

7.2.4. Динамические классификации
В основе динамических классификаций лежит представление о том, что каждый фитоценоз является определенным этапом в ряду последовательных сукцессии растительного покрова на конкретном однородном участке земной поверхности. Идея динамического критерия классификации растительных сообществ была обоснована Ф. Клементсом при разработке концепции климакса. Исходя из концепции климакса, Ф. Клементс составил сложную иерархическую систему классификационных единиц раздельно для климаксовой растительности, для серийной растительности и для растительности серулей - миниатюрных сукцессии, протекающих, например, на гниющих деревьях, на коре и т.п. Эта классификация была долгое время популярной в Америке, но в последние десятилетия потеряла свое значение.
Из отечественных динамических классификаций в первую очередь нужно отметить классификацию лесов Б.П. Колесникова (!956, 1967 и др.), основанную на учете выдвинутого Б.А. Ивашкевичем (1933) принципа - учета процесса возникновения и развития лесных сообществ. Сторонники данной классификации смотрят на лес, как на непрерывный лесообразовательный процесс, отдельные этапы которого принимаются за единицы классификации. Понятие лесообразовательного процесса аналогично понятию почвообразовательного процесса. Оно имеет общий характер и приложимо как к лесному покрову более или менее крупной территории, вплоть до лесного покрова всего 'Земного шара, так и к каждому конкретному участку леса. Лесообразовательный процесс охватывает все виды необратимой динамики леса - от фитоценогенеза до возрастных изменений. Однако в данной классификации в первую очередь учитываются формы динамики леса, совершающиеся в современную эпоху, в течение двух-трех столетий, а именно: возрастные, восстановительные, аллювиагенные и т.п. сравнительно быстро текущие смены.
По мнению Б.П. Колесникова (1956, 1958), ведущим факторами лесообразовательного процесса являются факторы географической среды (климат, рельеф, почвенно-грунтовые условия) и древостой - его состав, структура и производительность Наиболее важными из них будут рельеф, главная древесная порода и ее производительность, выражаемая бонитетом. При этом под главной древесной породой понимается не преобладающая численно в конкретный момент порода, а порода, способная обеспечить себе господство при достижении ею возраста спелости.
Поэтому в качестве главных критериев выделения основной единицы классификации, типа леса, принимаются рельеф, главная древесная порода и ее бонитет. Б.П. Колесников (1956) дает следующее определение типа леса: " Тип леса является качественно обособленным звеном в генетическом ряду развития лесной растительности определенной лесорастительной (ландшафтно-географической) области, этапом присущего ей лесообразовательного процесса. К одному типу леса относятся участки леса, принадлежащие к различным стадиям возрастных и коротковосстановительных смен, свойственных данному типу условий местопроизрастания и характеризующиеся общностью главной породы, а также других пород, закономерно сопутствующих главной на всех стадиях указанных смен".
Кроме типа леса, выделяются еще две важные единицы классификации; 1) тип насаждений, соответствующий возрастным и восстановительным стадиям типа леса; 2) формация, которая объединяет все типы леса с одной и той же главной породой, т.е. с одной и той же лесообразовательной породой в климаксовой стадии (Александрова, 1969). Выделяется также ряд вспомогательных единиц (рис. 34); а) зональный комплекс лесных формаций, который объединяет коренные и производные формации в пределах одной зоны, характеризующейся сходством своего исторического развития; б) климатическая фация, объединяющая типы леса какой-либо формации в пределах климатически однородного района; в) геоморфологический комплекс, включающий типы леса, связанные с одним типом рельефа; г) группа типов леса, объединяющая типы леса по принадлежности их к определенному элементу рельефа; д) лесной участок (насаждение), соответствующий лесному фитоценозу или биогеоценозу. Причем каждая из этих единиц понимается как одна из стадий, фаз, этапов единого лесообразовательного процесса.
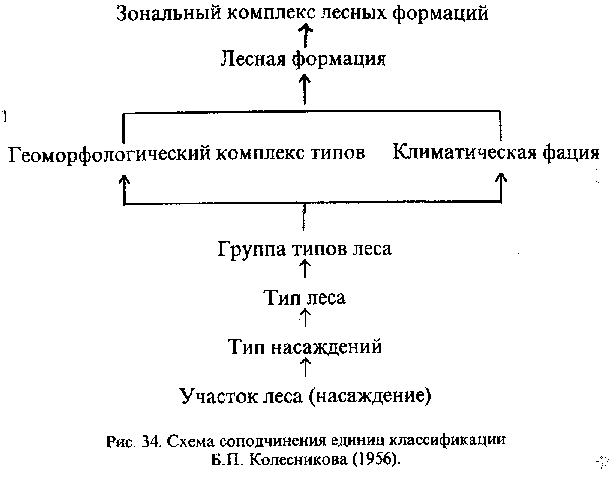
Для уяснения объема единиц классификации разных рангов приведем фрагмент классификации кедровых лесов Приморского края (Колесников, 1956);
Зональный комплекс лесных формаций - леса маньчжурского типа
Формация- кедровые леса (Приморского края)
Климатическая фация - типичные кедровники (кедрово-широколиственные леса)
Геоморфологическим комплекс - горные кедровники:
Группа типов леса I. Сухие кедровники с дубом на фрагментарных элювиальных почвах сильно ипсолируемых местообитаний:
Типы леса: 1. Рододендровые кедровники с дубом по скалистым вершинам гор и очень крутым склонам с россыпями и скалами, бонитет V-Va; 2. Леспедециевые кедровники с дубом по крутым склонам близ вершин гор, бонитет IV-V;
Группа типов леса. II. Периодически сухие кедровники с дубом на маломощных деллювиально-элювиальных почвах сильно инсолируемых местообитаний:
Типы леса; 3. Рододендрово-лещиновые кедровники с дубом по широким седловинам гребней водоразделов и среднскрутым склонам вблизи их, бонитет III-IV и т.д.
