

4.4. Сенсорная система боковой линии
Круглоротые, пластиножаберные и костистые рыбы, а также некоторые водные амфибии обладают сложной системой сенсорных каналов, которые располагаются по бокам тела и часто заходят на голову животного (рис. 4.16). Каждый канал представляет собой либо открытый желобок, либо закрытый по всей длине, но имеющий несколько отдельных выходов. Органы боковой линии расположены в коже и подкожных структурах, поэтому их можно отнести к кожной сенсорной системе.
Давно известно, что органы боковой линии выполняют четко выраженную механорецепторную функцию. Адекватным стимулом для этой системы являются смещения водной среды, возникающие при разного рода ее колебаниях, вибрациях и действии низкочастотных звуковых волн.
Однако в конце 50—х — начале 60—х гг. XX в. было установлено, что в составе боковой линии некоторых рыб наряду с механорецепторными структурами существуют и иные сенсорные образования, предназначенные для восприятия слабых электрических полей водной среды. Остановимся вначале на строении и функциях обычной системы боковой линии, выполняющей механорецепцию.
|
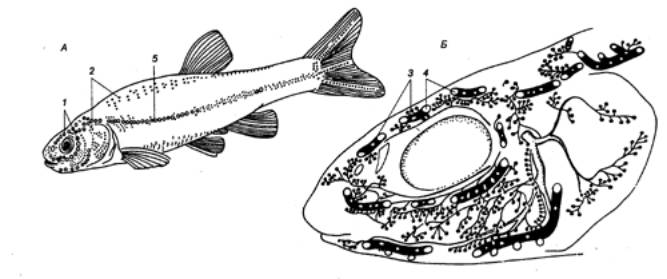
Рис. 4.16 Положение некоторых сенсорных структур на теле (А) и голове (Б) гольяна 1 — канальные поры, 2 — свободные невромасты, 3 — чувствительные каналы, 4 — поры, 5 — боковая линия на теле рыбы. |
4.4.1. Механорецепторные органы
Основной структурной единицей боковой линии является одиночный эпидермальный орган — невромаст. Невромасты могут располагаться на поверхности тела у водных амфибий и рыб (свободные) или внутри системы каналов боковой линии у большинства рыб (канальные). Каналы заполнены жидкостью или слизистым содержимым и контактируют с внешней средой через поры, закрытые у некоторых видов мембраной.
|
Рис. 4.17 Свободный невромаст у личинки костистой рыбы (разрез) 1 — опорная клетка. 2 — сенсорная клетка, 3 — купула, 4— киноцилия, 5 — стереоцилии, 6 — мантийная клетка, 7 — эфферентное нервное окончание, 8 — афферентное нервное окончание. |

Каждый невромаст содержит группу волосковых клеток, расположенных среди опорных клеток (рис. 4.17). На апикальной поверхности волосковой клетки находится набор сенсорных волосков (цилий) — 30—40 стереоцилий и единичная эксцентрично расположенная киноцилия. На противоположной, базальной поверхности волосковая клетка образует синаптические контакты с нервными окончаниями, которые могут быть афферентными и эфферентными, что позволяет рассматривать эти клетки как типичные вторичночувствующие рецепторные образования. С поверхности волосковых клеток цилии вдаются в купулу из желатинообразного слизистого вещества, которая лежит над сенсорной областью каждого невромаста. Купулы свободных невромастов, например у шпорцевой лягушки, часто имеют удлиненную форму и выдаются в водную среду, купулы канальных невромастов с овальными дискообразными сенсорными полями, как у налима, часто представлены сводчатой формой.
Таким образом, у свободных невромастов вся вспомогательная структура рецепторного аппарата ограничена купулой, тогда как у канальных между внешним стимулом и рецептирующей волосковой клеткой могут находиться не только вещество купулы, но еще и структуры, образующие стенку канала, а также жидкость, заполняющая этот канал с повышенным содержанием калия. Слизистые купулы содержат сеть микрофибрилл и прикрепляются ими к цилии волосковой клетки.
Ответ волосковой клетки вызывается усилением сдвига, создаваемого движением основания купулы, где расположены цилии. Микрофибриллы, связанные с цилиями, передают этим сенсорным структурам информацию о степени смещения. В результате возникающее движение вызывает градуальный потенциал волосковой клетки.
Свободные невромасты рыб и амфибий стимулируются преимущественно струями воды, омывающими поверхность тела. Канальные невромасты в большей или меньшей степени изолированы от этих раздражении и восприимчивы к гидродинамическим полям и инфразвуковым вибрациям. Установлено, что волосковые клетки невромастов реагируют на смещение водных частиц, вызываемое близлежащими источниками колебаний. У некоторых видов они также проявляют высокую чувствительность к температуре и солености воды.