

4.5.1. Гравитационная сенсорная система беспозвоночных животных
Одним из наиболее универсальных органов для восприятия гравитационного поля является статоцист, который имеется у всех типов беспозвоночных животных, за исключением класса насекомых. Статоцисты представляют собой, как правило, замкнутые полости, заполненные жидкостью и содержащие статолит, или статоконии, которые у разных видов могут быть биологического происхождения, т. е. секретироваться самим статоцистом или специальными клетками, или привнесенными из окружающей среды (например, песчинки).
Полость статоциста выстлана реснитчатыми клетками, которые у одних животных расположены равномерно, у других — образуют скопления, называемые манулами. При изменении положения тела происходит смещение статокониев в статоцисте, которое, в свою очередь, вызывает отклонение ресничек рецепторных клеток.
Статоцисты были известны морфологам еще в прошлом веке, но первоначально им приписывали слуховые функции. В 1893 г. Крейдль в своих классических опытах показал, что статоцист у омара выполняет в основном гравитационные функции. У высших ракообразных (омаров, креветок) статоцисты располагаются у основания первых антенн и открываются наружу. В качестве статолитов в них используются песчинки, которые во время линьки животных обновляются. Крейдль заменил песок в аквариуме железными опилками и показал, что с помощью сильного магнита он может заставить омаров располагаться в разных плоскостях соответственно результирующему действию магнитного и гравитационного полей. С тех пор статоцисты и их гравирецепторная функция были обнаружены во всех типах беспозвоночных (кроме насекомых).
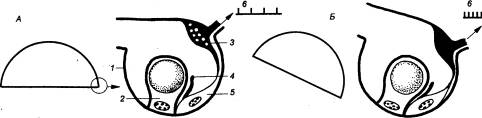
Впервые, по—видимому, подобие гравитационного рецептора намечается у одноклеточных. У инфузорий это мюллеровские пузырьки, заполненные круглыми минеральными конкрециями. У кишечнополостных (медузы) уже появляются все основные элементы сенсорной и моторной систем для оценки и удержания равновесия (рис. 4.19). По краю купола у них располагаются статоцисты, в которых статолит находится внутри специальной клетки — литоцита — и непосредственно связан с ее цитоплазмой. Рядом находится сенсорная клетка с ресничкой, которая в нормальном положении медузы не касается литоцита. При отклонении медузы от вертикали литоцит начинает давить на ресничку, заставляя ее изгибаться. Это возбуждает мембрану сенсорной клетки и приводит к возникновению РП и импульсов, которые распространяются по нервному кольцу и вызывают соответствующие рефлексы у медузы. Рецептор гравитации достигает особого развития у моллюсков (рис. 4.20).
|
Рис. 4.19 Статоцист медузы в горизонтальном положении (А) и при наклоне (В) 1 — литоцит, 2 — парус, 3 — подзонтичное нервное кольцо, 4 — чувствительный волосок (ресничка), 5 — сенсорная клетка, 6 — нейрограмма подзонтичного нерва. |
|
Рис. 4.20 Строение статоциста брюхоногого моллюска 1,8 — оболочки статоциста, 2 — центральный отросток, 3 — статический нерв, 4 — глиальная клетка, 5 — опорные эпителиальные клетки, 6 — базальная мембрана, 7 — мышечный элемент, 9 — вакуоль, 10 — статоконии, 11 — чувствительная поверхность. |

Статоцисты у моллюсков представлены парными сферическими пузырьками диаметром 50—500 мкм, заполненными вязкой жидкостью и содержащими один или много свободных статокониев. Полость статоциста выстлана сенсорным эпителием, состоящим из чувствительных реснитчатых клеток, каждая из которых окружена опорными клетками. Рецепторные клетки статоциста обычно называют волосковыми, хотя в отличие от типичных волосковых клеток позвоночных они являются первичночувствующими и имеют центральные отростки. Аксоны рецепторных клеток образуют статический нерв, оканчивающийся в церебральных ганглиях.
Статоцисты головоногих моллюсков являются наиболее высокоорганизованными и по своим функциональным возможностям, видимо, не уступают лабиринту позвоночных. Вероятно, это связано с повышением двигательной активности и, в частности, с реактивным способом передвижения этих животных.
Рецепторные элементы статоцистов головоногих моллюсков впервые среди беспозвоночных сгруппированы в те же основные структуры, что и у позвоночных, и выполняют такие же функции. Это макулы, снабженные статолитами, и кристы — группы волосковых клеток, покрытые купулами. Более подробно об их строении речь пойдет ниже, при рассмотрении лабиринта позвоночных. Здесь же лишь необходимо указать, что макулы головоногих слушав: рецепторами гравитации, т. е. обеспечивают статическое равновесие, тогда как кристы — рецепторы угловых ускорений —обеспечивают динамическое равновесие.
Таким образом, с развитием двигательной активности животных статоцист, возникший как орган чисто статического равновесия, превращается в орган, обладающий способностью регуляции не только поведения организма в гравитационном поле, но и координации его движений в трехмерном пространстве.
Система статоциста функционально связана с другими сенсорными системами, например зрительной, хеморецепторной, с системой рецепторов растяжения. Такие взаимодействия обеспечивают координацию двигательных актов, регуляцию мышечного тонуса и адаптивное поведение.
Наряду с гравитационной функцией система статоциста обладает способностью к сейсмо— и виброрецепции, а у некоторых высокоорганизованных форм и к акустическому приему. Это позволяет считать сенсорную систему статоциста ракообразных и моллюсков своего рода аналогом акустико—вестибулярных систем позвоночных животных.
У насекомых нет статоцистов. Однако чувство равновесия для насекомых чрезвычайно важно, и соответствующую информацию им поставляют наружные проприоцепторы — волосковые пластинки. Так как отдельные волосковые сенсиллы чрезвычайно чувствительны к любому механическому смещению, то действие силы тяжести на придатки тела определяет различные типы тонических реакций всего набора волосковых пластинок в теле насекомого, формируя гравитационную систему отсчета для ориентирования мышц насекомого.
У насекомых существует также механизм для восприятия вращения, т. е. получения информации об угловых ускорениях. У двукрылых насекомых для этой цели служат жужжальца. Они представляют собой два гантелевидных придатка и являются модифицированными задними крыльями. Когда насекомые летят, жужжальца быстро колеблются вверх и вниз и напоминают гироскопы, предназначенные для стабилизации самолетов и подводных лодок.