуляції — II, VII, IX, X. У лікарській практиці для профілактики та лікування тромбозів застосовують такі похідні 4-оксикумарину, як Неодикумарин, Синкумар.
84 ФІБРИНОЛІТИЧНА СИСТЕМА КРОВІ
Фібриноліз — процес ферментативного розщеплення фібрину кров’яного
згустка, що супроводжується руйнуванням тромбу. Завдяки функціонуванню фібринолітичної системи відбувається постійне розчинення внутрішньосудинних тромбів, що можуть утворюватися на стінках кровоносних судин внаслідок дії факторів, які активують згортальну систему крові. Фібриноліз складається з двох послідовних етапів: I етап — утворення з неактивного проферменту плазміногену (профібринолізину) активного ферменту — протеїнази плазміну (фібринолізину), яка розщеплює фібрин тромбу. Плазміноген — глікопротеїн із класу -глобулінів із м.м. приблизно 80 кД, складається з одного поліпептидного ланцюга. Активація плазміногену з утворенням активного плазміну здійснюється за рахунок розщеплення протеїназами внутрішнього пептидного зв’язку. Фізіологічними активаторами цього глікопротеїну є фактор XIIа, численнні тканинні та судинні активатори плазміногену. Важливим активатором глікопротеїн без розщеплення внутрішньомолекулярного пептидного зв’язку має стрептокіназа — білок, що міститься в -гемолітичного стрептокока. Плазмін — фермент, що за механізмом ферментативної дії є сериновою протеїназою трипсиноподібної дії. Він є глікопротеїном із м.м. 80 кД, складається з
двох поліпептидних ланцюгів: важкого (А) та легкого (В). Фізіологічним субстратом плазміну є фібрин, проте фермент в умовах in vitro має широку субстратну специфічність, розщеплюючи різні білкові субстрати: фібриноген, білкові фактори комплементу, казеїн.
II етап — розщеплення фібрину, що є основою фібринового згустка, до пептидних продуктів протеолізу. Процес каталізується активним плазміном, що утворився на I етапі фібринолізу. Розчинні продукти деградації фібрину екскретуються із сечею. Підвищення вмісту в сечі продуктів розщеплення фібрину свідчить про надмірне посилення
внутрішньосудинного згортання крові. Утворення плазміну з плазміногену, що постійно відбувається в крові, є фізіологічним механізмом, який протидіє внутрішньосудинному тромбоутворенню. З
іншого боку, надмірному посиленню фібринолізу перешкоджає природний антагоніст плазміну — білковий інгібітор глікопротеїн 2-антиплазмін. Спадкове зниження концентрації 2-антиплазміну в крові супроводжується підвищеною кровоточивістю. У медичній практиці з метою лізису тромбів та профілактики тромбозів застосовують такі фармакологічні препарати компонентів фібринолітичної системи: 1. Фібринолізин (плазмін) — препарат, який отримують із профібринолізину (плазміногену) крові людини шляхом активації плазміногену трипсином. 2. Урокіназу — препарат, який виділяють із клітин нирок людини. 3. Стрептокіназу — препарат, який отримують із культури -гемолітичного стрептокока групи С. 4. Тканинний активатор плазміногена (ТАП) — препарат, який синтезують біотехнологічним методом за допомогою генної інженерії.
.85.Імуноглобуліни
Відомо 5 класів імуноглобулінів (Ig): IgG, IgM, IgA, IgD, IgE. Імуноглобуліни різних класів відрізняються за біологічними властивостями, а саме: за здатністю до зв’язування з антигеном.
Функції імуноглобулінів: IgG (59-75 мкмоль/л) – основні антитіла вторинного імунітету. Вони становлять 80% усіх імуноглобулінів сироватки крові. Тобто це основні імуноглобуліни сироватки крові, які забезпечують захист організму від багатьох бактерій, вірусів та їх токсинів. IgG – це єдині імуноглобуліни, які здатні проникати крізь плаценту в організм плода. Відомо чотири підкласи IgG, які відрізняються за структурою важких ланцюгів.
IgA (19-25 мкмоль/л) – трапляються в сироватці крові і є основними антитілами в серозно-слизових секретах (слині, бронхіальному слизі, сльозах, жіночому молозиві, слизовій оболонці кишечника).
IgM (0,8-1,2 мкмоль/л) – основні антитіла первинного імунітету. Це потужний активатор системи комплементу. Імуноглобулінами цього класу є антитіла Вассермана, ревматоїдний фактор, холодові аглютини та ізогемаглютини. Ці імуноглобуліни першими починають синтезуватися в організмі плода та при імунізації дорослих більшістю антигенів.
IgD (0,26 мкмоль/л) – представлені на поверхні В-лімфоцитів, беруть участь у пізнаванні антигену.
IgE (0,3-30 нмоль/л) – представлені на поверхні тучних клітин та базофілів. Існує точка зору, що вони беруть участь в імунному захисті від гельмінтів і в реакціях гіперчутливості миттєвого типу. До складу цієї фракції входять реагіни, які беруть участь в алергічних реакціях.
При електрофорезі імуноглобуліни рухаються в зоні γ-глобулінів, але IgA та IgM знаходяться у фракціях β- та α2- глобулінів.
IgG, IgA, IgD, IgE секретуються головним чином плазматичними клітинами, IgM – переважно лімфоцитами. Основну масу імуноглобулінів складають IgG.
86. Медіатори та гормони імунної системи.
За хімічною природою медіатори та гормони імунної системи є білками, глікопротеїнами, низько- та високомолекулярними пептидами, що присутні в плазмі крові людини і здійснюють свої регуляторні функції в пікомолярних (10–12) концентраціях. Значна кількість речовин, що виконують функції лімфокінів, до цього часу недостатньо охарактеризовані як індивідуальні хімічні сполуки.
