Соматичний гіпермутагенез


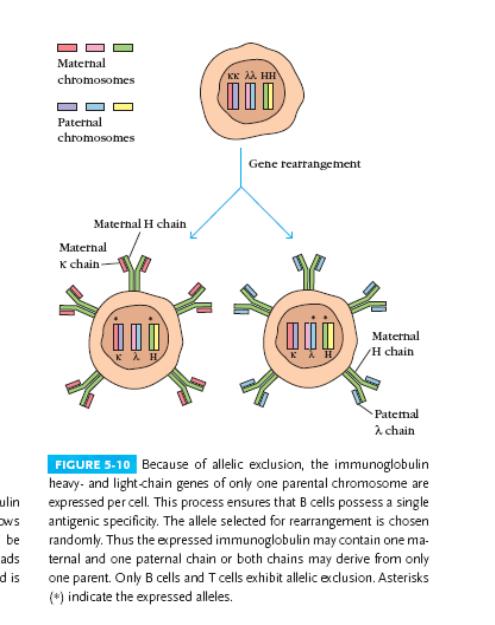
Алельне виключення (рис. Далі, звернути увагу на зірочки).
Експериментально було показано, що кожна В клітина продукує антитіла тільки однієї специфічності. Але ж у кожного гетерозиготного організма є два набори генів, із материнського і батьківського геномів. Гени імуноглобулінів підлягають алельному виключенню, механізм якого до кінця не з'ясований. Вважають, що функціональна перебудова одного алеля є негативним сигналом для другого.
Попередник В клітини має два набори кластерів Н-, к- і λ-генів. Спочатку починає рекомбінувати один алель Н-гену, батьківський або материнський (відбір є цілком випадковим). Після декількох циклів проліферації активується один із алелей L-гена (теж випадково). DJ рекомбінація відбувається в обох хромосомах (материнській і батьківській) на рівні ранніх про-В клітин , а VDJ - тільки в одній (пізні про -В-клітини). Якщо перебудова пройшла успішно, продукція повноцінного μ- ланцюга припиняє реорганізації в Н-генах і сигналізує про початок перебудов в L-генах. Спочатку рекомбінують VJ-гени к-ланцюга; якщо такий варіант невдалий, випробуються інші варіанти VJ κ; якщо знов невдало - рекомбінують гени λ ланцюга. Перша вдала комбінація дає функціональний ІgМ і припиняє подальшу реорганізацію, а саме виключаються гени RAG. Наявність двох пулів генів L-ланцюга, к і λ, підвищує надійність утворення повноцінної молекули імуноглобуліну.
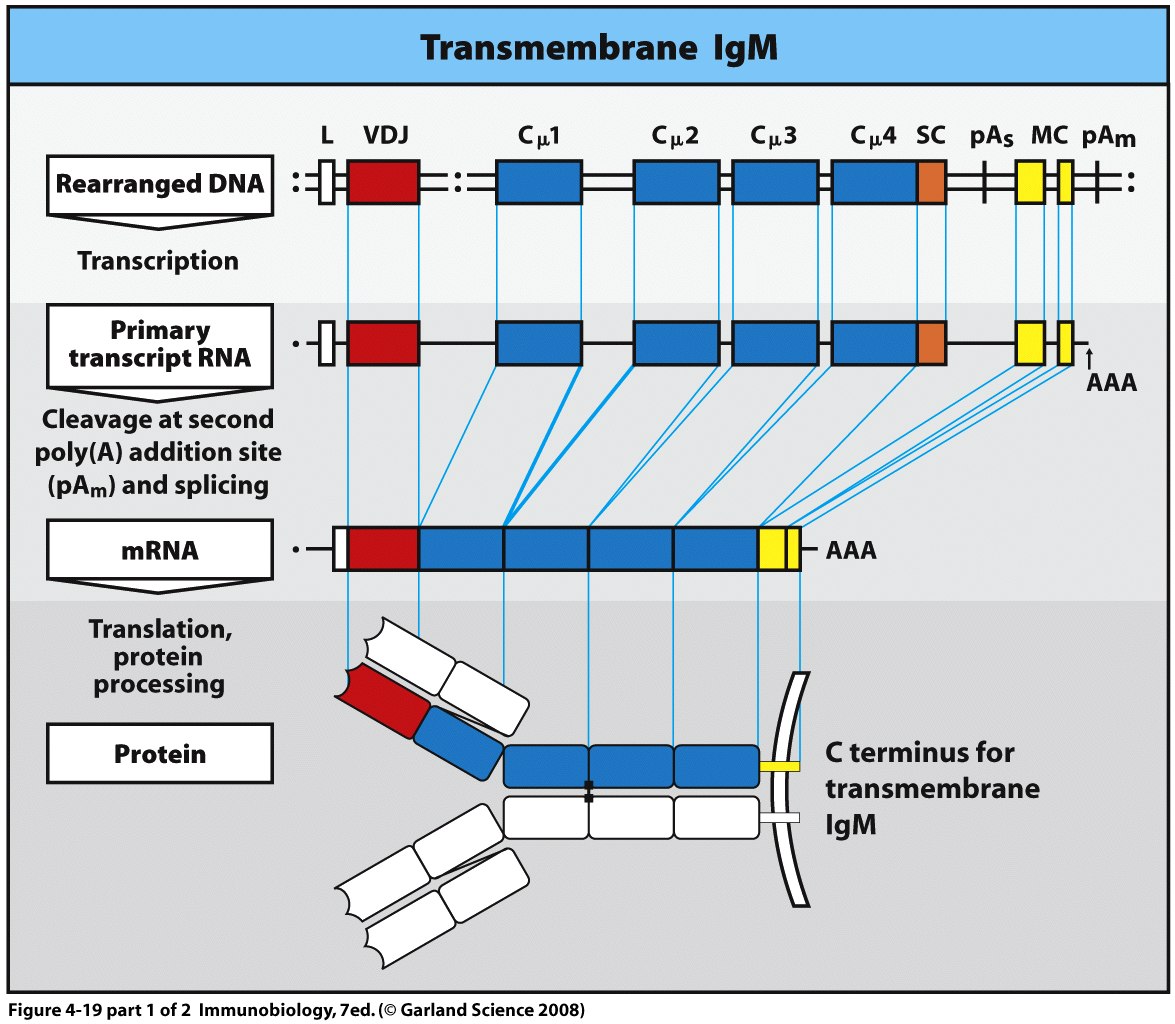
Переключення з мембранної на секретовану форму ІgМ.
