

Гени легких ланцюгів Події , що відбуваються при синтезі легких ланцюгів.
В процесі дозрівання в результаті рекомбінації ДНК відбувається зближення одного із V та J сегментів (V + J) – 1 рекомбінаційна подія.
При цьому всі гени, що знаходяться між ними, видаляються із хромосомної ДНК. Після цього з сайту ініціації транскрипції вказаного V –гену починається транскрипція з лідерної ділянки, яка закінчується на термінаторі транскрипції гена С. Утворена пре-мРНК підлягає сплайсингу, при цьому видаляються інтрони та додаткові J-сегменти, проходить з′єднання екзонів варіабельної і константної області та лідерної послідовності, після кепування та поліаденілювання така мРНК транслюється у легкий ланцюг каппа типу або за такими ж правилами – лямбда типу, не дивлячись на те, що в останньому J і С- сегменти є зчепленими.
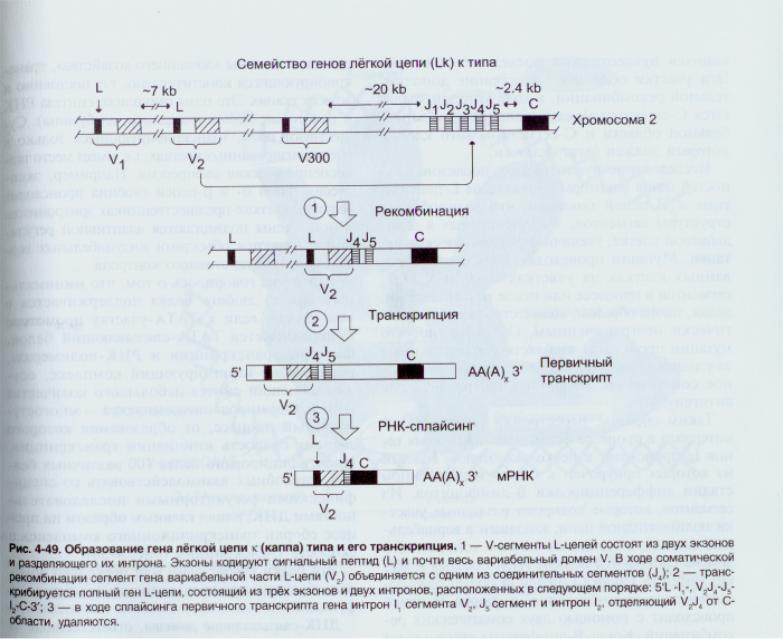
Для генів легких ланцюгів: V-сегмент кодує 1-95 ам.зал., J сегмент-96-108, далі С-сегмент. (Всього-220-230 а.з.).
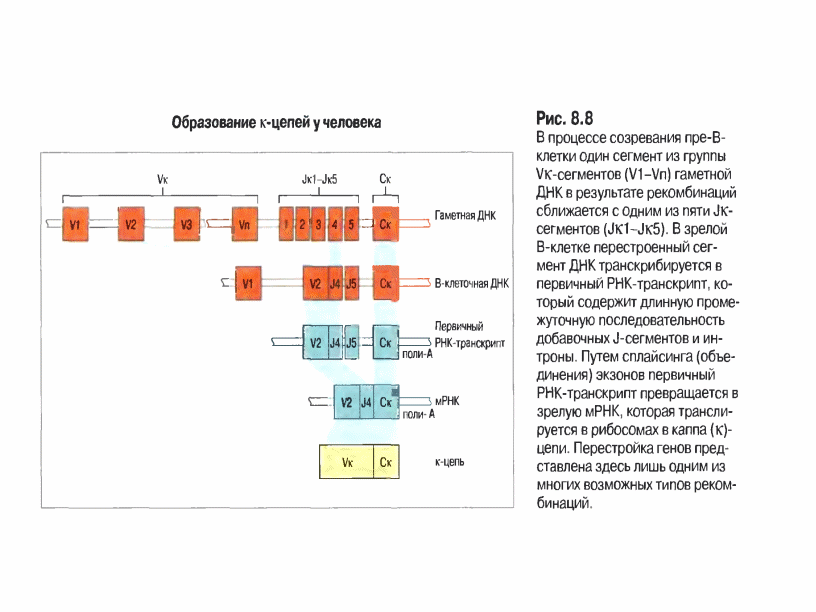

Але до складу генів V- доменів, як і константих, входять як екзони, так і інтрони, які делетуються під час сплайсингу первинного транскрипту (рис).
