

Общая, историческая и региоанальная геология
.pdfvk.com/club152685050 | vk.com/id446425943
моллюски, а именно: наутилоидеи (ранний палеозой), аммоноидеи (силур-мел), белемноидеи (юра-мел); двустворчатые моллюски (палеоген, неоген, антропоген); граптолиты (ордовик-силур); гастроподы (палеоген, неоген, антропоген). Благодаря своей хорошей сохранности эти фоссилии играют важную роль при расчленении морских отложений.
При стратиграфическом расчленении континентальных толщ роль руководящих форм принадлежит остаткам позвоночных животных рептилий (мезозой) и млекопитающих (палеогеновый, неогеновый и антропогеновый периоды).
Большее значение в палеонтологической практике, особенно при расчленении нефтегазоносных толщ, имеют мелкие фоссилии (микрофоссилии): фораминиферы, остракоды, конодонты. В лимнических осадках, таких как диатомит, руководящая роль принадлежит диатомеям – кремниевым водорослям. В угленосных, соляных и торфяных отложениях в качестве руководящих форм выступают растительные остатки: пыльца, споры, кутикула, остатки древесины.
4.3.2. Условия существования беспозвоночных
Внастоящее время известно около 20 типов и 70 классов животных. Из них представители 57 классов обитают в морях, 10 классов – в морях и на суше, 3 класса – только на суше. Для решения задач стратификации осадочных образований наиболее интересными являются водные беспозвоночные, поскольку именно они наиболее хорошо сохранятся в ископаемом состоянии. Это связано с тем, что хранилищем остатков прежде живших организмов служит морское дно. Трупы животных и растений отлагаются на дне, мягкие части тел сгнивают, а твердые покрываются морским илом и песком и сохраняются до тех пор, пока морское дно не станет сушей и слои осадков не выйдут из-под уровня моря.
По отношению к условиям обитания выделяют две группы организмов: приспособленные к широкому диапазону колебаний факторов среды – эврибионты – и способные существовать в очень узком диапазоне изменений условий среды – стенобионты. В зависимосит от факторов среды они могут быть стеногалинные и эвригалинные (фактор среды – соленость), эвритермные
истенотермные (фактор среды – температура) и пр.
Взависимости от условий обитания в морских бассейнах организмы делятся на: бентосные (обитающие на дне), пелагические пассивно (планктон) и активно (нектон) плавающие.
движения, вызванные процессами, происходящими в ее недрах (конвективные течения в мантии, стремление литосферы к гравитационному равновесию и т. д.).
Тектонические движения в течение всего известного нам времени происходили практически непрерывно, но с различной скоростью и интенсивностью. Каждая активизация тектономагматической активности сопровождалась возникновением и развитием геосинклиналей и завершалась их закрытием с образованием жестких консолидированных участков – континентов. В геологической истории, учитывая эту повторяемость событий, может быть выделен ряд естественно-исторических этапов, получивших название этапов тектогенеза. Рубежами, разделяющими эти этапы, являются тектономагматические эпохи закрытия геосинклинальных структур или платформообразующие
эпохи складчатости.
Этап тектогенеза – это периодически повторяющиеся тектонические процессы, каждый из которых начинается геосинклинальной стадией и заканчивается послеорогенной. Этап тектогенеза включает время заложения и развития геосинклиналей и время орогенеза (горообразования).
Эпоха складчатости – совокупность фаз складчатости, приводящих к закрытию (отмиранию) геосинклинальных систем на больших пространствах земного шара.
Фаза складчатости (орогенная фаза) – кратковременные, но интенсивные импульсы тектонических движений, своего рода вспышки, создающие наибольшие качественные изменения тектонической структуры.
4.9.1 История развития Земли в докембрии
История Земли в целом может быть разделена на догеологическую и геологическую. Догеологическая история (доархейский этап) начинается с момента возникновения протопланет, в том числе и Протоземли, и исчисляется примерно в 4,5-5,0 млрд лет. Протоземля – это сгусток холодного космического вещества, который со временем уплотнялся и постепенно разогревался за счет аккреции, радиоактивного распада и приливных сил [9]. Разогрев вещества способствовал его первичной дифференциации: вода, водород, углекислый газ и др. газы, а также смеси, состоящие из легкоплавких компонентов (SiO2 ,Al2O3, CaO, Na2O, K2O, MgO и др.), радиоактивные элементы начали подниматься в верхние слои Земли – сформировалась первичная оболочка базальтоидного состава – протокора. Со временем на ее поверхности возникли вулканоплутонические кольцевые структуры, мелкие и крупные метеоритные кратеры. Пейзаж напоминал современную панораму Луны, поэтому данный период по
164 |
189 |
vk.com/club152685050 | vk.com/id446425943
Конец неогена ознаменовался неоднократным похолоданием, которые привели к возникновению материковых ледников, изменению климата, фауны и флоры. К концу среднечетвертичного времени широко распространилась хо-
лоднолюбивая фауна – мамонты, волосатые носороги, бизоны, северные олени,
овцебыки, зайцы и др. Но в конце позднечетвертичного времени в связи с потеплением климата некоторые названные группы вымерли.
Появление и развитие человека – важная особенность антропогенового периода.
Растительный мир кайнозоя богат и разнообразен. Огромные пространства северного и южного полушарий были покрыты лесами из покрытосеменных (цветковых) и голосеменных (хвойных). В начале палеогена наметились провинции:
•тропическая с вечнозелёными растениями: пальмами, лаврами, миртами, магнолиями;
•северная с умеренным климатом (север Европы, Америки, Азии – с листопадной растительностью);
•южная с умеренным климатом.
