

Введение в психологию_Аткинсон, Смит, Бем и др_2003 -713с
.pdfРис. 4.19. Тестирование цветовой слепоты. В тесте на цветовую слепоту используются две картинки. На левой картинке некоторые индивиды, страдающие определенными видами краснозеленой слепоты, увидят только цифру 5, некоторые — только цифру 7, а некоторые — вообще никаких цифр. Сходным образом, на правой картинке люди с нормальным зрением видят число 15, тогда как люди с красно-зеленой слепотой никакого числа не увидят.
Теории цветового зрения. Имеются две основные теории цветового зрения. Первую из них выдвинул Томас Янг в 1807 году. 50 лет спустя его теорию развил Герман фон Гельмгольц.
Согласно трихроматической теории Янга—Гельмгольца (эту теорию называют также трехкомпонентной.— Прим. ред.), хотя человек может различать множество цветов, у него есть только три типа цветовых рецепторов (колбочек). Каждый рецептор чувствителен к широкому диапазону длин волн, но сильнее всего он реагирует на узкий участок диапазона. Как показано на рис. 4.20, рецептор коротких волн наиболее чувствителен к волнам короткой длины (синий цвет), рецептор средних волн — к волнам средней длины (зеленый и желтый цвет), а рецептор длинных волн — к длинным волнам (красный). Совместная работа этих трех рецепторов и определяет ощущение цвета. То есть свет с определенной длиной волны стимулирует эти три типа рецепторов в разной степени, и конкретное соотношение активности в этих рецепторах ведет к ощущению определенного цвета. Следовательно, в дополнение к нашему прежнему разговору о кодировании качества стимула можно сказать, что, согласно трихроматической теории, цветовое качество кодируется паттерном активности трех рецепторов, а не путем использования отдельных рецепторов для каждого цвета.
Рис. 4.20. Трихроматическая теория. Функции ответов рецепторов коротких, средних и длинных волн, согласно трихроматической теории. Эти кривые позволяют определять относительную реакцию каждого рецептора на свет любой длины волны. В показанном здесь примере для определения реакции каждого рецептора на свет с длиной волны 500 нм надо провести линию вверх от отметки «500 нм» и затем пометить, где она пересекает каждую из кривых (по: Wald & Brown, 1965).
Трихроматическая теория объясняет связанные с цветовым зрением факты, которые мы уже упоминали. Во-первых, человек может различать разные длины волн потому, что они воздействуют на три типа рецепторов, вызывая у них неодинаковую реакцию. Во-вторых, закон трех первичных цветов вытекает непосредственно из трихроматической теории. Для любого цвета мы можем
подобрать комбинацию из трех достаточно отстоящих друг от друга длин волн, потому что эти три различные волны активируют три различных типа рецепторов, и именно активность этих рецепторов стоит за восприятием тестового цвета. (Теперь мы понимаем значение числа три.) В-третьих, трихроматическая теория объясняет различные дефекты цветовосприятия отсутствием одного или более из трех типов цветовых рецепторов: у дихроматов с рождения отсутствуют рецепторы одного типа, а у монохроматов — два из трех типов рецепторов. Помимо объяснения этих давно известных фактов, трихроматическая теория позволила биологам открыть эти три типа рецепторов. Теперь мы знаем, что в сетчатке глаза человека действительно сосуществуют колбочки трех типов.
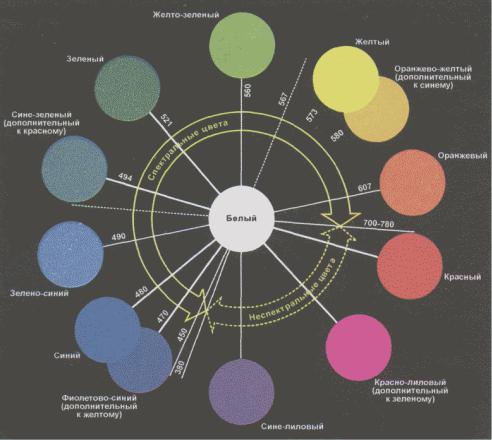
Несмотря на свои успехи, трихроматическая теория не может объяснить некоторые хорошо известные явления цветового восприятия. В 1878 году Эвальд Геринг заметил, что с феноменологической точки зрения все три цвета можно описать как состоящие из одного или двух следующих ощущений: красного, зеленого, желтого и синего. Геринг отметил также, что человек никогда не воспринимает что-либо как красновато-зеленое или желтовато-синее; смесь красного и зеленого скорее будет выглядеть желтой, а смесь желтого и синего — скорее белой. Из этих наблюдений следует, что красный и зеленый образуют оппонентную пару, так же как желтый и синий, и что цвета, входящие в оппонентную пару, не могут восприниматься одновременно. Понятие оппонентных пар получило дальнейшую поддержку из исследований, в которых испытуемый сначала смотрел на цветной свет, а затем — на нейтральную поверхность. При рассматривании нейтральной поверхности испытуемый говорил, что видит на ней цвет, дополнительный первоначальному (рис. 4.21).