Основні класи цитокінів
Інтерлейкіни (IL) — ростові фактори імунної системи: білки, що продукуються Т-лімфоцитами та макрофагами і стимулюють проліферацію лімфоцитів і деяких інших клітин організму. На даний час виділено більше десяти окремих типів інтерлейкінів (IL-1 — IL-16), що розрізняються за структурою, фізико-хімічними властивостями та спектром біологічної активності. Інтерлейкін-1, або фактор, що активує лімфоцити — медіатор, який продукується стимульованими макрофагами, а також поліморфноядерними лейкоцитами, епітеліальними клітинами шкіри, трансформованими клітинами хворих моноцитарною формою лейкемії. Головним ефектом дії IL-1 є стимуляція проліферації Т-хелперів та індукція секреції Т-хелперами інтерлейкіну-2. IL-1 є також активатором запалення як захисної рекції організму. Ця функція IL-1 пов’язана з його здатністю бути індуктором експресії генів фосфоліпази А2 та циклооксигенази, що є біохімічною передумовою стимульованого синтезу у вогнищах запалення простагландинів та лейкотрієнів; крім того, інтелейкін стимулює проліферацію фібробластів, синтез “білків гострої фази запалення” в гепатоцитах, колагенази в синовіальних оболонках суглобів. В організмі людини охарактеризовано два типи інтерлейкінів-1, що є білками, які складаються з 159 (IL-1) та 153 (IL-1) амінокислотних залишків. Молекулярна маса інтерлейкінів-1 дорівнює 12 кД. Інтерлейкін-2 — фактор, що продукується зрілими Т-хелперами в результаті їх стимуляції антигеном. IL-2 посідає центральне місце в системі інтерлейкінової регуляції імунітету, посилюючи процеси як клітинного, так і гуморального імунітету. Під впливом IL-2 відбувається диференціація Т-лімфоцитів в Т-кілери, що вбивають пухлинні та заражені мікробами клітини, а також активується продукція інтерферону Т- та NK-клітинами. Інтерлейкін-2 є глікопротеїном, що містить залишки сіалової кислоти; молекулярна маса IL-2 організму людини близько 13,5 кД. Інтерлейкін-3, або колонієстимулюючий фактор (CSF), синтезується активованими Т-хелперами. Основна мішень біологічної дії IL-3 — стовбурові гемопоетичні клітини — попередники лімфоцитів, ріст яких стимулює цей інтерлейкін. За хімічною природою інтерлейкін-3 є глікопротеїном, що складається з 134 амінокислотних залишків.
Інтерферони — білкові фактори, що синтезуються лімфоцитами та іншими клітинами тваринного організму при взаємодії з вірусами. Інтерферони є універсальними противірусними агентами, які активні відносно будь-яких вірусів, але мають видову специфічність, тобто кожному видові тварин притаманний свій клас інтерферонів. Розрізняють три типи інтерферонів, що продукуються різними клітинами і позначаються, як IFN-, IFN-та IFN-. IFN-синтезуються переважно лейкоцитами крові (“лейкоцитарні інтерферони”), IFN-— фібробластами (“фібробластні інтерферони”), IFN-— Т- та В-лімфоцитами (“імунні інтерферони”). Молекулярна маса різних інтерферонів коливається від 16 до 20-25 кД. IFN-та IFN-є одноланцюговими пептидами, що складаються з 166 амінокислотних залишків; IFN-містить 143 амінокислотні залишки. Інтерферони типу мають неглікозильовані пептидні ланцюги, інтерферони типів та є глікопротеїнами. Гени, що кодують IFN-та IFN-локалізовані на 10-й хромосомі каріотипу людини, ген IFN-— на 12-й хромосомі.
Взаємодіючи з клітинами, що інфіковані вірусними частинками, інтерферони інгібірують синтез вірусних білків, що призводить до блокування розмноження вірусу в зараженій клітині. Молекулярні механізми противірусної дії інтерферонів
1. Зв’язування інтерферонів із рецепторами клітин-мішеней. Інтерферони, які секретуються в зовнішньоклітинне середовище клітинами-продуцентами, взаємодіють із специфічними рецепторами на мембранах чутливих клітин, що є передумовою генерування хімічного сигналу та його трансмембранної передачі. 2. Активація внутрішньоклітинної 2',5'-олігоаденілатсинтетази. Хімічний сигнал за допомогою внутрішньоклітинних месенджерів досягає геному зараженої вірусом клітини й індукує синтез ферменту, який утворює з молекул АТФ 2',5'-олігоаденілову кислоту (2',5'-оліго-А-синтетази). 2',5'-олігоаденілат є активатором РНКази I, яка розщеплює односпіральні вірусні РНК (мРНК) та рибосомальні РНК, які необхідні для трансляції вірусних білків.
3. Активація внутрішньоклітинних протеїнкіназ. Трансмембранний хімічний сигнал, генерований інтерфероном, спричиняє також активацію протеїнкінази, що фосфорилює білковий фактор ініціації трансляції IF-2. Фосфорилювання фактора ініціації IF-2 призводить до його інактивації та блоку рибосомального синтезу вірусних білків.
Фактори некрозу пухлин (TNF). РозрізняютьTNF-та TNF-. TNF-(кахектин) — білок із м.м. 17 кД, продукується моноцитами та макрофагами. Його головними біологічними ефектами є індукція синтезу IL-1 та IFN-, цитотоксична та цитостатична дія. TNF
(лімфотоксин) — білок із м.м. 25 кД, продукується Т-лімфоцитами, спричиняє цитотоксичний ефект. При введенні в організм TNF спричиняють лізис деяких типів пухлинних клітин організму людини, їх розглядають як перспективні протиракові засоби. Колонієстимулюючі фактори (КСФ) — цитокіни, що стимулюють ріст кровотворних клітин (гранулоцитів, моноцитів, попередників еритроїдних клітин). Вони продукуються Т-лімфоцитами, макрофагами, моноцитами, ендотеліальними клітинами (див. IL-3). Трансформуючі фактори росту (ТФР) — білки, що продукуються різними класами лімфоцитів, тромбоцитами, плацентою, деякими пухлинами. Вони стимулюють процеси проліферації фібробластів, синтезу колагену та фібронектину, беруть участь в ангіогенезі, загоюванні ран. Разом із тим, ТФР пригнічують проліферацію Т та В-лімфоцитів, активність цитотоксичних та кілерних клітин.