К концу неогена облик флоры на Земле приблизился к современному. В северных районах сформировалась тундровая растительность, хвойная тайга покрыла обширные пространства Сибири и Северной Америки, обособились области с тропической и субтропической растительностью, области с листопадной флорой.
Вчетвертичном периоде с наступлением ледников границы фитогенических областей резко изменились. Наступавшие ледники уничтожали всё на своём пути и угнетающе влияли на растительность прилегающих районов: вблизи ледников произрастали мхи, карликовые берёзки, полярные ивы; южнее простирались тундростепи с карликовыми берёзами и степной флорой (злаки, полынь). Ещё южнее располагались лесные массивы из хвойных деревьев, степи и полупустыни.
4.9 ПЕРИОДИЗАЦИЯ ГЕОЛОГИЧЕСКОЙ ИСТОРИИ ЗЕМЛИ
Историческая геология повествует о геологической истории Земли, начиная со времени возникновения земной коры и кончая современной эпохой. Эта сложная и длительная геологическая история насыщена колоссальным числом природных явлений, событий и процессов. Одним из важных факторов в развитии геологических процессов, изменяющих лик Земли, являются тектонические
4.4 КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТИПОВ ЖИВОТНЫХ И РАСТЕНИЙ
4.4.1 Царство животных
Простейшие ( P r o t o z o a ) . Простейшие – примитивно устроенные одноклеточные организмы, большей частью микроскопических размеров. Крупные размеры раковин (до 10 см) имели лишь единичные роды нуммулитид и орбитолинид, существовавших в карбоне и мелу. Тело простейших – комочек протоплазмы с ядром внутри, заключенный в твердую оболочку (раковину) или без нее. Размножаются путем распада материнской клетки на отдельные части. Большинство простейших – представители морской среды, пассивно плавающие или обитающие на дне. Некоторые виды приспособились к жизни в пресных водоемах, в почвах или паразитируют на животных. В ископаемом состоянии известно около 30 тыс. видов.
Простейшие делится на ряд типов, наиболее значимым из которых для геологии является тип саркодовых (Sarsodina), а именно: классы фораминифер
(Foraminifera, рис. 52) и радиолярий (Radiolaria, рис. 53). Представители этих классов имеют скелет в виде раковины и хорошо сохраняются в различных отложениях. В осадочном покрове Земли они распространены в отложениях всех возрастов: от кембрийских до современных. Видовые разновидности фораминифер используются для детального расчленения нефтеносных отложений.
Радиолярии известны с докембрия. В отличие от фораминифер в строении их скелета отмечается большое количество кремниевых лучей. Радиолярии – руководящие формы перми и триаса.
б
Nummulites
Ст Л
а
г
|
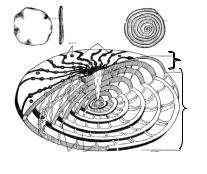
Рис. 52. Строение раковины нуммулитид: |
|
в |
а, б – внешний вид рода Nummulites; |
|
а – вид сверху; б – вид сбоку уплощенной |
||
|
Iмонетовидной раковины;
в– поперечное сечение раковины (видны многочисленные спиральные обороты);
IIг – схема строения раковины нуммулитид: I – продольное (осевое) сечение,
II – поперечное (экваториальное) сечение; Ст – столбики; Л – линии прикрепления перегородок камер [19]
188 |
165 |

vk.com/club152685050 | vk.com/id446425943
Рис 53. Разнообразные формы раковин радиолярий (увел.)
Простейшие, обладая минеральным скелетом, являются породообразующими организмами, давая материал для формирования многометровых толщ фузулиновых, нуммулитовых, швагериновых известняков, широко развитых в Альпах, на севере Африки (использовались для строительства пирамид), в Центральной Азии, Поволжье, Приуралье, Прикаспии. Руководящие формы карбона и мела.
Многоклеточные. Тело многоклеточных состоит из клеток и межклеточного вещества, образующих ткани и органы. Они делятся на низших (ненастоящих) многоклеточных и высших (настоящих) многоклеточных.
Низшие многоклеточные (Parazoa). Тело состоит из клеток, не дифференцированных на ткани и органы.
Тип губки (Spongia). Губки – наиболее примитивные одиночные или колониальные представители многоклеточных организмов (рис. 54).
|
|
|
|
|
|
|
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
|
б |
|
в |
|
|
|
|
|
|
|
|
|
|
|
гв |
|
|
|
|||
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис 54. Представители типа Spongiа:
а, б – кремниевые губки ордовикского и силурийского периодов (х 100); в, г – стеклянные (шестилучевые) губки юры и мела, высота ~ 13 см
не доминировали разнозубые. Их родовой состав к концу периода приблизился к современному.
Кайнозойские гастроподы освоили не только море, но и сушу. Из фораминифер наибольшего расцвета достигли глобигерины, роталииды, милиолиды. Теплые окраинные моря, входившие в состав океана Тетис, заселяли нуммулитиды. В эоцене они достигли своего расцвета и увеличились в размере (до 10-15 см), а в конце эоцена-олигоцена – вымерли. Лишь отдельные их виды дошли до наших дней.
Кайнозой – время расцвета радиолярий и морских ежей. Скопления сферических, дисковидных и колпачковидных скелетов на дне мелких морей привело к образованию радиоляритов. В песчаных грунтах мелководных морских бассейнов вели малоподвижный образ жизни щитовидные морские ежи, в ил зарывались сердцевидные ежи.