Рис. 4.21. Цветовой круг. Простой способ представления цветовых смесей — это цветовой круг. Спектральные цвета (цвета, соответствующие длинам волн в диапазоне чувствительности человека) представлены пятнами, расположенными по периметру круга. Два конца спектра не сходятся; пространству между ними соответствуют неспектральные красные и пурпурные тона, которые можно получить при смешении длинных и коротких волн. Внутри круга находятся цветовые смеси. Цвета, расположенные ближе к центру круга, — менее насыщенные (белее); белый цвет находится в самом центре. Смеси любых двух цветов располагаются на прямой линии, соединяющей два пятна. Если линия проходит через центр круга, то смесь этих цветов, взятых в нужной пропорции, будет выглядеть белой; такие пары цветов называются дополнительными цветами.
Эти феноменологические наблюдения побудили Геринга предложить другую теорию
цветового зрения, названную теорией оппонентных цветов. Геринг полагал, что в зрительной системе имеются два типа цветочувствительных элементов. [В контексте теории Геринга «элемент» следует понимать как устройство, дающее противоположные реакции на цвета оппонентной пары. Согласно Герингу, имеются три таких пары: помимо упомянутых в оригинале двух основных третья пара представляет соотношение «белое—черное». — Прим. ред.] Один тип реагирует на красный или зеленый, другой — на синий или желтый. Каждый элемент противоположно реагирует на свои два оппонентных цвета: у красно-зеленого элемента, например, сила реакции возрастает при предъявлении красного цвета и снижается при предъявлении зеленого. Поскольку элемент не может реагировать сразу в двух направлениях, при предъявлении двух оппонентных цветов одновременно воспринимается белый цвет (рис. 4.21). Теория оппонентных цветов может объяснить наблюдения Геринга, относящиеся к цвету, а также другие факты. Она объясняет, почему мы видим именно те цвета, которые видим. Мы воспринимаем только один тон — красный или зеленый и желтый или синий, — когда баланс смещен только у одного типа оппонентной пары, и воспринимаем комбинации тонов, когда баланс смещен у обоих типов оппонентных пар. Объекты никогда не воспринимаются как красно-зеленые или желто-синие потому, что элемент не может реагировать в двух направлениях сразу. Кроме того, эта теория объясняет, почему испытуемые, которые сначала смотрели на цветной свет, а затем — на нейтральную поверхность, говорят, что видят дополнительные цвета; если, например, испытуемый сначала смотрит на красное, то красная компонента пары утомляется, в результате чего вступает в игру зеленая компонента.
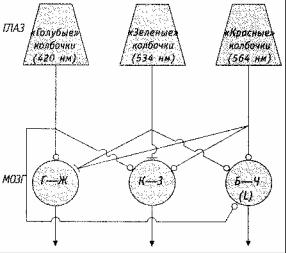
Таким образом, есть две теории цветового зрения — трихроматическая и теория оппонентных цветов, и каждая из них какие-то факты может объяснить, а какие-то нет. Десятилетиями эти две теории считались конкурентными, пока исследователи не предложили компромисс в виде двухстадийной теории, согласно которой три типа рецепторов, предусмотренных в трихроматической теории, поставляют информацию для цвето-оппонентных пар, расположенных на более высоком уровне зрительной системы (Hurvich & Jameson, 1974). Данная точка зрения предполагает, что в зрительной системе должны существовать нейроны, функционирующие как блоки оппонентных цветов и обрабатывающие зрительную информацию после ретины (сетчатки) (которая содержит три рода рецепторов согласно трехкомпонентной теории). И действительно, такие цветооппонентные нейроны были обнаружены в таламусе — одной из промежуточных станций между сетчаткой и зрительной корой (DeValois & Jacobs, 1984). Эти клетки обладают спонтанной активностью, которая повышается при реакции на один диапазон длин волн и снижается при реакции на другой. Так, некоторые клетки, расположенные на более высоком уровне зрительной системы, возбуждаются быстрее, когда сетчатка стимулируется синим светом, чем когда она стимулируется желтым светом; такие клетки составляют биологическую основу сине-желтой оппонентной пары. Суммирующая нейронная проволочная диаграмма, показывающая, как могут быть связаны между собой трихроматическая и оппонентно-процессуальная теории, представлена на рис. 4.22.