Гени важких ланцюгів
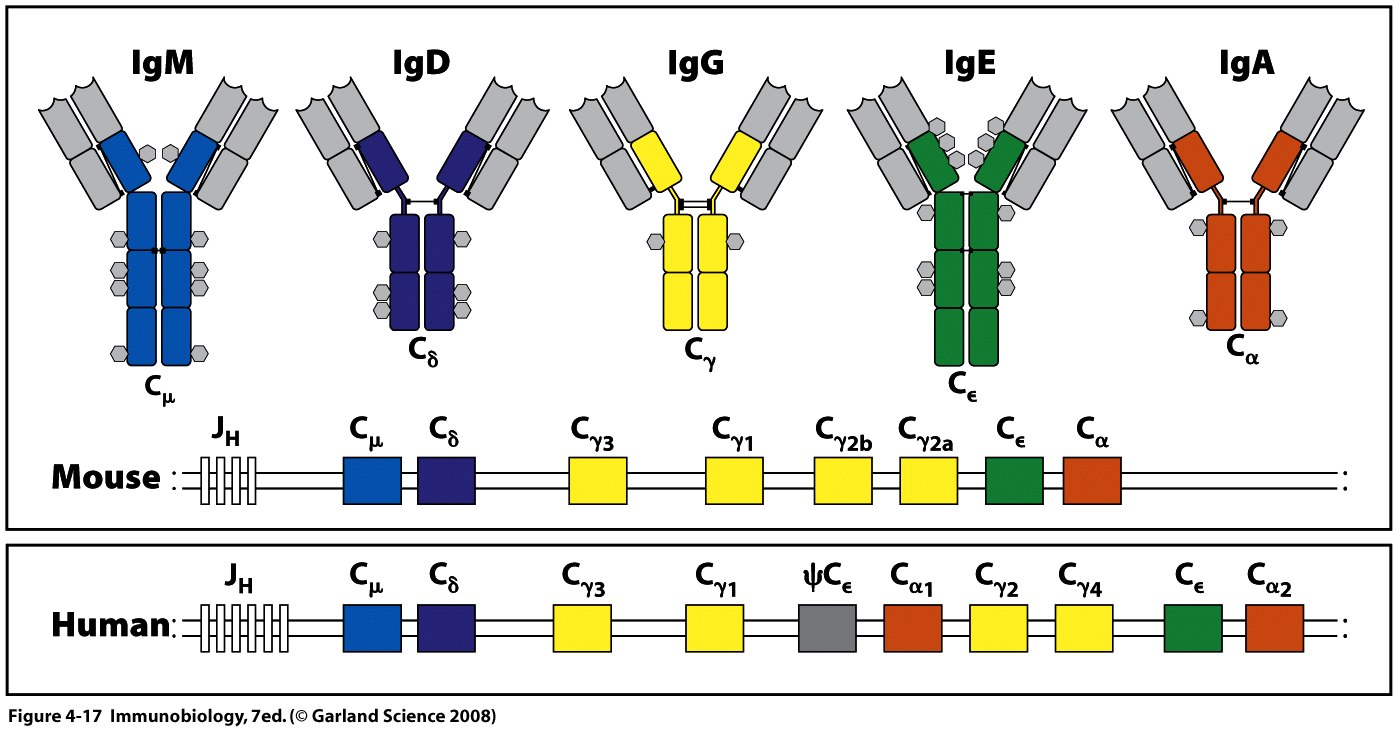
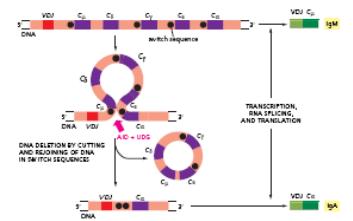
При формуванні функційного гену важкого ланцюга відбувається ряд перебудов на рівні ДНК, а саме 2 рекомбінаційні події (V +D J ).Спочатку відбувається об'єднання вибраних D J сегментів. При цьому всі сегменти ДНК, що є між ними, делетуються. На другому етапі- об'єднання перебудованих D J з V сегментами з наступним видаленням всіх проміжних сегментів ДНК. Таким чином шляхом рекомбінації V ,D ,J сегментів утворюється перебудований V ген . Нижче J генів розташовані всі гени (відстань між V і С – 8,5 тис. п.н.) константних областей, причому у миші по одному гену для кожного класу Ig, а також для кожного підкласу IgG. Гени константної області , що кодують важкі ланцюги у мишей, показані на рис. Після цього починається транскрипція, яка закінчується на термінаторі транскрипції, що розташований після кластера генів Сμ і С δ. Перед кожним СН екзоном , крім С δ, містяться ділянки переключення- свіч- сайти (SWICH SITE=S), які побудовані з нуклеотидних послідовностей, що повторюються і є копіями послідовності swich site перед екзоном Сμ .Саме swich site дозволяють певному СН гену рекомбінувати з перебудованим геном варіабельних доменів та полегшують протікання або альтернативного сплайсингу у випадку IgM –IgD, або додаткової рекомбінації , в ході якої видаляються С-сегменти між повним геном варіабельної області, та С-сегментом того класу, який має бути включеним. Утворена пре-мРНК підлягає сплайсингу, при цьому видаляються інтрони та додаткові J і D сегменти, проходить з′єднання екзонів варіабельної і константної області та лідерної послідовності ,після кепування та поліаденілювання така мРНК транслюється. Для важких ланцюгів- V –кодує 1-99 ам-ти, D -100-107 ам-ти, J- 108-123 ам-ти. Всього від 440-450 ам-т ( крім IgM і IgЕ ).