Вэкваториальных зонах широким развитием пользовались шестилучевые кораллы, где они вместе с мшанками и губками создавали рифовые постройки. Мощные пласты диатомита и известковых илов обязаны своим происхождени-
ем диатомовым и золотистым водорослям.
Из позвоночных в морях и пресноводных бассейнах палеогена и неогена господствовали костные рыбы, видовой состав которых постепенно приближался к современному. На суше господствовали млекопитающие, теплокровность, живорождение и достаточно высокая организация мозга которых способствовали приспособлению их к различным условиям существования на Земле. В начале палеогена они были представлены клоачными, сумчатыми и примитивными плацентарными (насекомоядные, древние копытные, приматы). Среди плацентарных во второй половине палеогена появились представители почти всех ныне существующих отрядов. Это китообразные (киты, дельфины,
тюлени, моржи, сирены), настоящие хищники (креодонты), ластоногие, хоботные, парнокопытные и непарнокопытные (лошадиные, бронтотериевые, носо-
рогообразные и пр.).
Вначале неогена началось резкое осушение климата, что повлекло за собой вымирание многих групп влаголюбивых животных. Им на смену пришли новые семейства, существующие и доныне. Приспособление к быстрому бегу и питанию жесткими травами обусловило быструю эволюцию лошадиных. В середине неогена Европу, Азию, Африку и Северную Америку заполонили трехпалые лошади (гип-
парионы), саблезубые кошки (махайродусы), гиены, жирафы, олени, носороги, свиньи, хоботные, страусы, человекообразные обезьяны. К этому же времени относитсяпоявлениенаЗемле древныхпредковчеловека.
166 |
187 |
vk.com/club152685050 | vk.com/id446425943
Втриасовый период получили широкое развитие рептилии, обитавшие на суше и в море: черепахи, крокодилы, чешуйчатые и звероподобные ящеры, те-
кодонты, динозавры. Динозавры к началу юры стали господствующими, а звероподобные ящеры дали начало млекопитающим. Юрские динозавры – ящеротазовые, меловые – птицетазовые. В среднем мелу появились змеи.
Мезозойские рептилии завоевали и воздушную среду, в начале юры появились летающие ящеры – птерозавры. Юрский период – время развития хвостатых форм, меловой – время расцвета бесхвостых ящеров с размахом крыльев до 17 м.
Среднеюрский период – время появления первых птиц. В раннем мелу поя-
вились зубастые плавающие и летающие птицы, в конце мела – веерохвостые.
Втриасовый период возникли млекопитающие. В юрский период существовали наиболее примитивные группы, в раннемеловой – клоачные и сумчатые, в позднемеловой – насекомоядные плацентарные.
Вконце триаса полностью вымерли стегоцефалы, зато появились бесхвостые амфибии. На суше господствовали различные членистоногие: пауки, клещи, насекомые. В пресных водах водились лучеперые, кистеперые и двоякодышащие рыбы, остракоды, листоногие раки (филлоподы).
Морской растительный мир – диатомовые и золотистые водоросли.
Диатомиты появились в юре, а начиная с мела, стали породообразующими. Золотистые водоросли известны в ископаемом состоянии с мела.
Из наземных растений господствовали голосеменные, папоротниковидные. Настоящие папоротники встречались редко. Со среднего мела преоблада-
ли покрытосеменные (цветковые).
Меловой период в истории Земли – период резкого изменения органического мира: голосеменные сменили покрытосеменных, вымерли аммониты, белемниты, многие роды брахиопод, динозавры, летающие ящеры, почти все морские рептилии.
4.8ОРГАНИЧЕСКИЙ МИР КАЙНОЗОЯ
Вкайнозойских морях из беспозвоночных были распространены двустворчатые и головоногие моллюски, фораминиферы, радиолярии, морские ежи, шестилучевые кораллы, губки, мшанки; из растений – диатомовые и золотистые водоросли.
Впалеогене двустворки заселяли мелководные участки дна морских бассейнов с нормальной соленостью, а также лагуны и пресные водоемы. В неоге-
Их тело представляет собой мешочек бокало-, грибоили кубковидной формы с большим количеством каналов и пор, по которым вода и пища поступают во внутреннюю полость организма.
Некоторые виды имеют форму воронки, кустика, ветвистого дерева. Стенки мешочка поддерживаются органическим или минеральным скелетом, состоящим из множества одно-, трех-, четырехили шестиосных элементов – спикул. Губки – морские организмы – прикрепленный бентос, и лишь отдельные виды приспособились к жизни в пресной воде. Известны с венда. Руководящее значение имеют для меловой и палеогеновой систем. В мезозое этот тип имел массовое распространение и выступал в качестве рифостроящих организмов. Спонголиты – породы, состоящие из спикул губок.
Тип археоциаты (Archaeoceatha). Вымершие одиночные и колониальные животные. Их известковый скелет состоит из двух стенок, напоминающих два вложенных один в другой кубка или цилиндра, поэтому они и называются «древними кубками». Между стенками располагаются радиальные и горизонтальные перегородки – опорные септы. Стенки полностью или частично перфорированы. Через поры центральной полости внутрь тела проникала вода с пищей, а затем выходила наружу через большое отверстие.
Археоциаты вели прикрепленный, бентосный образ жизни в мелководных теплых морях и участвовали в формировании органогенных построек. В истории Земли они являются первыми рифостроителями. Известны в отложениях нижнего кембрия, в среднем кембрии обнаружены лишь отдельные представители.