Рис. 4.22. Как связаны между собой трихроматическая теория и теория оппонентных процессов. На схеме показано, как три типа рецепторов связаны с продуцированием оппонентнопроцессуальных нейронных реакций на поздних стадиях обработки. Числа в трапециях, изображающих колбочки, указывают длины волн, соответствующих максимальной чувствительности. Линии со стрелками соответствуют связям, повышающим активность; линии с точками соответствуют связям, понижающим активность. Заметьте, что это лишь небольшая
часть всей системы. Существует и другой набор оппонентно-процессуальных элементов, с противоположным характером повышающих и понижающих активность связей.
Это исследование цветового зрения является замечательным примером успешного взаимодействия психологического и биологического подходов к проблеме. В рамках трихроматической теории было выдвинуто предположение, что существуют три типа цветовых рецепторов, и в последующих биологических исследованиях было установлено наличие в сетчатке колбочек трех типов. В теории оппонентных цветов было высказано предположение о существовании в зрительной системе элементов другого рода, и в дальнейшем биологи нашли цветооппонентные клетки в таламусе. Более того, для успешной интеграции этих двух теорий требовалось, чтобы трихроматические клетки поставляли информацию цветооппонентным клеткам, — и это также подтвердилось в последующих биологических исследованиях. Так что во многих случаях проблемная работа на психологическом уровне указывала путь к биологическим открытиям. Неудивительно, что многие ученые приняли анализ цветового зрения в качестве прототипа для анализа работы других сенсорных систем.
Слух
Так же как и зрение, слух является важнейшим средством получения информации об окружении. Для многих из нас это основной канал коммуникации и средство передачи музыки. Как мы увидим, все это возможно благодаря тому, что небольшие изменения звукового давления приводят в колебательное движение мембрану внутреннего уха.
Мы будем рассматривать слух по тому же плану, что и зрение. Сначала мы рассмотрим природу физического стимула, к которому чувствителен слух, потом опишем слуховую систему, уделив особое внимание преобразованиям в рецепторах, и наконец обратимся к кодированию интенсивности и качества звука слуховой системой.
Звуковые волны
Звук возникает вследствие движения или вибрации объекта, — например, когда ветер дует сквозь ветви деревьев. Когда что-либо движется, молекулы находящегося впереди воздуха сжимаются. Эти молекулы толкают другие молекулы и затем возвращаются в исходное положение. Так волна меняющегося давления (звуковая волна) передается по воздуху, хотя отдельные молекулы воздуха далеко не уходят. Эта волна аналогична ряби на поверхности пруда, когда туда бросают камень.
Звуковую волну можно описать графиком давления воздуха в зависимости от времени. График давления в зависимости от времени для одного из видов звука показан на рис. 4.23. На нем представлена синусоидальная волна (названная так потому, что она аналогична синусоидальной функции в математике). Звук, соответствующий синусоидальной волне, называется чистым тоном. Такие звуки важны для анализа слуха, потому что более сложные звуки можно разложить на чистые тона, т. е. на ряд различных синусоидальных волн. Чистые тона определяются двумя параметрами, от которых зависит их ощущение человеком. Один параметр — это частота тона. Частота тона — это количество колебаний за одну секунду (или герц), т. е. частота, с которой молекулы двигаются тудасюда (см. рис. 4.23). Частота — основа воспринимаемой высоты тона, одного из наиболее примечательных качеств звука.
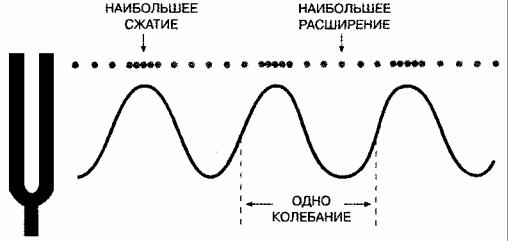
Рис. 4.23. Чистый тон. Вибрирующий камертон создает последовательность волн сжатия и расширения воздуха, подчиняющихся синусоидальному закону. Такой звук называется чистым тоном. Он описывается параметрами частоты и интенсивности. Когда камертон делает 100 колебаний в секунду, он создает звуковую волну со 100 сжатиями в секунду, или с частотой 100 герц. Интенсивность (или амплитуда) чистого тона — это разница в давлении между пиками и впадинами. Форму волны любого звука можно разложить на ряд синусоидальных волн с различной частотой, амплитудой и фазой. Когда эти синусоидальные волны складываются, получается первоначальная форма волны.