87. Комплемент — ферментна система, необхідна для здійснення лізису чужорідних клітин (бактеріальних, тваринних) після їх взаємодії із специфічними антитілами. За біохімічною природою білки комплементу є каскадною системою протеаз, що послідовно активуються після утворення комплексу антиген-антитіло і спричиняють розщеплення мембранних структур клітин, які підлягають руйнації в процесі імунної реакції. Існує дев’ять основних компонентів (білкових субодиниць) системи комплементу, які позначають С1, С2, С3, С4, С5, С6, С7, С8, С9. Субодиниця С1 є комплексом, що складається з трьох білків: C1q, C1r, C1s. У фізіологічних умовах білкові компоненти системи комплементу знаходяться в плазмі крові в неактивному стані. Активовані форми білків комплементу позначаються рискою над відповідною латинською цифрою: С1, С2 тощо. Активація системи комплементу реалізується за одним із механізмів — класичним або альтернативним (рис. 30.2). За умов дії будь-якого з означених механізмів ключовою подією в активації системи комплементу є активація компоненту С3, який має аутокаталітичні властивості, запускаючи каскад ферментативних реакцій за участю термінальних компонентів С7, С8 та С9. Класичний шлях активації комплементу — послідовність ферментативних реакцій, що запускається при взаємодії першого компонента комплементу С1 з комплементзв’язуючими ділянками антитіл (IgG, IgM), які утворили комплекс антиген-антитіло на поверхні клітини. Зв’язування компонента С1 з імуноглобулінами, що фіксуються на поверхні клітини, відбувається з участю білка С1q, який отримав назву фактора розпізнавання. Взаємодія С1q з антитілом призводить до активації субодиниці С1r, яка розщеплює молекулу С1s, перетворюючи останню в активну серинову протеїназу. Таким чином реалізується пусковий механізм активації системи комплементу. Субстратами активного протеолітичного ферменту С1s є компоненти С4 та С2, які послідовно розщеплюються на відповідні молекулярні фрагменти: активний С4b і неактивний С4а та активний С2а і неактивний С2b. Активний комплекс С4bC2a позначають так: С3-конвертаза. Вона відщеплює від компонента С3 активну протеазу С3b (С5-конвертазу), яка активує систему термінальних компонентів комплементу (С5—С9). Активовані термінальні компоненти утворюють на поверхні клітинної мембрани надмолекулярний білковий комплекс (С5b, С6, С7, С8, С9), що спричиняє лізис мембрани, тобто розщеплення її білково-ліпідного бішару. Під впливом літичного комплексу в мембрані виникають канальці діаметром до 10 нм, через які відбувається вільне надходження всередину клітини Са2+ та Na+ і втрата К+, що і зумовлює остаточне руйнування клітини. Альтернативний (пропердиновий) шлях активації комплементу не потребує участі імуноглобулінів і має біологічне значення, ймовірно, на ранніх етапах боротьби макроорганізму з інфекційним агентом, тобто до продукування необхідної кількості антитіл. Як і в класичному шляху, в умовах альтернативного шляху також відбувається активація С3-компонента комплементу з утворенням С5-конвертази, але без участі компонентів С1, С2 та С4. Первинним етапом тут є взаємодія бактеріальних антигенів (полісахаридів, ліпополісахаридів клітинної стінки грамнегативних бактерій тощо) з білками системи пропердину. Система пропердину складається з трьох білків: білка Р — власне пропердину; білка В та білка D (серинової протеази). Під впливом антигенів бактеріальної стінки відбувається послідовна активація факторів Р, D та B. Активований фактор B (Bb) розщеплює основний компонент системи комплементу — С3, — утворюючи С5-конвертазу С3bBb, яка активує компонент С5, що відповідає за формування літичного мембранного комплексу.
88. Порушення у функціонуванні імунної системи людини — імунодефіцитні стани — розвиваються за умов пошкоджень окремих ланок клітинного або гуморального імунітету. За механізмом походження виділяють первинні та вторинні імунодефіцити.
Первинні імунодефіцити — патологічні стани, що виникають при спадковій нездатності організму людини реагувати на антигенне подразнення синтезом відповідних класів антитіл (імуноглобулінів) або формуванням клітинної імунної реакції. Вони розвиваються внаслідок молекулярних порушень у певних ділянках геному, що відповідають за фенотипічні прояви Т- та В-систем імунітету. Перші
прояви первинних імунодефіцитів спостерігають у ранньому дитячому віці. Розрізняють чотири класи первинних імунодефіцитів, що характеризують неповноцінність окремих основних компонентів імунної системи: 1. В-клітинна недостатність (дефіцит антитіл). 2. Т-клітинна недостатність. 3. Патологія клітин, що фагоцитують. 4. Патологія системи комплементу. Розглянемо приклади деяких найбільш поширених первинних імунодефіцитів людини. Хвороба Брутона — агаммаглобулінемія, зчеплена з Х-хромосомою. Імунодефіцит характеризується різким зниженням активності антибактеріального імунітету, що проявляється тяжким перебігом бактеріальних інфекцій. У сироватці крові хворих спостерігають значне зменшення концентрації IgG (приблизно в 10 разів, порівняно з нормою), IgA й IgM (приблизно в 100 разів), відсутність В-лімфоцитів та плазматичних клітин.
Швейцарський тип агаммаглобулінемії — хвороба, при якій наявний дефіцит як клітинного (Т-лімфоцити), так і гуморального (В лімфоцити) імунітету. Імунодефіцит спадкується за автосомно-рецесивним типом, проявляючись, переважно, в осіб чоловічої статі. Дисімуноглобулінемії — група первинних імунодефіцитів, що характеризуються різними варіантами порушень синтезу та секреції окремих класів імуноглобулінів (здебільшого IgG, IgM та IgA). Синдром Луї-Бар — імунологічна недостатність, що проявляється неврологічними порушеннями (атаксія мозочкового типу) та патологічним розширенням кровоносних судин кон’юнктиви і шкіри (телеангіектазії). У крові хворих спостерігають зниження активності реакцій клітинного імунітету, відсутність IgA та низький рівень IgG. Синдром Ди-Джорджи — уроджена гіпоплазія тимуса; захворювання характеризується порушенням Т-системи імунітету в дитячому віці. Вторинні імунодефіцити — патологічні стани, що розвиваються внаслідок ушкодження окремих ланок клітинного або гуморального імунітету патогенними факторами біологічного, хімічного або фізичного походження. Найчастіше вони розвиваються при дії на організм людини лімфотропної вірусної інфекції (СНІД, або синдром набутого імунодефіциту людини), токсичних факторів, іонізуючої радіації.
89. Біохімічні функції печінки в організмі
Вуглеводна (глікогенна) функція печінки
Ця функція полягає в здатності гепатоцитів утворювати лабільні резерви вуглеводів, що використовуються для підтримання необхідних концентрацій глюкози в крові та постачання цього цукру в інші органи (насамперед, головний мозок) у періоди між прийомами їжі.
Утворення й утилізація глюкозо-6-фосфату
Ініціюючим етапом включення глюкози в метаболічні перетворення є її фосфорилювання до глюкозо-6-фосфату (Г-6-Ф):
глюкоза + АТФ ______ глюкозо-6-фосфат + АДФ.
Ця ключова реакція вуглеводного метаболізму може каталізуватися в печінці двома ферментами: специфічною глюкокіназою та неспецифічною гексокіназою, що розрізняються за своєю спорідненістю до субстрату. Км для цих ферментів дорівнюють 0,01-0,1 мМ для гексокінази та 10 мМ для глюкокінази. Звідси зрозуміло, що при фізіологічних концентраціях глюкози (3,3-5,5 ммоль/л) каталітично активною є гексокіназа, а ферментативна дія глюкокінази включається лише за умов значного збільшення надходження в гепатоцити глюкози після споживання збагаченого на вуглеводи раціону .