Імуноглобуліни можуть існувати як в мембранній (рецептори В лімфоцитів), так і в розчинній формі (антитіла). Вони відрізняються природою С-кінцевого пептиду: у мембраній формі він гідрофобний, і є частиною (трансмембранною) рецептору, а у секретованої - гідрофільний. Для ІgМ показано наявність гідрофобних залишків між 568 і 594 а.з.

Розглянемо механізм переключення на прикладі ІgМ. При цьому у процес є залученими як механізми альтернативного сплайсингу, так і поліаденілування мРНК.
І мембранний, і секретований ІgМ кодуються одними генами. В Сμ гені є фрагмент ДНК, що кодує гідрофільний пептид, і фрагмент, що кодує гідрофобний пептид, їх транскрипція зупиняється, відповідно, двома стоп-кодонами. Спочатку, коли клітина ще не секретує iмуноглобуліни, працює стоп-кодон 2, при цьому відбувається навішування полі –А хвоста в мРНК після цього кодону (принциповий момент), а внутрішній полі-А сигнал ігнорується). Утворюється транскрипт, що несе інформацію як про гідрофобний, так і про гідрофільний пептиди. На наступному етапі фрагмент, що відповідає за гідрофільний пептид, видаляється шляхом сплайсингу мРНК. Таким чином синтезуються ІgМ, зв’язані з плазматичною мембраною, оскільки ці мРНК на 3'-кінці містять екзон, який кодує ділянку поліпептидного ланцюга , що складається з гідрофобних амінокислот (лише 595-597- гідрофільні а.з. в цитоплазматичній частині СООН –кінця ІgМ).
На наступному етапі, коли клітина отримує відповідні сигнали, то в результаті альтернативного сплайсингу утворюється мРНК, в якій залишається інтрон, що містить стоп-кодон 1 (внутрішній полі-А сигнал спрацьовує) . Відбувається більш раннє поліаденілювання після стоп-кодону 1 і делетується та частина , яка кодувала гідрофобну ділянку поліпептидного ланцюга, і транскрибується короткий ген .Таким чином починають секретуватись розчинні форми ІgМ .