Второй параметр чистого тона — его интенсивность (амплитуда), т. е. различие давлений между пиком и впадиной на графике зависимости давления от времени (см. рис. 4.23). Интенсивность
— основа восприятия громкости. Интенсивность звука обычно измеряется в децибелах (дБ); росту интенсивности на 10 децибел соответствует увеличение мощности в 10 раз, росту на 20 децибел — увеличение в 100 раз, 30 децибел — 1000 раз и так далее. Например, тихий шепот в беззвучной обстановке библиотеки имеет интенсивность около 30 децибел, в шумном ресторане уровень звука может равняться 70 децибелам, уровень звука на рок-концерте может достигать 120 децибел, а шум взлетающего самолета может превышать 140 децибел. Постоянное воздействие уровня звука, превышающего 100 децибел, может повлечь за собой необратимую потерю слуха.
И последней характеристикой звука является тембр — наше восприятие сложности звука. Практически ни один из звуков, окружающих нас в повседневной жизни, не является столь простым, как чистые тона, о которых мы говорили выше. (Исключение составляют лишь камертоны и некоторые электронные музыкальные инструменты.) Звуки, издаваемые акустическими инструментами, автомобилями, человеческим голосом, животными и водопадами, характеризуются сложными паттернами звукового давления.
<Рис. Музыкальные инструменты производят сложные паттерны звукового давления. Эти паттерны принято называть тембром звука.>
Слуховая система
Кслуховой системе относятся уши, некоторые участки мозга и проводящие нервные пути. Нас
впервую очередь будут интересовать сами уши; к ним относят не только отростки по обеим сторонам головы, но и весь слуховой орган, большей частью находящийся внутри черепа (рис. 4.24).
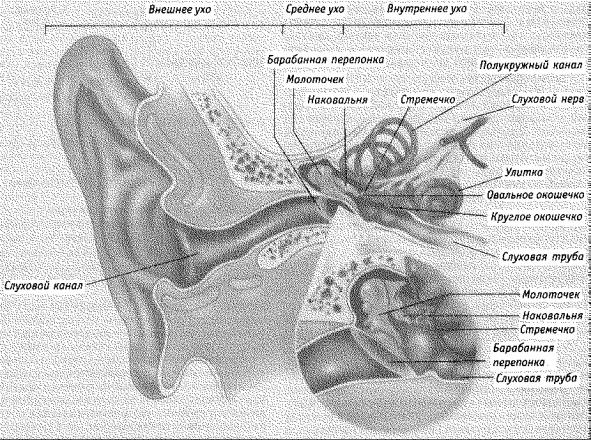
Рис. 4.24. Поперечный разрез уха. На рисунке показано общее строение уха. Внутреннее ухо состоит из улитки, содержащей слуховые рецепторы, и вестибулярного аппарата (полукружные каналы и вестибулярные мешочки), служащего органом для чувства равновесия и движения тела.
Как и глаз, ухо содержит две системы. Одна система усиливает и передает звук к рецепторам, после чего за дело принимается другая система, которая преобразует звук в нервные импульсы. Передающая система включает наружное ухо, состоящее из внешнего уха (pinna — ушная раковина) и слухового канала, а также среднее ухо, состоящее из барабанной перепонки и цепочки из трех костей — молоточка, наковальни и стремечка. Система преобразования расположена в части внутреннего уха, называемой улиткой и содержащей рецепторы звука.
<Рис. Если на рок-концерте сидеть или стоять перед акустическими системами, это может вызвать необратимую потерю слуха.>
Рассмотрим передающую систему подробнее (рис. 4.25). Наружное ухо помогает улавливанию звуков и передает их через слуховой канал к упругой мембране, которая называется барабанной перепонкой. Барабанная перепонка — самая наружная часть внутреннего уха. Ее заставляют вибрировать звуковые волны, приходящие по слуховому каналу. Задача внутреннего уха — передать вибрации барабанной перепонки через заполненную воздухом полость к другой мембране, овальному окошечку, служащему воротами ко внутреннему уху и рецепторам. Эту передачу внутреннее ухо осуществляет посредством механического мостика, построенного из молоточка, наковальни и стремечка. От барабанной перепонки вибрации передаются первой из этих косточек, передающей их второй, которая, в свою очередь, передает их третьей, результатом чего являются вибрации овального окошечка. Это механическое приспособление не только передает звуковую волну, но и значительно усиливает ее.