Печінка є основним органом глюконеогенезу, тобто синтезу глюкози з невуглеводних метаболітів (лактату, глюкогенних амінокислот, гліцеролу), який активується в умовах вичерпання глікогенних резервів печінки (180-350 г глікогену) та зменшення (або відсутності) надходження цукрів із їжею. В організмі людини запаси вуглеводів використовуються приблизно за 12 год, разом з тим людина може витримати голодування протягом декількох місяців. У цих умовах забезпечення глюкозою тканин, енергетичний обмін яких значною
мірою залежить від цього моносахариду (головний мозок, інші нервові тканини, еритроцити, мозковий шар нирок, сім’яники), відбувається саме за рахунок глюконеогенезу в печінці та, частково, в нирках.
Перетворення на глюкозу інших моносахаридів
При змішаному харчуванні в кров людини з травного каналу, крім глюкози, всмоктується багато інших моносахаридів, зокрема D-фруктоза (надходить у складі сахарози), D-галактоза (у складі лактози молока) та D-маноза (у складі рослинних продуктів). Включення цих цукрів у загальний метаболізм відбувається завдяки наявності в гепатоцитах ферментних систем, що перетворюють вказані моносахариди на фосфорильовані ефіри глюкози та інтермедіати гліколізу.
Білоксинтезуюча функція печінки
Роль печінки в білковому обміні цілісного організму полягає в утворенні більшості білків плазми крові, які виконують важливі біохімічні та фізіологічні функції, регуляції розподілу амінокислот між окремими органами та тканинами, та синтезі сечовини як кінцевого продукту азотистого катаболізму.
(1) В гепатоцитах синтезуються всі альбуміни плазми крові (13-18 г/добу), які беруть участь у підтриманні нормального онкотичного тиску плазми, транспорті багатьох метаболітів та інших біомолекул; гіпоальбумінемія є інформативною клініко-діагностичною ознакою гострої та хронічної недостатності печінки. (2) В печінці синтезується більша частина (близько 80 %) глобулінів плазми: гепатоцити беруть участь у біосинтезі певної частини -глобулінів, ретикулоендотеліальні клітини продукують - глобуліни та частину -глобулінів (імуноглобулінів); захворювання печінки, при яких наявні важкі порушення структури та функцій органа, супроводжуються зниженням концентрацій у крові 1-, 2- та - глобулінів; з іншого боку, патологічні процеси, які перебігають з активацією імуно-компетентних клітин печінки, призводять до збільшення рівня -глобулінів. (3) В клітинах печінки синтезується багато білкових факторів, що входять до складу згортальної, антизгортальної та фібринолітичної систем крові: V, XI, XII, XIII фактори згортання, компоненти протромбінового комплексу (II, VII, IX, X фактори), фібриноген, антитромбін, антиплазмін, гепарин. (4) Завдяки активному перебігу реакцій обміну амінокислот (трансамінування, дезамінування, декарбоксилювання), печінка бере участь у підтриманні відносної біохімічної сталості амінокислотного складу крові; порушення білоксинтезуючої функції гепатоцитів (зокрема, при дії хімічних та біологічних пошкоджуючих факторів на рибосомальну систему трансляції) супроводжується значним збільшенням концентрації вільних амінокислот у плазмі крові; рівень аміноазоту плазми (в нормі 2,9 4,3 ммоль/л) може у хворих з важкою печінковою недостатністю збільшуватися до 21 ммоль/л, що супроводжується вираженою аміноацидурією. Сечовиноутворювальна функція печінки
Печінка є єдиним органом, що містить повний набір ферментів утворення сечовини з продуктів азотистого (переважно білкового) катаболізму. Порушення функціонування циклу сечовиноутворення, що спричиняються екзогенними ушкоджуючими\ факторами, або спадковими ензимопатіями (генетичними дефектами в синтезі окремих ферментів біосинтезу сечовини) призводять до накопичення в крові та тканинах вільного аміаку. Найчутливішими до такої патобіохімічної ситуації є нейрони головного мозку, в яких надлишковий аміак здатний до пригнічення функціонування циклу трикарбонових кислот за рахунок взаємодії NH4+ з -кетоглутаратом у реакції відновлювального амінування. Гальмування реакцій ЦТК та відповідне зниження рівня АТФ в нервовій тканині спричиняють деполяризацію мембран нейронів, порушення синаптичної передачі, що клінічно проявляється розвитком печінкової енцефалопатії та коматозного стану.
90. Жовчоутворювальна та пігментна функції печінки
Важливою для фізіології та патології організму людини є роль, яку печінка відіграє в катаболізмі гемоглобіну та інших гемовмісних білків, при розщепленні яких утворюються жовчні пігменти — білірубін і білівердин, що екскретуються через кишечник (див. розділ 31.3). Ці сполуки разом з іншими органічними речовинами (жовчними кислотами, холестерином, фосфоліпідами), які продукуються гепатоцитами, входять до складу жовчі, надаючи їй специфічного золотисто жовтуватого кольору. Біохімічний склад жовчі Жовч — це рідкий секрет клітин печінки, що служить як для надходження в дванадцятипалу кишку поверхнево активних сполук (жовчних кислот, фосфоліпідів), необхідних для перетравлювання і всмоктування нейтральних жирів, так і для екскреції з організму кінцевих продуктів катаболізму біомолекул і ксенобіотиків. За добу в дорослої людини утворюється 500-700 мл жовчі; вміст основних біоорганічних сполук в печінковій та міхуровій жовчі
91. Детоксикаційна функція печінки
Однією з найважливіших біологічних функцій печінки є знешкодження — детоксикація хімічних сполук, що не є нормальними метаболітами організму і можуть спричиняти несприятливу дію, здійснюючи загальнотоксичні, некрозогенні, мутагенні, канцерогенні ефекти. Біохімічна сутність процесу детоксикації, що відбувається в гепатоцитах, полягає в переведенні (біотрансформації) хімічної сполуки в результаті певних ферментативних реакцій в молекулярну форму з менш вираженими токсичними властивостями; продукти біотрансформациї токсинів у печінці є, як правило, більш водорозчинними (гідрофільними) речовинами, що можуть виводитися з організму різними системами екскреції (нирками, кишечником, легенями, шкірою). Ця функція, у зв’язку з її надзвичайною важливістю для збереження хімічного гомеостазу внутрішнього середовища організмів людини та тварин, детально розглядатиметься нижче.