Ізотипи імуноглобулінінів кодуються кластерами генів константних доменів важких ланцюгів
Механізми диверсифікації (урізноманітнення)=(diversification) імуноглобулінів.
Комбінаторна диверсифікація (див. табл.)
1) L -ланцюги :
λ -ланцюг - VJ: 30Vх 4J λ = 120;
κ –ланцюг- VJ: 40V х 5Jк =200;
120+ 200= 320 варіантів L-ланцюга
2) Н ланцюги:
DJ: 27D х 6J = 162;
V(DJ): 51 х 162 = 8262;
D ген має три вірогідні рамки зчитування: 8262 х 3 = 24786 варіантів
Н-ланцюга.
3) Комбінація Н х L: 320 х 24786 = 7,9 х 106
3. Мутації у сайтах рекомбінації, генна конверсія, випадковість сайтів розщеплення за участі ендонуклеази Artemis, додавання Р і N-нуклеотидів - додають різноманітності >108.
Таким чином, за рахунок комбінацій обмеженої кількості V, D, J генів утворюється дуже велика кількість потенційних варіантів антитіл (і відповідно, рецепторів В лімфоцитів).
Згадані механізми утворюють доімунний репертуар. Після зустрічі з антигеном зрілі В клітини, що знаходяться у вторинних лімфоідних органах, підлягають соматичним мутаціям. В селезінці і лімфовузлах утворюються спеціальні морфологічні структури, так звані зародкові центри, де в генах імуноглобулінів включається механізм гіпермутагенезу і вже перебудовані V-гени починають швидко змінюватись (в основному завдяки вставкам і замінам). Швидкість накопичення мутацій сягає 10-3 на пару нуклеотидів на генерацію клітин. Це означає, що у кожному поколінні утворюється одна мутація на 1000 пар нуклеотидів - у 106 разів частіше, ніж відбуваються мутації в інших генах. В результаті соматичних мутацій утворюється багато варіантів антитіл, які не дуже відрізняються одне від одного. Особливо це зв’язано з районами CDR- різноманіття CDR 2 закодоване в геномі, CDR 1 утворюються за рахунок соматичних мутацій, а CDR 3- рекомбінації.
Антиген, що присутній при цьому, служить фактором відбору і сприяє проліферації (розмноженню) тих клітин, рецептори яких якнайбільше до нього підходять. Відбувається тонка підгонка репертуару В клітин, які частково перетворюються на клітини пам'яті. При повторному введенні антигену, у вторинній відповіді, саме ці В клітини активуються, тому антитіла вторинної відповіді є більш специфічними, ніж первинної. Відбувається так зване созрівання імунної відповіді, підвищення її афінності, спорідненості до антигену. Явище гіпермутагенезу описане тільки для генів імуноглобулінів.
Сьогодні вважається актуальним залучення ферменту активаційно-індукованої дезамінази (АІД) до процесу соматичного гіпермутагенезу. Цей фермент експресується тільки в активованих В клітинах, дезамінує цитозин до урацилу в локусах генів, що кодують варіабельні області антитіл. Накопичуються некласичні У-Г пари (замість Ц-Г), що призводить до репараційних процесів, пов’язаних з виникненням різних типів мутацій.