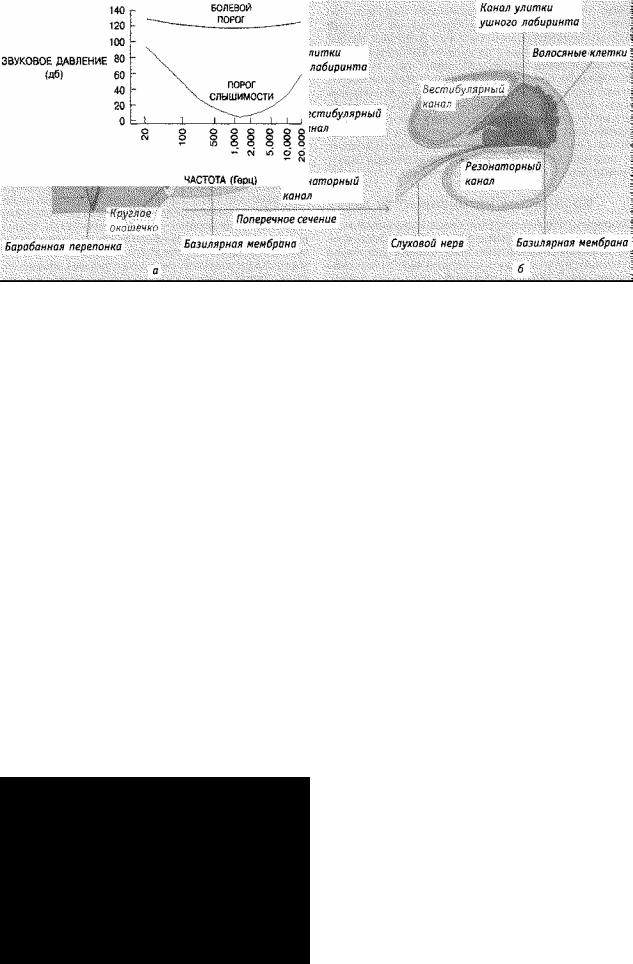
Рис. 4.25. Схематическое строение среднего и внутреннего уха. а) Движения жидкости внутри улитки изгибают базилярную мембрану и стимулируют волосяные клетки, служащие слуховыми рецепторами, б) На поперечном сечении улитки показана базилярная мембрана и волосяные клетки-рецепторы.
Теперь рассмотрим систему преобразования. Улитка — это спиралевидная трубка из костного вещества. Мембраны разделяют ее на секции, заполненные жидкостью; одна из мембран — базилярная, к ней прикреплены слуховые рецепторы (см. рис, 4.25). Эти рецепторы называются волосяными клетками, потому что по строению они похожи на волоски, проникающие в жидкость. Давление на овальном окошечке (соединяющем среднее и внутреннее ухо) создает изменения давления жидкости в улитке, что, в свою очередь, заставляет базилярную мембрану вибрировать, приводя к изгибанию волосяных клеток и появлению электрического импульса. Таков сложный процесс преобразования звуковой волны в электрический импульс. Нейроны, синаптически соединенные с нервными клетками, имеют длинные аксоны, которые образуют часть слухового нерва. Большинство слуховых нейронов соединены с отдельными нервными клетками. В слуховом нерве около 31 000 слуховых нейронов, что гораздо меньше одного миллиона нейронов, составляющих зрительный нерв (Yost & Nielson, 1985). От каждого уха слуховые пути идут к обеим сторонам мозга и заканчиваются на синапсах различных ядер, прежде чем достигают слуховой коры.
Восприятие интенсивности звука
Вспомним, что наше зрение более чувствительно к одним длинам волн, чем к другим. В слуховом восприятии есть аналогичное явление. Человек более чувствителен к звукам в середине частотного диапазона, чем к звукам с частотой ближе к его краям. Это показано на рис. 4.26, где приведена зависимость абсолютного порога интенсивности звука от частоты. У многих людей слух в той или иной степени ослаблен, и порог у них выше того, что показан на рис. 4.26.