92. Реакції мікросомального окислення
Головна роль серед реакцій першої фази біотрансформації ксенобіотиків та ендогенних сполук належить ферментним системам мембран ендоплазматичного ретикулума, що функціонують за участю цитохрому Р-450. У зв’язку з тим, що біохімічним еквівалентом мембран ендоплазматичного ретикулума клітин печінки є отримувана методом диференційного центрифугування мікросомальна фракція, тип реакцій, який розглядається, отримав у науковій літературі назву “реакцій мікросомального окислення”, а відповідні ферменти — “мікросомальних оксигеназ. Реакції, що каталізуються цими ферментами, належать до типу монооксигеназних, тобто таких, що каталізують включення атома одного кисню безпосередньо в молекулу субстрату, який окислюється (глава 9):
R–H + 1/2 O2 ______ R–OH. Така реакція (окислювальне гідроксилювання) є основною в метаболізмі гідрофобних сполук у мембранах ендоплазматичного ретикулума гепатоцитів і вимагає участі в ролі донора електронів НАДФН:
НАДФН + R-H + O2 ______ НАДФ+ + R-OH + H2O.
Оскільки в розглянутому процесі один із атомів молекули кисню включається в молекулу води, а другий — в молекулу субстрату, що гідроксилюється, ферментні системи, які каталізують ці реакції, отримали також назву “мікросомальних окси-
геназ мішаної функції”.
Цитохром Р-450
Ферментні системи, що каталізують реакції мікросомального окислення гідро- фобних субстратів, є електронотранспортними ланцюгами, локалізованими в мембранах ендоплазматичного ретикулума гепатоцитів (та клітин деяких інших органів, що також беруть участь у реакціях детоксикації). Компонентами цих ферментних ланцюгів є ФАД-вмісний флавопротеїн, цитохром b5 та кінцева монооксигеназа — цитохром Р-450:
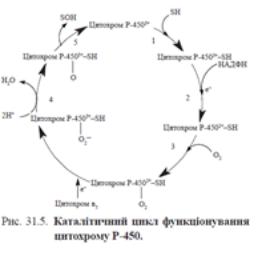 НАДФН
______ФП
(ФАД) ______цитохром
b5
______цитохром
Р-450 Подібний цитохром Р-450-залежний
електронотранспортний ланцюг каталізує
реакції окислювального гідроксилювання
стероїдів (синтезу та біотрансформації),
що наявні в мітохондріях кори наднирникових
та статевих залоз. Цитохром Р-450 —
фермент, вперше відкритий у 1958 р.
Американськими дослідниками Д.Гарфінкелем
та М.Клінгенбергом (D.Garfinkel,
M.Klingenberg).
Це сімейство гемопротеїнів з молекулярною
масою близько 50 кД; у різних біологічних
об’єктах та тканинах виявлено більше
300 ізоформ цитохрому Р-450, що розрізняються
за своєю субстратною специфічністю та
особливостями первинної структури.
Фізіологічне значення ізоформ цитохрому
Р-450 полягає в захисті тваринного
організму від численних низькомолекулярних
ксенобіотиків, що надходять у внутрішнє
середовище; ця система є додатковою до
системи імунного
захисту
(“друга
імунна система”), яка протидіє надходженню
в організм чужорідних високомолекулярних
сполук біологічного походження.
Каталітичний цикл, що реалізує реакції
окислювального гідроксилювання
субстратів (SH)
за участю цитохрому Р-450, складається з
декількох парціальних реакцій, поданих
на рис. 31.5. Реакції
кон’югації в гепатоцитах
Реакції кон’югації з утворенням “парних”
сполук — шлях детоксикації більшості
ксенобіотиків, що мають функціональні
групи –OH,
–COOH,
–NH2,
–SH
(або утворюють їх у реакціях 1-ї фази
біотрансформації). Цей же механізм
використовується для утворення
молекулярних форм, що підлягають
екскреції з організму, з таких ендогенних
субстратів, як жовчний пігмент білірубін,
продукти бактеріального розщеплення
в кишечнику циклічних амінокислот
(фенолу, крезолу, індоксилу), стероїдні
гормони та продукти їх гідроксилювання,
продукти моноамінооксидазного розщеплення
катехоламінів, серотоніну та інших
біогенних амінів. Найбільш поширеними
реакціями кон’югації є: 1.
Реакції
глюкуронування (основний
тип кон’югації в організмі людини та
тварин як ксенобіотиків, так і ендогенних
субстратів) Фермент, що каталізує реакції
глюкуронування — УДФ-глюкуронілтранс-
фераза.
2.
Реакції
сульфування,
донором сульфатних радикалів у яких є
біологічно активна форма сірчаної
кислоти — 3'-фосфоаденозин-5'-фосфосульфат
(ФАФС). Прикладом реакції сульфування
є утворення в печінці кон’югату на
основі індоксилу —
продукту мікробного розщеплення в
товстій кишці амінокислоти L-триптофану.
3.
Реакції
метилювання та ацетилювання —
поширений тип кон’югації, в яких беруть
участь як ксенобіотики, так і ендогенні
субстрати; в цих реакціях беруть участь
S-аденозилметіонін
(О-метилювання) та ацетил-КоА (N-ацетилювання)
4.
Реакції
кон’югації з гліцином;
клінічно важливим прикладом реакції є
утворення гіпурової
кислоти при
взаємодії ендогенного гліцину з введеною
в організм бензойною кислотою
НАДФН
______ФП
(ФАД) ______цитохром
b5
______цитохром
Р-450 Подібний цитохром Р-450-залежний
електронотранспортний ланцюг каталізує
реакції окислювального гідроксилювання
стероїдів (синтезу та біотрансформації),
що наявні в мітохондріях кори наднирникових
та статевих залоз. Цитохром Р-450 —
фермент, вперше відкритий у 1958 р.
Американськими дослідниками Д.Гарфінкелем
та М.Клінгенбергом (D.Garfinkel,
M.Klingenberg).
Це сімейство гемопротеїнів з молекулярною
масою близько 50 кД; у різних біологічних
об’єктах та тканинах виявлено більше
300 ізоформ цитохрому Р-450, що розрізняються
за своєю субстратною специфічністю та
особливостями первинної структури.
Фізіологічне значення ізоформ цитохрому
Р-450 полягає в захисті тваринного
організму від численних низькомолекулярних
ксенобіотиків, що надходять у внутрішнє
середовище; ця система є додатковою до
системи імунного
захисту
(“друга
імунна система”), яка протидіє надходженню
в організм чужорідних високомолекулярних
сполук біологічного походження.