Высшие многоклеточные (Eumetarazoa). Тело состоит из клеток,
дифференцированных на ткани и органы.
Тип стрекающие (Cnidaria). Низкоорганизованные морские животные, по форме напоминающие губки. Имеют органы чувств, мускульную систему, нервные и стрекательные клетки, служащие для защиты от врагов и умерщвления мелких животных, которыми они питаются. Кровеносная система отсутствует. Пищеварительная полость открывается наружу ротовым отверстием, которое служит для введения пищи и выведения отбросов. Дышат всей поверхностью тела. В составе стрекающих выделяется 4 класса, в ископаемом состоянии сохраняются только некоторые гидроидные и коралловые полипы.
Из ископаемых гидроидных наиболее распространены строматопораты, составляющие вымершую группу стрекающих. Они образовывали массивные, клубко- и клинообразные колонии, прираставшие к субстрату. Особи выделяли известковый скелет с параллельными наружной поверхности волнистыми ла-
186 |
167 |
vk.com/club152685050 | vk.com/id446425943
меллями и лежащими между ними вертикальными опорными элементами – столбиками. Как рифообразующие организмы строматопораты существовали от кембрия до мела. Максимального расцвета достигли в силурийском и девонском периодах.
Коралловые полипы или «животные-цветы» с их массивными скелетными образованиями относятся к важнейшим фоссилиям. Они встречаются в отложениях всех систем. Предпочитают теплые (не ниже 20оС) чистые моря с нормальной соленостью, насыщенные кислородом и обильным планктоном (продукт питания кораллов). За время своего существования дважды пережили расцвет: в силурийский-пермский и юрский-современный периоды. Преобладают колониальные формы, в которых известковый скелет одной особи (полипа) получил название кораллита, а скелет колонии, образованный совокупностью кораллитов, – полипняка. Внутри кораллита имеются перегородки: мягкие радиальные – септы и горизонтальные – днища. Посередине располагается известковый выступ – столбик.
Класс коралловых полипов подразделяется на подклассы, из которых наибольшее стратиграфическое значение имеют табуляты (Tabulatomorpha) и четырехучевые (Tetracorallia, рис. 55) кораллы. В современных морских бассейнах обитают шестилучевые (Hexacorallia) и восьмилучевые (Octocorallia) кораллы (рис. 56). Днем они обычно сжимаются, а ночью вытягиваются и расправляют щупальца, чтобы ловить мелкий планктон. Часть видов кораллов живут за счет симбиоза с одноклеточными водорослями. Колониальные кораллы участвуют в образовании органогенных построек и коралловых рифов, поэтому кроме стратиграфического значения имеют важное породообразующее значение.
аб
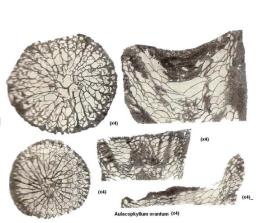
Рис. 55. Представитель рода
Aulacophyllum orantum (подкласс Tetracorallia), сирачойская свита,
Ухтинский район, Южный Тиман: а – поперечный разрез кораллита; б – продольный разрез кораллита
Подобная зональность сохранялась до конца перми. В конце перми климат стал наиболее засушливым. В первой климатической области вместо влаголюбивых растений появились различные голосеменные, цикадовые, гинкговые. Климатические изменения во второй и третьей областях произошли значительно позднее.
4.7 ОРГАНИЧЕСКИЙ МИР МЕЗОЗОЯ
Рубеж между пермским и триасовым периодами характеризуется значительным изменением органического мира и, прежде всего, животных. Полностью или почти полностью закончили свое развитие фузулиниды, табуляты, гониатиты, трилобиты, брахиоподы, некоторые амфибии и рептилии. Появились новые группы цефалопод, двустворок, гастропод, иглокожих, мелких фораминифер, кораллов, рыбиморскихрептилий, началасьперестройкавмирерастений.
Аммоноидеи – одна из самых распространенных морских групп. В триасе доминировали формы с цератитовой, а в юре – с аммонитовой лопастной линией. Аммониты мелового периода чаще имели неправильно свернутую или прямую раковину – результат смены образа жизни и условий обитания.
Белемниты максимального расцвета достигли в юрский и меловой периоды. В конце мала количество их резко сократилось, а к началу палеогена сохранились лишь единичные формы.
Из гастропод господствовали переднежаберные (мезогастроподы), среди двустворок – разнозубые, связкозубые и толстозубые (рудисты).
Четырехлучевых кораллов сменили шестилучевые. Они образовывали массивные поселения в юрских и меловых морях, слагая вместе с губками, рудистами и мшанками рифовые постройки.
Малоподвижных и прикрепленных иглокожих сменили подвижные формы. Возникли новые группы морских ежей, морские лилии приобрели способность к передвижению при помощи рук.
Вмеловой период максимального развития достигли мелкие фораминиферы. Различные карбонатные породы этой системы образованы, в основном, из их раковин и обломков раковин. Максимального расцвета достигли также
конодонты.
Вмезозое существовали, но не играли значительной роли губки, строматопораты, черви, членистоногие (мечехвостые, раки, рачки), наутилиды, мшанки, брахиоподы, морские звезды, голотурии.
Из морских позвоночных наиболее широко были распространены хрящевые (акулы и скаты) и костные (кистеперые, лучеперые, костистые) рыбы, пресмыкающиеся (завроптеригии, ихтиоптеригии, мезозавры).