Рис. 4.26. Абсолютный порог для слуха. Нижняя кривая показывает абсолютную пороговую интенсивность для различных частот. Наибольшая чувствительность наблюдается в окрестностях частоты 1000 герц. Верхняя кривая показывает болевой порог (данные аппроксимированы по различным источникам).
Есть два основных варианта недостаточности слуха. При одном из них пороги повышаются примерно в равной степени для всех частот в результате плохой проводимости среднего уха (потеря проводимости). В другом случае потери слуха порог повышается в неравной степени, причем более всего он повышается на высоких частотах. Такая ситуация обычно является следствием повреждения внутреннего уха и часто связана с частичным разрушением волосковых клеток (потеря нервной чувствительности). Волосковые клетки после разрушения не восстанавливаются. Потеря нервной чувствительности возникает у многих пожилых людей. Вот почему им часто трудно расслышать высокие звуки. Однако потеря нервной чувствительности не происходит исключительно у пожилых. Она возникает и у молодых, если на них воздействует чрезмерно громкий звук. Необратимой потерей слуха обычно страдают рок-музыканты, работники взлетно-посадочных полос в аэропортах и работающие с отбойным молотком. Например, у Пита Таунзенда, известного гитариста из группы «The Who», возникло серьезное ослабление слуха из-за того, что на него постоянно воздействовала громкая рок-музыка; с тех пор он предупреждал многих молодых людей об этой опасности.
Естественно предположить, что воспринимаемая интенсивность звука одинакова для обоих ушей, но на самом деле здесь есть тонкие различия. Если звук приходит справа, то для правого уха его слышимость будет больше, чем для левого; это происходит потому, что голова образует «звуковую тень», которая снижает интенсивность звука, доходящего до дальнего уха. Но это вовсе не ограничение слуховых возможностей, поскольку человек использует величину междуушного расхождения в интенсивности для локализации направления звука (это как если бы мы рассуждали, что «если интенсивность звука в моем правом ухе больше, чем в левом, должно быть, звук пришел справа»). Аналогично, звук, приходящий с правой стороны, поступает в правое ухо на долю секунды раньше, чем в левое (и наоборот, если звук пришел слева). Человек также использует это междуушное расхождение во времени, чтобы локализовать звук («если звук сначала пришел в мое правое ухо, значит, он пришел справа»).
Восприятие высоты звука
Высота и частота. Когда мы слышим чистый тон, то воспринимаем не только его громкость, но и высоту. Подобно тому как цвет — главное качество света, так и высота — главное качество звука, ранжированного по шкале от низкого до высокого. И подобно тому как цвет определяется частотой света, высота определяется частотой звука. При возрастании частоты высота увеличивается. Как и длину световой волны, частоту звука человек различает очень хорошо. Молодой взрослый может слышать частоты в диапазоне от 20 до 20 000 герц (колебаний в секунду), причем ЕЗР составляет менее 1 герца при частоте 100 герц и возрастает до 100 герц при 10 килогерцах.
Однако в слуховом восприятии нет ничего похожего на смешение цветов. Когда две и более частот звучат одновременно, можно слышать высоту каждой частоты при условии, что они достаточно различаются. Если частоты различаются несильно, ощущение будет более сложным, но все равно звук не будет похож на один чистый тон. При изучении цветового восприятия обнаружение того факта, что смешение трех цветных источников света дает ощущение одного цвета, привело к идее о трех типах рецепторов. Отсутствие аналогичного явления в слуховом восприятии позволяет предположить, что если есть рецепторы, настроенные на различные частоты, то типов таких рецепторов должно быть множество.
Теории восприятия высоты звука. Как и в случае цветового зрения, для объяснения того, как частота кодируется ухом в высоту звука, были предложены две теории.
Первая теория была создана британским физиком Резерфордом в 1886 году. Он предположил, что: а) звуковая волна заставляет вибрировать всю базилярную мембрану и частота вибраций соответствует частоте звука; б) частота вибраций мембраны задает частоту нервных импульсов, передаваемых по слуховому нерву. Так, тон частотой 1000 герц заставляет базилярную мембрану вибрировать 1000 раз в секунду, в результате чего волокна слухового нерва разряжаются с частотой 1000 импульсов в секунду, а мозг интерпретирует это как определенную высоту. Поскольку в этой теории предполагается, что высота зависит от изменений звука во времени, ее назвали временной теорией (ее называют также частотной теорией).