Каталітичний цикл, що реалізує реакції
окислювального гідроксилювання
субстратів (SH)
за участю цитохрому Р-450, складається з
декількох парціальних реакцій, поданих
на рис. 31.5. Реакції
кон’югації в гепатоцитах
Реакції кон’югації з утворенням “парних”
сполук — шлях детоксикації більшості
ксенобіотиків, що мають функціональні
групи –OH,
–COOH,
–NH2,
–SH
(або утворюють їх у реакціях 1-ї фази
біотрансформації). Цей же механізм
використовується для утворення
молекулярних форм, що підлягають
екскреції з організму, з таких ендогенних
субстратів, як жовчний пігмент білірубін,
продукти бактеріального розщеплення
в кишечнику циклічних амінокислот
(фенолу, крезолу, індоксилу), стероїдні
гормони та продукти їх гідроксилювання,
продукти моноамінооксидазного розщеплення
катехоламінів, серотоніну та інших
біогенних амінів. Найбільш поширеними
реакціями кон’югації є: 1.
Реакції
глюкуронування (основний
тип кон’югації в організмі людини та
тварин як ксенобіотиків, так і ендогенних
субстратів) Фермент, що каталізує реакції
глюкуронування — УДФ-глюкуронілтранс-
фераза.
2.
Реакції
сульфування,
донором сульфатних радикалів у яких є
біологічно активна форма сірчаної
кислоти — 3'-фосфоаденозин-5'-фосфосульфат
(ФАФС). Прикладом реакції сульфування
є утворення в печінці кон’югату на
основі індоксилу —
продукту мікробного розщеплення в
товстій кишці амінокислоти L-триптофану.
3.
Реакції
метилювання та ацетилювання —
поширений тип кон’югації, в яких беруть
участь як ксенобіотики, так і ендогенні
субстрати; в цих реакціях беруть участь
S-аденозилметіонін
(О-метилювання) та ацетил-КоА (N-ацетилювання)
4.
Реакції
кон’югації з гліцином;
клінічно важливим прикладом реакції є
утворення гіпурової
кислоти при
взаємодії ендогенного гліцину з введеною
в організм бензойною кислотою
93. ОБМІН ЖОВЧНИХ ПІГМЕНТІВ. БІОХІМІЯ ЖОВТЯНИЦЬ
В організмі людини йде постійне руйнування зрілих еритроцитів, швидкість якого складає (1-2) · 108/год; середня тривалість життя еритроцитів — 100-120 днів. Жовчні пігменти (білірубін, білівердин) є продуктами катаболізму гемоглобіну еритроцитів та частково — інших гемовмісних білків. Руйнування еритроцитів та розщеплення гемоглобіну, який складає 90-95 % сухого залишку еритроцитів, відбувається в клітинах ретикулоендотеліальної системи органів та тканин людини — селезінці (переважно), купферовських клітинах печінки, кістковому мозку, гістіоцитах сполучної тканини.
Патобіохімія жовтяниць
Перевищення концентрації загального білірубіну сироватки крові людини понад 2-4 мг % проявляється характерною жовтуватістю шкіри та слизових оболонок (особливо склер очного яблука) і позначається як жовтяниця (icterus — лат.). Причинами виникнення гіпербілірубінемії і розвитку жовтяниць є надмірне утворення білірубіну в організмі, пошкодження печінки з порушенням її детоксикаційної та екскреторної функцій або наявність механічних перешкод в системі жовчовивідних шляхів, що протидіють нормальному виведенню жовчі в кишечник. Відповідно розрізняють декілька типів жовтяниць. Передпечінкова (гемолітична) жовтяниця — розвивається внаслідок патологічно підсиленого руйнування (гемолізу) еритроцитів та розщеплення гемоглобіну і надмірного накопичення в крові білірубіну. Причинами такого стану можуть бути резус-конфлікт у новонароджених, переливання несумісної крові, радіаційне ураження, дія гемотоксичних отрут тощо. Для передпечінкової жовтяниці характерним є збільшення концентрації в крові загального білірубіну, переважно за рахунок непрямої фракції, тобто вільного білірубіну, який не встигає бути кон’югованим у печінці в умовах його надмірного
утворення. Надходження в цих умовах значних кількостей білірубіну в кишечник призводить до посиленого утворення стеркобіліногену (в деяких випадках — до 10 г), що в збільшеній кількості виділяється з калом (стеркобілін) та (після всмоктування в товстій кишці) з сечею (уробілін сечі). Печінкова (паренхіматозна) жовтяниця — розвивається внаслідок порушення структури та ферментативних властивостей гепатоцитів в результаті дії пошкоджувальних факторів вірусного, бактеріального, хімічного походження (вірусні, інфекційні, токсичні гепатити). При цьому типі жовтяниць спостерігається значна гіпербілірубінемія (збільшення концентрації загального білірубіну) внаслідок таких причин: а) порушення кон’югації білірубіну як результат пошкодження мембран ендоплазматичного ретикулума гепатоцитів і зменшення активності УДФ-глюкуронілтрансферази; дія цього фактора призводить до зростання рівня в крові непрямого білірубіну;
б) порушення секреторної функції гепатоцитів, тобто їх здатності транспортувати білірубін-глюкуронід у жовч (транспорт проти градієнта концентрації); ця обставина, а також некроз печінкових клітин призводять до надходження в сироватку крові надмірної кількості прямого білірубіну. Зростання в крові прямого білірубіну (білірубін-глюкуроніду), що здатен проходити через ниркові мембрани, супроводжується (у важких випадках захворювання) появою білірубін-глюкуроніду в сечі (лабораторні проби на наявність жовчних пігментів у сечі стають позитивними). Внаслідок порушення здатності гепатоцитів до розщеплення тетрапіролів (що всмоктуються у вигляді мезобіліногену), ці сполуки також надходять у сечу (підвищена реакція на “уробілін” сечі).
Післяпечінкова (обтураційна) жовтяниця — спричиняється неможливістю надходження жовчі в дванадцятипалу кишку внаслідок закупорки жовчних шляхів (наявність пухлин, жовчнокам’яної хвороби). Цей тип жовтяниць характеризується знебарвленням калових мас внаслідок відсутності в них стеркобіліногену (“ахолічний” кал) та цілковитою відсутністю уробіліну в сечі. Внаслідок утрудненого надходження білірубін-глюкуроніду в жовч (зростання гідростатичного тиску в жовчних шляхах), кон’югований пігмент у збільшеній кількості всмоктується в кров, що призводить до зростання рівня прямого білірубіну; в цих умовах можлива поява жовчних пігментів у сечі, як при паренхіматозній жовтяниці. Ферментативні (спадкові) жовтяниці — виникають внаслідок генетичних ензимопатій, що спричинені порушеннями експресії генів, які відповідають за синтез у гепатоцитах ферментів кон’югації білірубіну (УДФ-глюкуронілтрансферази та/або УДФГ-дегідрогенази — ферменту, що утворює УДФГК з УДФ- глюкози), його абсорбції з крові або екскреції в жовч. Некон’югований білірубін, що накопичується в сироватці крові в надмірній кількості, за цих типів жовтяниць може проникати через гематоенцефалічний бар’єр в головний мозок і відкладатися в базальних гангліях та ядрах стовбура мозку, спричиняючи важкі неврологічні зрушення (“ядерні жовтяниці”).