168 |
185 |

vk.com/club152685050 | vk.com/id446425943
Вдевоне на континентах появились первые четырехногие позвоночные
земноводные – стегоцефалы или крышеголовые, достигшие максимального расцвета в карбоне и перми. В карбоне и перми широкое развитие получили также батрахозавры – лягушкоящеры – предки пресмыкающихся. Основные группы стегоцефалов вымерли в конце перми.
Первые пресмыкающиеся или рептилии завоевали господство на суше в конце карбона. Они потеряли связь с водной средой и проникли вглубь континентов. Размножались при помощи яиц, откладывая их прямо на суше. В карбоне и перми были широко распространены котилозавры, в перми – растительноядные и хищные парейазавры, хищные иностранцевии, мезозавры.
Из наземных беспозвоночных в девоне уже существовали паукообразные, клещи, в карбоне появились первичнобескрылые и первые древнекрылые насе-
комые с размахом крыльев до 100 см.
Континенты в позднем палеозое были покрыты пышной растительностью. В раннем и среднем девоне в болотистых местах произрастали риниофиты, со среднего девона широкое развитие получили все основные группы споровых растений – плауновидные, членистостебельные, папоротниковидные.
Позднедевонскую флору часто называют археоптерисовой – разновидность разноспоровых папоротников.
Доминантами каменноугольной флоры являются древовидные плауновидные (лепидодендроны, сигиллярии с высотой ствола до 30-40 м), членистостебельные (древовидные и кустарниковые формы с членистым строением ствола
имутовчатым расположением листьев), папоротники, птеридоспермиды, кордаитовые. Папоротниковидные размножались семенами, что и позволило им проникнуть глубоко внутрь континентов. В конце карбона появились первые гинкговые и хвойные, завоевавшие засушливые районы.
Враспределении каменноугольной флоры наметилась четкая зональность, обусловленная климатическими условиями: выделились три флористические области:
•тропическая Вестфальская или Еврамериканская, протягивающаяся от центральной части северной Америки до Юго-Восточной Азии, с плауновидными, членистостебельными, птеридоспермидами;
•умеренная Тунгусская с кордаитами, папоротниками, птеридоспермидами и травянистыми членистостебельными;
•умеренная Гондванская, занимавшая почти весь материк Гондвана с глоссоптериевой и кордаитовой листопадной флорой.
184
Рис 56. Схема строения восьмилучевых кораллов (по Л. В. Шимкевич):
1 – щупальца; 2 – ротовое отверстие;
3- глотка; 4 – септа;
5 – мезентериальные1 нити; 6 – яйца
2Колониальные кораллы участвуют в образовании органогенных построек и коралловых рифов, поэтому кроме стратиграфического значения имеют важное породообразующее значение.
Тип низшие черви (Scolecida). Объединяет большую группу низкоорганизованных червей, населяющих моря, пресные водоемы, почву, паразитирующих на теле животных и растениях. В отличие от кишечнополостных, у них имеются пищеварительный тракт, кровеносная система, примитивные органы выделения. В ископаемом состоянии известны с каменноугольного периода.
Тип высшие (кольчатые) черви (Annelida). Имеют тело, дифферен-
цированное на голову, туловище, анальную полость. У единичных родов оно заключено в известковистый скелет. Известны с венда. В Европе обнаружена порода серпулит раннемелового возраста, состоящая из ископаемых трубок червей рода Serpula.
Тип моллюски или мягкотелые (Mollusca). Чрезвычайно разнооб-
разная и многочисленная группа. Число современных видов превышает 100 тыс., столько же видов известно в ископаемом состоянии. Из современных наиболее широко распространены двустворки, брюхоногие, головоногие моллюски, из ископаемых – головоногие – аммониты и белемниты. Большинство моллюсков обитает в морских водах всех широт и на различных глубинах (от 0 до 5000 тыс. м), отдельные двустворки и брюхоногие приспособились к жизни в солоноватых и пресных водоемах, некоторые брюхоногие ведут наземный образ жизни (виноградная улитка).
1 Мезентериальные нити – нити, на которые распадаются внутренние края мезентериальных складок, делящих полость полипа на камеры. Нити выделяют вещество, близкое по составу к желудочному соку.
169

vk.com/club152685050 | vk.com/id446425943
По способу питания выделяются: растительноядные, хищные формы и падалееды. Одни активно собирают пищу, другие фильтруют воду, улавливая из нее питательные вещества; одни ведут бентосный, другие – нектонный образ жизни.
Брюхоногие (Gastropoda, gaster – «желудок», podos – «нога») моллюски – многочисленный класс, объединяющий около 100 тыс. современных и ископаемых видов. Тело брюхоногих заключено в асимметричную спиральноконическую (рис. 57), плоскоспиральную или колпачковидную раковину и делится на голову, ногу, висцеральный мешок и мантию. Мантия выделяет карбонат кальция, необходимый для постройки раковины. Поверхность раковины чаще всего осложнена различными скульптурами: ребрами, шипами, бугорками. Между мантией и находящимся внутри нее телом имеется мантийная полость, где расположены дыхательные органы. Брюхоногие, в зависимости от расположения сердца по отношению к жабрам и способа дыхания, разделены на 3 класса: переднежаберные, заднежаберные и легочные. Наибольшее стратиграфическое значение имеют переднежаберные. Известны с кембрия, максимального расцвета достигли в пермский, триасовый периоды и в кайнозое.