Особый подход в построении динамических классификаций связан с объединением элементарных единиц растительности (ассоциаций, групп ассоциаций) в обобщенные сукцессионные ряды, которые называют эколого-динамическим, эколого-генетическими, эволюционными рядами, или рядами развития Данные ряды затем систематизируют по факторам, определяющим сукцессионные процессы, различая, например, ряды демутационные, денудационные, гидрогенные, галогенные аллювиагенные и т.д. Впервые такой подход в нашей стране был применен М.И. Крашенинниковым (1922) при изучении растительности речных долин, а позднее широко использовался представителями школы В.Б. Сочавы (Сочава, 1960, 1962; Ильина 1968; Липатова, 1971 и др.), а также другими геоботаниками (Куваев, 1968; Тимофеев, 1970 и др.).
Ниже приводится фрагмент динамической классификации растительности южной части Якутии, разработанной В.Б. Куваевым (1968). В качестве низших единиц автор использует тип фитоценозов и группу типов, понимание которых в целом соответствует эколого-физиономической трактовке ассоциации и группы ассоциаций. Типы и группы типов объединяются в эколого-генетические ряды, для которых дается следующее определение (Куваев, 1968): " Ряд - это обобщенная последовательность сменяющих друг друга на местности в определенной ландшафтной единице типов, закономерная, поскольку она обычно отражает хронологические этапы идущего в этой единице природного процесса, неотъемлемого от эволюции ландшафта в целом. Типы в ряду преемственны: каждому из них предшествуют и наследуют определенные для данного ряда типы. Ряд принимается как единица, следующая в классификации за группой типов (табл. 20).
Различные ряды закономерно сочетаются на местности, образуя определенный тип ландшафта, который представляет собой следующую за эколого-генетическим рядом единицу классификации. Для территории южной Якутии автор выделяет два типа ландшафта: 1) аласно-таежный тип ландшафта центральной части Лено-Вилюйской равнины, 2) мелкодолинно-таежный тип ландшафта периферической части Лено-Вилюйской равнины.
Эколого-динамические классификации используются при изучении динамических тенденций в растительном покрове и позволяют отражать на геоботанических картах динамические признаки растительности.
Заканчивая обзор доминантных классификаций, нужно отметить, что в геоботанике уже давно обращалось внимание на недостатки доминантов, как критерия классификации растительных сообществ. Во-первых, многие доминанты характеризуются широкими экологическими амплитудами, и потому выделяемые по доминантному критерию единицы растительности часто являются экологически неоднородными. Особенно это касается формаций, которые нередко оказываются настолько экологически аморфными, что разные части их относятся не только к разным группам и классам формаций, но и к разным типам растительности, что нарушает соподчиненность высших синтаксонов. Так, например, разные сообщества тростниковой формации входят в состав болотистых лугов, травяных болот и прибрежно-водной растительности; основная часть сосновой формации относится к лесному типу растительности, а другая часть - к болотному типу и т.д.
Таблица 20
Фрагмент системы эколого-генетических рядов южной части Якутии (Куваев, 1968)
Зколого-генетические ряды- |
Важнейшие группы типов |
I. Денудационный (элювиально- деллювиальный водораздельный) |
1-сосновые боры, 2 - лиственничные боры, 3 - березняки чаранов, 4 - ковыльные степи чаранов, 5 -осочково-типчаковая степь чаранов, 6 - равнинная лиственничная тайга, 7 - равнинная лиственничная тайга с багульником |
11. Озерно-аллювиальный |
1 - пояс погруженных и плавающих гидрофитов, 2 -пояс обводненной растительности, 3 - пояс болотной растительности, 4 - галофитно-травяной пояс, 5- пояс кустарниковой растительности на торфяниках |
III. Аласный |
1 - пояс гидрофитной растительности, 2 - солончаковатые луга, 3 - солонцово-солончаковая растительность, 4 - солонцеватые лугостепи |
IV. Дефляционный |
1 - барханы и дюны, 2 - кедровники на барханах и дюнах, 3 - сосновые редколесья и боры |
Во-вторых, многие доминанты, особенно в травяных сообществах, неустойчивы во времени и в пространстве. С одной стороны, они могут замещать друг друга по годам в пределах одного и того же фитоценоза в результате его флюктуацией но и изменчивости и образуют сменнодоминантные комплексы (Миркин, 1974). С другой стороны, доминанты могут замещать друг друга в сходных экологических условиях в разных фитоценоза*, образуя свиты замещения (Ниценко, 1955). Кроме того, в связи с экологической индивидуальностью видов, комбинации доминантов в природе, особенно в травяных сообществах, оказываются относительно независимыми и потому разнообразными и многочисленными. В результате при ортодоксальном доминантном подходе выделяется огромное число узких по объему и неустойчивых ассоциаций, представляющих собой временные сочетания доминантов.
Поэтому некоторые геоботаники (Brockmann-Jerosch, 1907; Braun-Blanquet, 1913; Раменский, 1924) уже на ранних этапах разрешения проблемы классификации растительности стали отказываться от доминантного критерия и разрабатывать новую теорию классификации, которая получила название концепции детерминантов и на основе которой была создана флористическая классификация.