96.Ренин-ангиотензиновая система (РАС) или ренин-ангиотензин-альдостероновая система (РААС) — это гормональная система человека и млекопитающих, которая регулирует кровяное давление и объём крови в организме.Компоненты системы:Ангиотензиноген,Ангиотензин I,Ангиотензин II,Проренин,Ренин,Ангиотензин-конвертирующий фермент,Альдостерон.Почками секретируется ренин,который влияет на ангиотензиноген и он превращается в ангиотензин 1,который в свою очередь превращается в ангиотензин 2 под влиянием спицефического фермента.Ангиотензин 2 увеличивает секрецию альдостелона.Ангиотензин 2 и альдостерон уменьшают количество выделяющейся воды из органзма.
97.склад сечі Колір від солом`яно-жовтого до насиченого жовтого; зумовлений пігментами: урохромом (95% забарвлення), уробіліном, уроеритрином, урозеїном та ін. Червоний колір (рожево-червоний)– при гематурії чи гемоглобінурії та після прийому амідопірина, антипірина та ін. лік. засобів. Коричневий чи червоно-бурий – при ↑конц. уробіліна і білірубіна. Зелений або синій – при веденні метиленового синього і при гнитті білків у кишечнику→ ↑конц. в крові індоксилсірчаних кислот→ розкладаються з утворенням індіго). Мутність може бути викликана солями, клітинними елементами, бактеріями, слизом, жиром (ліпурія). Відносна щільність – 1,002-1,035 (частіше 1,012-1,020). При важкій недостатності нирок – ізостенурія – сеча низької щільності (ультрафільтрат, первинна сеча). рН 5,3-6,5. Кисла р-ція КН2РО4, NаН2РО4; лужна – К2НРО4, Nа2НРО4, КНСО3, NаНСО3. Сечовина – за добу виводиться ~30г сечовини – 80-90% азота сечі. При ураженнях печінки ↓сечовини в сечі. Креатинін – утвор. із креатинфо
98.Структурною одиницею м’яза є міозит.Міоцит має скорочувальні елементи – міофібрили.Скорочувальні білки – актин,міозин. Саркомер – структурно-функціональні елементи скорочувального апарату скелетних м*язів.Міозин – фібрилярний білок,що утворює товсті філаменти міофібрил.Складаеться з 2 важких та 4 легких поліпептидних ланцюгів.Актин – білок,що існує в 2 формах :G- та F-актину. G-актин – глобулярний білок,сполучаючись між собою вони утворюють ланцюги фібрилярного F-актину.Ф-актин – основа тонких ниток саркомера.Тропоміозин – білкові молекули витягнутої форми,що складається з двох поліпептидних ланцюгів(альфа та бета),які утворюють спіраль.Тропонін – білок тонких філаментів,що складається з 3 субодиниць:TnT,TnI,TnC.Тропонінові комплекси мають глобулярну форму і роміщюються впродовж акти нового філамента. TnC – кальційзв*язуючий білок. TnI- взаємодіє з актином. TnT- забезпечує взаємодію тропонінового комплексу з тропоміозином.
99.мязове скороченняА.У м*язі,що перебуває в стані спокою,С1-головки міозину не сполучені з активними філаментами.Продукти гідролізу АТФ зв*язані з міозином.В.При збудженні м*яза С1-головки зсуваються в напрямку тонких філаментв і сполучаються з нитками актину.Фн вивільняється з комплексу з міозином.С.Вивільнення АДФ з комплексу з міозином супроводжується конфірмаційним зсувом у просторовому розташуванні головки С1,що зв*язана з актином.Зміна положення головки призводить до розвитку напруги і пересування тонкого філаменту відносно товстого приблизно на 10нм у напрямку середини саркомера.Д.Взаємодія з актином молекули АТФ супроводжується розривом зв*язку між актином і міозином.С1-головка знову віддаляється від тонкого філамента.
100.біоенергетика м язаДжерела АТФ:1.глікогеноліз,який постачає глюкозо-6-фосфат,що окислюється гліколітичним шляхом за аеробним чи анаеробним механізмом.2.Окислення глюкози,яка надходить у м*язи з крові.3.Окисне фосфорилювання в саркосомах(найбільше енергії).4.Аденілат-кіназна реакція:2АДФ-АТФ +АМФ.Креатинфосфокіназна реакія :креатин фосфат-креатин+АТФ.