Рис. 57. Широко распространненые спиральноконические раковины представителей класса Gastropoda:
1 – левозавитая раковина и
2 – правозавитая раковина тропической наземной гастроподы, род Bulimus perversus (по Мартенсу)
Двустворки (Bivalvia, bi – два, valva – створка), именуемые также топороногие, пелециподы, пластинчатожаберные, являются обитателями пресных, солоноватых и соленых бассейнов. Мягкое тело животных имеет двустороннюю симметрию и покрыто двумя створками. Внутренние органы заключены в мантию, состоящую из двух лопастей – левой и правой, каждая из которых прикреплена к своей створке. След присоединения мантии к створке (мантийная линия) виден на внутренней стороне раковины. Мантийная линия проходит параллельно краю раковины, иногда образуя изгиб – мантийный синус. Тело двустворки состоит из туловища и ноги; последняя занимает срединное положение
трехзонные кораллы, у которых отмечается дальнейшее усложнение скелета – возникновение внутри кораллита известкового столбика. В раннепермскую эпоху началось угасание четырехлучевых кораллов. В девоне утратили свое былоегосподствотабуляты, акконцупермиосталисьлишьединичные виды.
Из иглокожих дно мелких морей в девоне и карбоне заселяли бластоидеи и криноидеи. Роль морских ежей стала заметна лишь во второй половине позднего палеозоя. К концу перми исчезли бластоидеи, многие группы криноидей и древние морские ежи.
Гониатиты – одна из важнейших групп, населявших морские бассейны позднего палеозоя. Первые гониатиты с плоскоспиральной раковиной и простой лопастной линией появились в девоне. В фаменском веке пышного развития и разнообразия достигли климении. В начале карбона число гониатитов резко сократилось, но в среднем и позднем карбоне их количество снова заметно возросло, появились формы с более усложненной лопастной линией и скульптурой. К концу перми гониатиты уступили место цератитам, первые представители которых появились еще в артинском веке. К концу перми исчезли наутилоидеи с прямой раковиной, их место заняли наутилоидеи с плоскоспиральной раковиной.
Широкое развитие в позднем палеозое получили бактриты – организмы с небольшой прямой или согнутой раковиной – «предки» каменноугольных белемнитов (колеоидей). Новые места обитания завоевали двустворки и гастро-
поды, продолжали существовать губки, строматопораты, мшанки. К концу перми заметно уменьшился состав остракод, а тентакулиты, ракоскорпионы,
трилобиты и граптолиты почти полностью исчезли.
В позднем палеозое дальнейшее развитие получили конодонты. В настоящее время они используются для детального расчленения и межрегиональной корреляции девонских, каменноугольных и пермских разрезов.
Большого разнообразия достигли и позвоночные. Они расселились в морских водоемах и на суше. В морях и пресноводных бассейнах девонского пе-
риода господствовали бесчелюстные рыбообразные, панцирные, костные и хрящевые рыбы. Хрящевые (древние акулы) испытали расцвет в каменноугольное время, а в перми им на смену пришли рыбы, у которых зубной аппарат представлял собой свернутую в 2-3 оборота плоскую спираль. Из костных рыб существовали кистеперые, двоякодышащие и лучеперые. Плавники вымерших кистеперых рыб обнаруживают большое сходство с конечностями первых наземных позвоночных – земноводных. Одна из групп кистеперых рыб дожила до наших дней и обитает в Индийском океане у юго-восточных берегов Африки.
170 |
183 |
vk.com/club152685050 | vk.com/id446425943
пузыри), в среднем – криноидеи (морские лилии) и бластоидеи. По дну ползали
морские звезды и правильные древние морские ежи. Силурийский период – время расцвета криноидей.
В нижнепалеозойских породах, начиная со среднекембрийских, часто встречаются конодонты. Широкое развитие червей подтверждается наличием челюстных аппаратов – сколекодонтов и обилием ходов в глинистых породах.
Наряду с беспозвоночными в морских бассейнах ордовика и силура существовали и развивались позвоночные. В породах ордовика найдены кожные зубы бесчелюстных рыбообразных организмов. В конце силура появились первые челюстные позвоночные – настоящие рыбы.
Из низших растений широким развитием пользовались сине-зеленые водоросли, известные нам по сохранившимся в породах строматолитам и онколитам, красные, бурые и зеленые водоросли. В конце силура на суше появились высшие растения – риниофиты, подготовившие выход на сушу животных.
4.6ОРГАНИЧЕСКИЙ МИР ПОЗДНЕГО ПАЛЕОЗОЯ
Кначалу позднего палеозоя резко уменьшилось разнообразие трилобитов, граптолитов, иглокожих, табулят и наутилоидей. Появились новые группы животных и растений, обитавших не только в морях, но и на суше, определивших облик органического мира позднего палеозоя. Из беспозвоночных особенно широкого развития достигли: замковые брахиоподы, фораминиферы, четырехлучевые кораллы, гониатиты, иглокожие (криноидеи) и мшанки.
Замковые брахиоподы максимального расцвета достигли в девоне. Характеризовались богатством видов, быстрой изменчивостью во времени, что и позволило использовать их в качестве важнейшей группы для определения возраста пород. В каменноугольном периоде количество их сократилось, но все же они остались одной из доминирующих групп. По обилию продуктид, раковины которых достигали до 40 см ширины, отложения этого возраста часто называют «продуктусовыми». В пермском периоде закончили свое существование представители многих групп замковых брахиопод, поэтому граница перми и триаса легко устанавливается по этому сокращению.
Фораминиферы обитали на дне мелководных теплых морей, где образовывали массовые скопления и являлись породообразующими организмами. В девоне широкое развитие получили спирально-свернутые многокамерные формы – эндотириды, в карбоне – фузулиниды, в перми – швагериниды.
