

Сальтаторное проведение потенциалов действия
Как указывалось выше, если какой-либо участок волокна в силу каких-либо причин не формирует собственный ПД, то этот «блокированный» участок может быть преодолён электротонически (вспомните понятие «перескок»). Этот тип проведения можно было бы назвать «сальтаторным» (от лат. salto — прыжок). Однако чаще этот термин относят к проведению ПД по так называемым миелинизированным нервным волокнам.
Впервые прямые электрофизиологические исследования проведения возбуждения в миелинизированнных нервных волокнах лягушки провели Като (1924) и Тасаки (1953). Однако предположение о скачкообразном распространении возбуждения в нервных волокнах впервые было высказано Б.Ф.Вериго (1899).
Миелинизированные нервные волокна характерны для «скоростных линий» нервной связи у позвоночных животных. Миелиновые муфты волокон, каждая из которых формируется одной шванновской клеткой, являются хорошими электроизоляторами: у миелина R = 160 кОмсм2 , С = 5 нФ/см.
Поэтому в миелинизированном волокне проводят электрические токи и генерируют в ответ на них ПД, по существу, только узкие (1 мкм) оголенные участки, расположенные между муфтами, — перехваты Ранвье.
Итак распространение ПД осуществляется скачкообразно (сальтаторно) от перехвата к перехвату (рис. 0810000500).
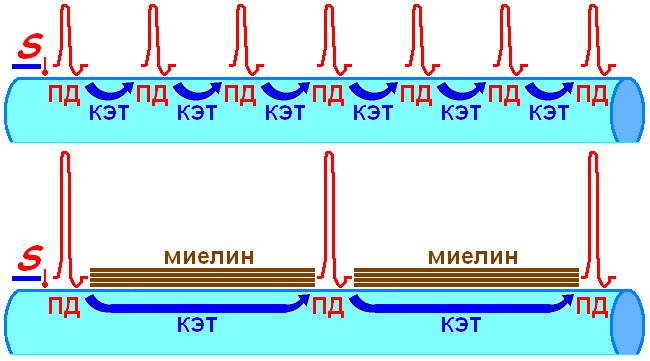
Рис. 0810000500. Проведение возбуждения в немиелизированных (вверху) и миелизированных (внизу) нервных волокнах.
|
|
Миелиновые сегменты гораздо протяженнее перехватов (1 2 мм против 1 мкм). Длина миелиновых сегментов пропорциональна диаметру волокна: миелиновые нервные волокна диаметром 10 – 20 мкм имеют сегменты длиной 1 – 2 мм, а тонкие волокна диаметром 1 – 2 мкм — 0,2 мм.
Однозначно, сальтаторное проведение повышает скорость передачи ПД, за счёт включения быстрой КЭТ-составляющей (табл. 210030704).
Таблица 210030704
Протяжённость возбуждённого участка у некоторых возбудимых структур
Возбудимая структура |
Длительность ПД, мс |
Скорость распространения ПД, мм/мс |
Протяженность возбуждённого участка, мм |
Гигантский аксон кальмара |
1 |
25 |
25 |
Тонкие немиелинизированные волокна позвоночных |
1 |
1 |
1 |
Мышечные волокна позвоночных |
2 |
3 |
6 |
Миелинизированные волокна млекопитающих (d = 20 мкм) |
0,4 |
120 |
48 |
![]()
Принято считать, что сальтаторная форма функционирования проводника экономична в смысле расхода ионов, нагрузки на ионный насос и экономии энергии. Однако это вызывает сомнение. За увеличение скорости проведения необходимо платить! Ведь «бег» требует больше энергии, чем «ходьба»?!
Так плотность натриевых каналов в перехватах Ранвье очень велика — 10 000 мкм‑2, что в 200 раз превышает плотность их в мембране гигантского аксона кальмара.
В перехватах Ранвье происходят весьма большие траты энергии вследствие большой плотности здесь натрий-калиевых каналов.
Миелинизация нервных волокон у позвоночных – важное направление в эволюции нервных элементов