102.Білки волокон сполучної тканини Колаген — головний білок сполучної тканини тварин та білок, що має найбільший вміст у ссавців, до 25 % від повної маси білків організму. Слово «колаген» походить від грецького κόλλα [колла] – клей. Молекула колагену являє собою правозакручену спіраль з трьох α-ланцюгів. Таке утворення відоме під назвою тропоколагену[1]. Один виток спіралі α-ланцюга містить три амінокислотних залишки. Молекулярна маса колагену становить близько 300 кДа, довжина 300 нм, товщина 1,5 нм.Для первинної структури білка характерний високий вміст гліцину, низький вміст сірковмісних амінокислот і відсутність триптофану. Колаген належить до тих небагатьох білків тваринного походження, які містять залишки нестандартних амінокислот: близько 21% від загального числа залишків припадає на 3-гідроксипролін, 4-гідроксипролін та 5 -гідроксилізин Кожен з α-ланцюгів складається з тріад амінокислот. У тріади третя амінокислота завжди гліцин, друга — пролін або лізин, перша — будь-яка інша амінокислота, крім трьох перерахованих Колаген існує в кількох формах. Основна структура всіх типів колагену є схожою. Колагенові волокна утворюються шляхом агрегації мікрофібрил, мають рожевий колір при забарвленні гематоксиліном і еозином і блакитний або зелений при різних трьоххромних забарвленнх, при імпрегнації сріблом фарбуються в буро-жовтий колір. Эласти́н — белок, обладающий эластичностью и позволяющий тканям восстанавливаться, например, при защемлении или порезе кожи. Эластин – белок, отвечающий за упругостьЭластин – белок, отвечающий за упругость соединительных тканей.В составе эластина также присутствуют уникальные белки – десмозины. Присоединяясь к клеткам кожи, они образуют жесткий каркас. В основном, эластин состоит из глицина, валина, аланина и пролина. Эластин является специализированным белком с молекулярной массой от 64 до 66 кДа. Эластин синтезируется клетками фибробластов через предшественник — растворимый тропоэластин, не содержащий поперечных связей. Из множества тропоэластина эластин получается в реакции, катализируемой лизилоксидазой. В эластине также присутствуют десмозин и изодесмозин, из-за чего эластин может растягиваться в двух направлениях. В поджелудочной железе синтезируется эластаза.Название фермент получил от субстрата эластина,который он гидролизует. Глікопротеїни (застаріла назва — глікопротеїди) — складні білки, в яких білкова (пептидна) частина молекули ковалентно сполучена з однією або декількома гетероолігосахаридними групами (гліканами). Моносахариди, пов'язані з конкретним білком, можуть бути різними: це може бути глюкоза, фруктоза, маноза, глюкозамін, галактозамін, фруктозамін, сіалова кислота та інші.Ті або інші моносахариди, пов'язані з білком, змінюють біохімічні та імунологічні властивості білка, його просторову конфігурацію та інші властивості. Важливим окремим випадком є зв'язування білків з сіаловою кислотою, що приводить до формування сіалоглікопротеїнів. У типових випадках саме зв'язування з сіаловою кислотою зумовлює збільшення T1/2 білка в плазмі крові. Глікопротеїни є важливим структурним компонентом клітинних мембран тваринних і рослинних організмів. До глікопротєїнів відносяться більшість пептидних гормонів. Глікопротеїни мембран еритроцитів, специфічно глікозільовані тими або іншими вуглеводними залишками, але що мають гомологічну білкову частину, зумовлюють групу крові у людини. Також глікопротеїнамі є все антитіла, інтерферони, компоненти комплемента, білки плазми крові, молока, рецепторні білки багато інших. Протеогликаны — гликопротеины с высокой степенью гликозилирования, углеводные остатки которых представляют собой длинные неразветвленные полисахаридные цепи — гликозаминогликаны, образованные чередующимися остатками гексозамина и уроновой кислоты (глюкуроновой, идуроновой или галактуроновой) либо галактозы. Гликозаминогликановые цепи гликопротеинов зачастую сульфированыПротеогликаны являются одним из основных компонентов внеклеточного матрикса соединительной ткани.Протеогликаны состоят из сердцевинных белков (лат. core protein) с молекулярной массой от 10 до 600 кДа, с которыми ковалентно связаны несколько гликозаминогликановых цепей (до 80 гликозидных звеньев массой до 50 - 60 кДа); число связанных с белком полисахаридных цепей варьирует от одной (декорин фибробластов) до ста и выше (аггрекан хрящевой ткани). Такое строение обуславливает высокую молекулярную массу протеогликанов (у аггрекана до 5 МДа), при этом на белковую часть приходится ~5-10% и на углеводную ~90-95% массы.
103.Біохімія нервової системи.У головному мозку наявні білки(8%) значна кількість ліпідів (10-12%) вуглеводи (1%) інші низькомолекулярні сполуки неорганічні солі і води.До ліпідів головного мозку належать:фосфогліцерини,сфінголіпіди,гліколіпіди),холестерин,триацелгліцероли.У білій речовині ліпідів більне ніж у сірій.Білки головного мозку різноманітні і поділяються на білки-ферменти, регуляторні й структурні білки.Особливо виділяють нейроальбуміни, нейроглобуліни, нейросклеропротеїни.
104.Енергетичний обмін головного мозкуБіоенергетика мозку значними чином залежить від постачання киснем,який використовується переважно для аеробного окислення глюкози.мозок поглинає 20-25% усього кисню.Основна потреба в АТФ полягає у генерації нервового потенціалу,для якого потрібно постійне функціонування натрієвого насоса.Основним джерелом енергії для мозку є глюкоза.Перехід на кетонові тіла спостерігається при голодуванні та виснажливій фізичній роботі.Аміак який з*являється в наслідок метаболізму знешкоджується за допомогою глутамату.
105.Біохімія нейромедіаторів Нейромедіатори або нейротрансмітери — біологічно активні хімічні речовини, за допомогою яких здійснюється передача електричного імпульсу з нервової клітини через синаптичний простір. Нейромедіатори характеризуються здатністю реагувати із специфічними білковими рецепторами клітинної мембрани, ініціюючи ланцюг біохімічних реакцій, що викликають зміну трансмембранного струму іонів, що приводить до деполяризації мембрани і виникнення потенціалу дії.До нейромедіаторів відносять як деякі малі молекули (катехоламіни): серотонін, дофамін, адреналін, норадреналін, гістамін, ацетилхолін, глютамат, аспартат, гліцин, ГАМК, ендоканабіноїд, N-ацетиласпартилглутамат, так і деяку кількість пептидів (біля 50): вазопресин, соматостатин, нейротенсин.
Рецептори – мембранні білки,що локалізовані в постсинаптичних мембранах нейронів або плазматичних мембранах клітин ефекторних органів і здатні до зв*язування фізіологічних ефекторів(нейромедіаторів) і передавання зовнішньо клітинного хімічного сигналу всередину нейрона.Вони поділяються на іонотропні(іонні канали) на метаботропні(цАМФ,цГМФ) рецептори.
107.Порушення обміну медіаторівпсихотропні засоби – фармакологічні препарати,що застосовуються при порушеннях психічної діяльності людини. Нейролептики – лікарські засоби,які використовуються для лікування психозів,головним чином шизофренії,а також інших ендогенних тп екзогенних психічнх розладів,що проявляються важкими психо-емоційними порушеннями з явищами маячення,галюцинацій,збудження. Антидепресанти — це група лікарських засобів, що виводять людину із стану депресії або субдепресії. Екзогенна або реактивна депресія розвивається внаслідок тяжкої психічної травми, соматичних захворювань, розумово-психічного стомлення, зловживання психотропними засобами, ендогенна — при психічних інфекційних захворюваннях, на фоні атеросклеротичних змін судин головного мозку. Анксиолитики (от лат. anxietas — тревожное состояние, страх + греч. lytikos — способный растворять, ослабляющий), или транквилизаторы (от лат. tranquillo — успокаивать), или атарактики (от греч. ataraxia — невозмутимость) — психотропные средства, уменьшающие выраженность или подавляющие тревогу, страх, беспокойство, эмоциональное напряжение.