В девонском периоде дальнейшее развитие получили двузонные кораллы как одиночные, так и колониальные. В каменноугольном периоде появились
и имеет языковидную или клино- и топорообразную форму. С обеих сторон от нее свисают жабры.
На спинном (верхнем) крае обеих створок находятся макушки, с внутренней стороны которых имеются известковые отростки (зубы), разделенные межзубными ямками. В совокупности они образуют зубной (замочный) аппарат. Иногда вместо зубов присутствует связка. Замыкаются створки при помощи мускул. На внутренней стороне створок часто можно наблюдать мускульные отпечатки. Классификация двустворок осуществляется на основании строения замка.
Двустворки известны с кембрия. Наибольшее значение имеют для расчленения мезозойских и кайнозойских отложений (мел, палеоген, неоген).
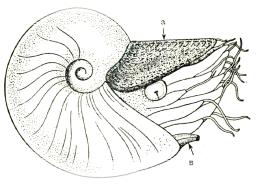
Головоногие (Cephalopoda, kephale – голова, podos – нога) – высокоразви-
тые морские организмы, получившие название «приматы моря». К ним относятся: каракатицы, осьминоги, кальмары, наутилиды, вымершие аммониты и белемниты и т. д. Размеры их зачастую бывают фантастические и достигают до 6,5 м в длину и 1,95 м в поперечнике. Общее количество видов – более 10 тыс., в настоящее время обитает поряка 650 видов.
Головоногие характеризуются обособленной головой (рис. 58), высоко развитыми органами чувств и совершенными глазами. На голове моллюска располагаются многочисленные «руки» (щупальцы), служащие для захвата пищи и частично для передвижения. Число щупальцев может достигать 100. Основным средством передвижения является воронка – трубчатый мускульный орган, сообщающийся с жабрами.
а
Рис. 58. Раковина аммонита с открытым устьем
(Schindewolf, 1958);
а – аптих; в – воронка
Важной частью мягкого тела является мантия – кожистая складка, покрывающая туловище. С помощью залегающих в кайме мантии желез животное строит раковину.
182 |
171 |
vk.com/club152685050 | vk.com/id446425943
Класс делится на шесть подклассов, пять из которых объединены в группу наружнораковинных, а шестой является внутреннераковинным.
Наружнораковинные имеют форму прямых, согнутых или спирально закрученных конических трубок, разделенных поперечными перегородками на камеры. Мягкое тело моллюска расположено в передней – жилой – камере. Остальные камеры заполнены газом и называются фрагмакон. Они помогают животному удержаться на воде. Во всех перегородках имеется отверстие, через которое проходит кожистая трубка – сифон. След прикрепления перегородок к стенкам раковины – перегородочная (лопастная) линия.
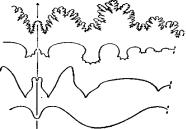
На основании строения лопастной линии, которая бывает четырех типов (рис. 59), выделяют наутилид, цератитов, гониатитов, бактритов, аммоноидей. Наутилоидеи и цератиты достигли своего расцвета в раннем палеозое, гониатиты – в позднем палеозое; мезозой – эпоха расцвета цератитов и аммонитов.
Внутреннераковинные моллюски имеют известковую или роговую внутреннюю раковину, лежащую внутри висцерального мешка. Выделяют четыре отряда: каракатицы, кальмары, осьминоги, белемниты. Белемниты, они же «громовые стрелы», «фигурные камни» – вымершие организмы, являющиеся важными руководящими формами юры и мела. В задней части тела они имели разделенную на камеры раковину – фрагмакон, к которой примыкал конический, цилиндрический или сигарообразный ростр (в народе – «чертов палец») из битуминозного радиально-лучистого кальцита. При трении ростр издавал своеобразное зловоние, напоминавшее запах кошачьей мочи, за что получил еще название «кошачий камень». Благодаря особенностям кальцитового состава, ростры очень устойчивы к выветриванию и в массовом количестве встречаются в ископаемом состоянии, образуя так называемые «поля битв белемнитов».
А) аммонитовый тип;
Б) цератитовый тип;
В) гониатитовый тип;
Г) агониатитовый тип
Рис. 59. Перегородочные линии головоногих моллюсков
Трилобиты – важнейшая в стратиграфическом отношении группа раннепалеозойских животных. Малочленистые трилобиты (миомеры) испытали расцвет в кембрии и закончили свое развитие в ордовике.
Расцвет многочленистых трилобитов (полимеров) также приходится на кембрийский период; именно в кембрийских отложениях находят остатки большинства известных родов. Начиная с позднего кембрия, полимеры постепенно теряют завоеванные ими позиции, но при этом обнаруживают прогрессивное непрерывное развитие – удивительное разнообразие их строения и внешнего облика.
Граптолиты появились в кембрии, максимального расцвета достигли в ордовике и силуре. В раннем и среднем кембрии преобладали стереостолонаты – бентосные колониальные формы, в конце кембрия появились планктонные и псевдопланктонные мономорфные колонии. В раннем ордовике получили расцвет безосые граптолиты, во второй половине ордовика – осеносные с двурядным расположением тек, в силуре – однорядные осеносные формы.
Брахиоподы обитали на мелководье. В раннем кембрии существовали исключительно беззамковые формы, в конце кембрия появились замковые брахиоподы, достигшие значительного развития в ордовике и силуре.