Гипотеза Резерфорда вскоре встретилась с серьезными проблемами. Было доказано, что нервные волокна могут передавать не более 1000 импульсов в секунду, и тогда неясно, как человек воспринимает высоту тона с частотой более 1000 герц. Вивер (Weaver, 1949) предложил способ
спасения временной теории. Он предположил, что частоты выше 1000 герц кодируются различными группами нервных волокон, каждая из которых активируется в несколько разном темпе. Если, например, одна группа нейронов выдает 1000 импульсов в секунду, а затем 1 миллисекунду спустя другая группа нейронов начинает выдавать 1000 импульсов в секунду, то комбинация импульсов этих двух групп даст 2000 импульсов в секунду. Эту версию временной теории подкрепило открытие, что паттерн нервных импульсов в слуховом нерве повторяет форму волны стимульного тона, несмотря на то, что отдельные клетки реагируют не на каждое колебание (Rose et al., 1967).
Однако способность нервных волокон отслеживать форму волны обрывается примерно на частоте 4000 герц; тем не менее мы можем слышать высоту звука, содержащего гораздо более высокие частоты. Отсюда следует, что должно существовать другое средство кодирования высотного качества звука, по крайней мере на высоких частотах.
Другая теория восприятия высоты звука относится к 1683 году, когда французский анатом Жозеф Гишар Дювернье предположил, что частота кодируется высотой звука механически, путем резонанса (Green & Wier, 1984). Чтобы разобраться в этом предположении, полезно сначала рассмотреть пример резонанса. Когда ударяют по камертону, который находится рядом с пианино, струна пианино, настроенная на частоту камертона, начинает колебаться. Если мы говорим, что ухо работает по тому же принципу, это значит, что в нем есть некая структура, сходная по конструкции со струнным инструментом, причем различные ее части настроены на различные частоты, так что когда на ухо предъявляется некоторая частота, соответствующая часть этой структуры начинает колебаться. Эта идея была в общем правильной: такой структурой оказалась базилярная мембрана.
В XIX веке Герман фон Гельмгольц, исходя из гипотезы резонанса, предложил для объяснения восприятия высоты теорию локальности. Согласно этой теории, каждый конкретный участок базилярной мембраны, когда он начинает реагировать, создает ощущение определенной высоты тона. Предполагаемое множество участков на мембране согласуется с фактом существования множества рецепторов высоты. Заметьте, что теория локальности не означает, что мы слышим звук базилярной мембраной; просто от того, какие участки мембраны вибрируют, в наибольшей степени зависит, какую высоту мы услышим. Это пример органа чувства, в котором кодирование качества осуществляется путем «включения» тех или иных нервных волокон.
Как именно колеблется базилярная мембрана, не было известно до 1940 года, когда Георг фон Бекеши измерил ее движения при помощи маленьких отверстий, просверленных в улитках морских свинок и человеческих трупов. Учитывая результаты Бекеши, потребовалось модифицировать теорию локальности; базилярная мембрана вела себя не как пианино с раздельными струнами, а как простыня, которую встряхнули за один конец. В частности, Бекеши показал, что при большинстве частот вся базилярная мембрана приходит в движение, но место наиболее интенсивного движения зависит от конкретной частоты звучания. Высокие частоты вызывают вибрацию в ближнем конце базилярной мембраны; по мере повышения частоты паттерн вибрации сдвигается к овальному окошечку (Bekesy, 1960). За это и другие исследования слуха Бекеши получил в 1961 году Нобелевскую премию.
Как и временные теории, теория локальности объясняет многие, но не все явления восприятия высоты звука. Основные затруднения у теории локальности связаны с тонами низких частот. При частотах ниже 50 герц все части базилярной мембраны вибрируют примерно одинаково. Это значит, что все рецепторы активируются в равной степени, из чего следует, что у нас нет способа различения частот ниже 50 герц. На самом же деле мы можем различать частоту всего в 20 герц.
Таким образом, теории локальности затрудняются объяснить восприятие низкочастотных звуков, а временные теории — восприятие высоких частот. Все это навело на мысль, что восприятие высоты звука определяется как временными паттернами, так и паттернами локализации, причем временная теория объясняет восприятие низких частот, а теория локальности — восприятие высоких частот. Ясно, однако, что там, где один механизм отступает, начинает преобладать другой. На самом деле не исключено, что частоты от 1000 до 5000 герц обслуживаются обоими механизмами (Coren, Ward & Enns, 1999).