Головоногие моллюски – первые морские хищники – вели подвижный образ жизни. Кембрийские головоногие представлены мелкими формами, ордовикские наутилоидеи и цератитиды достигали в длину нескольких метров. Их раковина имела форму бочонка, длинной палки, рога, плоской спирали.
Кишечнополостные представлены гидроидными (строматопораты), коралловыми полипами и медузами. Строматопораты в течение ордовика и силура строили крупные колонии и наравне с другими организмами участвовали в образовании рифовых построек. Из коралловых полипов наиболее широко были распространены табуляты. Первые табуляты появились в кембрии, в ордовике широкого расцвета достигли формы с несообщающимися между собой кораллитами. В силуре к ним присоединились группы табулят с сообщающимися при помощи пор кораллитами. К концу силура табуляты достигли исключительного расцвета и получили широкое распространение во всех морях. В конце ордовика появились первые четырехлучевые кораллы. Это были простые однозонные формы. В силуре они поднялись на более высокую эволюционную ступень: развитие получили двузонные кораллы.
Важное место на морском дне занимали иглокожие. В кембрии преобладали капроидеи; в раннем ордовике к ним присоединились цистоидеи (морские
172 |
181 |
vk.com/club152685050 | vk.com/id446425943
Покрытосеменные (цветковые) – наиболее высокоорганизованные растения. Они имеют семена, которые образуются внутри завязи цветка. В последствии стенки завязи превращаются в плод. К цветковым относятся пальма, дуб, береза, ясень, клен и т.д. Время расцвета – кайнозой.
Членистостебельные широко были распространены в палеозое. Они имели полый внутри членистый стебель. В карбоне и перми членистостебельные были крупнейшими древесными растениями, известными под названием каламитов. Их стволы достигали 30 м в высоту и 1 м в поперечнике. В настоящее время это исключительно травянистые растения (32 вида хвощей).
Особенно большое значение имеют семена, споры, пыльца растений, позволяющие расчленять континентальные толщи, которые практически не содержат ископаемых остатков животных. На изучении спор и пыльцы растений основан метод определения возраста континентальных отложений, получивший название спорово-пыльцевого метода.
4.5 ОРГАНИЧЕСКИЙ МИР РАННЕГО ПАЛЕОЗОЯ
Органический мир раннего палеозоя характеризуется развитием животных, низших и высших растений. Важной особенностью органического мира является способность организмов строить разные по химическому составу наружные или внутренние скелеты. Эта особенность может быть объяснена, вопервых, появлением хищников (скелет – защитная реакция организмов), вовторых, изменением химического состава вод океанов и, в-третьих, переходом от планктонного (в докембрии) образа жизни к бентосному (скелет – опора организму в зоне подвижных вод).
Особенно широко в раннепалеозойских морях были развиты археоциаты, трилобиты, граптолиты, головоногие моллюски, коралловые полипы и иглокожие. Возможно, в водах обитали и другие беспозвоночные, но у одних групп остатки не сохранились (черви), а другие только начали свое историческое развитие и не сыграли существенной роли. Из этих групп можно назвать двустворок, гастропод, остракод, тентакулитов, мшанок, ракоскорпионов и водных скорпионов. Общий специфический облик раннепалеозойского органического мира морских беспозвоночных дополняли появившиеся еще в протерозое простейшие – фораминиферы, радиолярии, губки.
Археоциаты населяли теплые мелководные моря. Часто они сплошным ковром покрывали дно и принимали участие в построении рифов. Максимального расцвета достигли в раннем кембрии, к концу кембрия вымерли.
Тип членистоногие (Arthropoda). Данный тип объединяет примерно ¾ видов всего животного царства. Это насекомые, раки, пауки, крабы, скорпионы, клещи, вымершие трилобиты и т. д. – организмы, тело которых состоит из большого количества члеников или сегментов. Тип разделен на 4 подтипа: трилобитообразные, жабродышащие, хелицеровые и трахейные.
В подтипе трилобитообразных наиболее изучены трилобиты – вымершие животные, обитавшие на морском дне. Массовое распространение они имели в раннем палеозое. Хитиново-известковый панцирь трилобитов разделен на 3 части (рис. 60): головной щит, туловищный отдел и хвостовой щит. Туловищный отдел состоит из сегментов, число которых у разных видов изменяется от 2 до 44. Каждый сегмент снабжен парой конечностей. Конечности (4-5 пар) располагаются и на нижней стороне головного щита. Трилобиты с количеством сегментов 2-4 имеют размеры 1-2 см и относятся к подклассу малочленистых (Miomera). К многочленистым (Polimera) относятся трилобиты с числом сегментов 5-44, их длина может достигать 75 см. Руководящие формы кембрия, ордовика, силура.
а 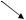
б
в
Рис. 60. Руководящие формы трилобитов песчано-известковых фаций мелководья (по Г. Крумбигель и Х. Вальтер):
1 – Asaphus expansus; 2 – Chasmops odin.; 3 – Megistaspis limbata; 4 – Cryptolithus goldfussi; 5 – Niobe sp.;
а– головной щит; б – туловищный отдел; в – хвостовой отдел
Кподтипу жабродышащих относят класс ракообразных (Crustacea), из которых наибольшее значение имеют отракоды (Ostracoda, рис. 61) – ракушковые рачки. Их тело заключено в раковину, поверхность которой либо гладкая, либо скульптированная, покрытая бугорками, шипами, ребрами. На голове располагаются 3 глаза, 2 пары конечностей, антенны и антеннулы. Тело несет от 1 до 3 пар конечностей.
180 |
173 |