Поскольку наши уши и глаза играют столь важную роль в нашей повседневной жизни, были предприняты значительные усилия, направленные на то, чтобы заменить их на искусственные у индивидуумов, страдающих неизлечимыми дефектами этих органов. Некоторые из этих усилий описаны в рубрике «На переднем крае психологических исследований».
Другие ощущения
По сравнению со зрением и слухом, другим ощущениям недостает тех богатых функциональных возможностей, из-за которых зрение и слух называют «высшими чувствами». И все же эти другие чувства жизненно важны. Например, ощущение запаха (обоняние) является одним из наиболее примитивных и наиболее важных из этих ощущений. Возможно, это связано с тем, что запах проникает в мозг по более прямому маршруту, чем любые другие ощущения. Рецепторы, расположенные в носовой полости, связаны с мозгом без посредства синапсов. Более того, в отличие от зрительных и слуховых рецепторов, обонятельные рецепторы испытывают непосредственное воздействие окружающей среды — они находятся прямо в носовой полости и не имеют перед собой защитной оболочки. (Тогда как зрительные рецепторы расположены позади роговой оболочки, а слуховые защищены наружным и средним ухом.) Поскольку запах с очевидностью является важной сенсорной модальностью, мы начнем наше обсуждение других ощущений с ощущения запаха, называемого также обонянием.
Обоняние
Чувство запаха, или обоняние, помогает нашему выживанию: оно необходимо для обнаружения испорченной пищи или незакрытого газа, а потеря обоняния может привести к притуплению аппетита. И все же для многих других биологических видов обоняние еще важнее. Поэтому неудивительно, что у них обонянию отведена большая часть коры, чем у нас. У рыб обонятельная кора почти целиком охватывает полушария мозга, у собак — примерно одну треть, у человека — всего одну двадцатую часть. В этом отражены межвидовые различия в обонятельной чувствительности. Пользуясь преимуществом превосходной обонятельной способности собак, Почтовая служба Соединенных Штатов и Таможенное бюро готовят их для проверки невскрытых упаковок на героин. А специально натренированные полицейские собаки могут разнюхать спрятанную взрывчатку.
Поскольку обоняние у других видов развито столь хорошо, они часто используют его как ведущее средство коммуникации. Насекомые и некоторые высшие животные выделяют химические вещества, известные как феромоны и распространяющиеся по воздуху, так чтобы их могли унюхать другие представители этого же вида. Например, самка мотылька может выделять настолько сильный феромон, что самцов влечет к ней с расстояния в несколько миль. Установлено, что самец мотылька реагирует именно на феромон, а не на вид самки; его будет влечь к самке, находящейся в контейнере из проволочной сетки, несмотря на то, что ее вид недоступен, но не к самке в стеклянном контейнере, где ее хорошо видно, но путь для запаха блокирован.
Насекомые пользуются запахом, чтобы сообщать не только о «любви», но и о смерти. Когда муравей умирает, химические вещества, образующиеся при разложении его тела, стимулируют других муравьев отнести его тело на мусорную кучу снаружи гнезда. Если живого муравья пропитать этим феромоном разложения, другие муравьи тут же относят его на мусорную кучу. Когда он возвращается в гнездо, его уносят опять. Эти попытки преждевременных похорон продолжаются, пока «запах смерти» не выдохнется (Wilson, 1963).
Остались ли у нас, людей, пережитки этой примитивной системы общения? Эксперименты показывают, что как минимум мы можем отличать по запаху себя от других и мужчин от женщин. В одном из исследований испытуемые носили майку в течение 24 часов, не принимая душ и не пользуясь дезодорантом. Затем они сдавали майки экспериментатору. Каждому испытуемому экспериментатор предъявлял для обнюхивания три майки: собственную майку испытуемого, одну мужскую и одну женскую.
Основываясь только на запахе, большинство испытуемых обычно могли отличить свою собственную майку, а также определить, какую из двух остальных носил мужчина, а какую — женщина (Russel, 1976; Schleidt, Hold & Attili, 1981). Другие исследования показывают, что по запаху мы можем определять и более тонкие вещи. Между женщинами, которые живут или работают вместе, видимо, происходит обмен информацией посредством запаха относительно их менструального цикла, так что со временем их менструальные циклы синхронизируются и начинаются в одно время (Russel, Switz & Thompson, 1980; McClintock, 1971).
Система обоняния. Стимулом для запаха являются испускаемые веществом летучие молекулы. Молекулы выходят из вещества, проносятся по воздуху и входят в носовой проход (рис.