

Дозрівання в-лімфоцитів

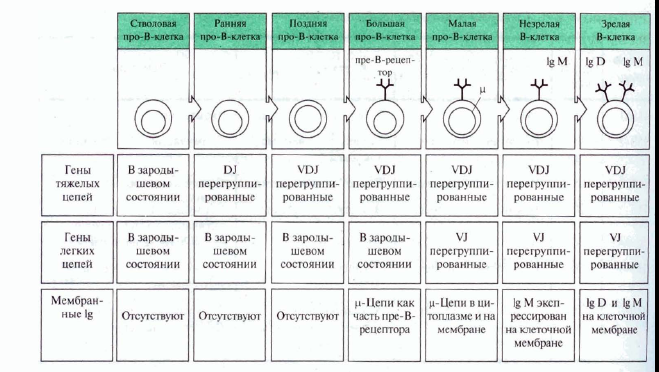
0-лімфоїдна стовбурова кровотворна клітина.
А-ранні про-В-клітини-не мають рецепт. В клітин, початок реаранжування DJ-сегментів в локусі важкого (Н) ланцюга імуноглобулінів, може відбуватись в обох гомологічних хромосомах.
В- пізні про-В-клітини- V- DJ реаранжування генів- підлягає алельному виключенню- тобто проходить лише в одній хромосомі. Отже на цій стадії перебудовуються гени і екс пресується важкий ланцюг в комплексі з сурогатним легким, що є сигналом для позитивної селекції цих клітин.
С- великі пре-В-клітини- експресують пре-В-клітинний рецептор, мають форму бластів, утворюють клони : 6 поділів, не працюють рекомбінази.
D- малі пре-В-клітини, перебудова генів легкого κ–ланцюга, який експресується на мембрані разом з перебудованим μ- важким ланцюгом. Непродуктивність одного гену-включається інший, ні- перебудова генів в λ-локусі.
E- незрілі В-клітини, мають сформовані Ig M-рецептори, але підлягають негативному відбору. На цій стадії В клітини виходять з кісткового мозку до селезінки і вступають на стадію дозрівання.
F- зрілі наївні В-клітини, які рециркулюють (до зустрічі з антигеном).
Таким чином, перебудова генів та синтез білкового продукту пов’язана з:
А) На стадії про-В-клітини з перебудовою важкого ланцюга Іg.
Б) На стадії пре-В-клітин- перебудова легкого ланцюга Іg.
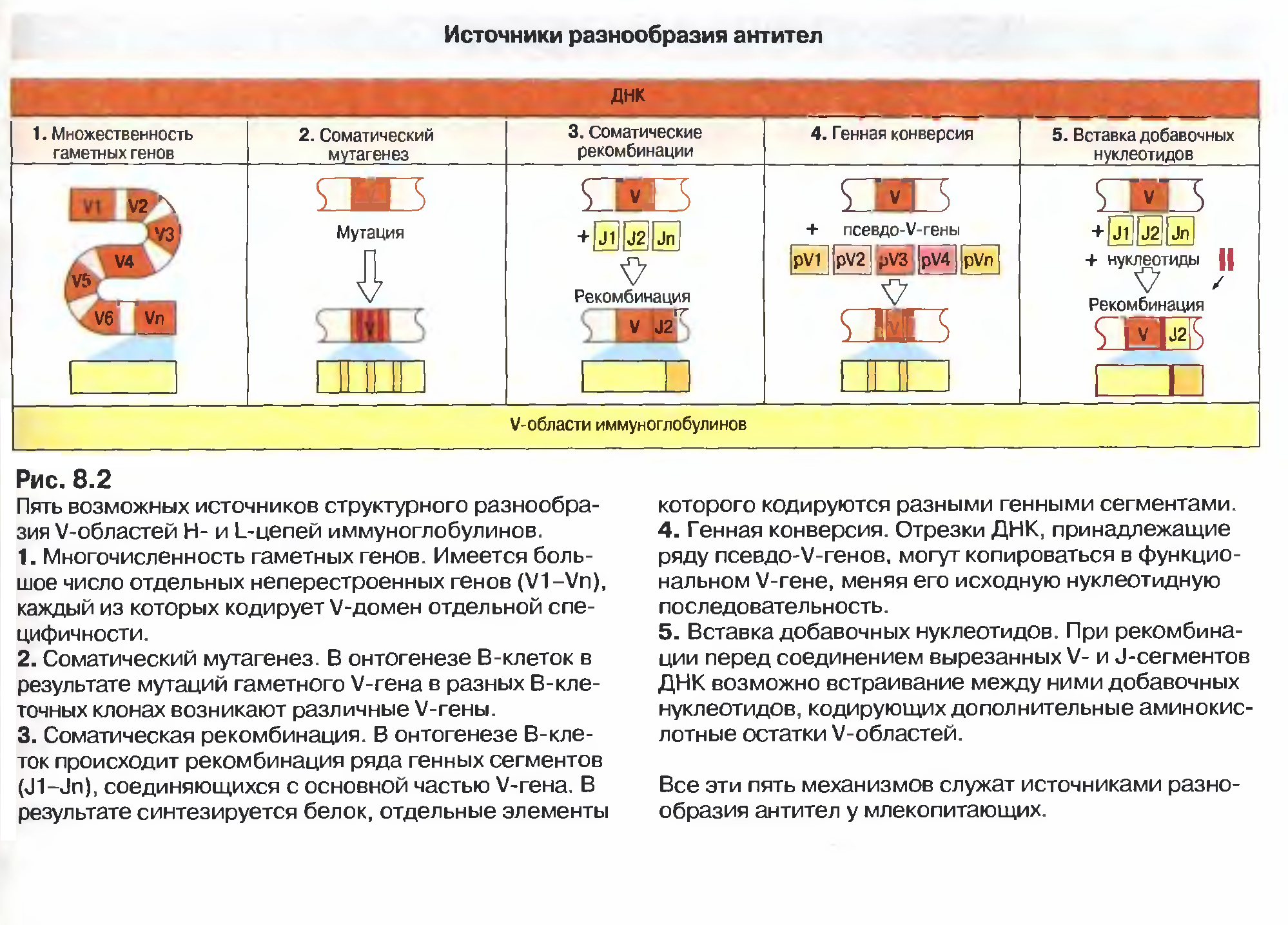
Насьогодні виділяють 6 можливих джерел різноманіття Іg :
1) множинність гаметних генів (існування копій майже кожного генного сегмента),
2) соматичні рекомбінації (урізноманітнення за рахунок сайт- специфічної рекомбінації варіабельних ділянок молекули),
3)вбудовування (інсерції) та делеції додаткових нуклеотидів=junction diversity=урізноманітнення з′єднань,
4) механізм комбінаторної диверсифікації (утворення можливих варіантів імуноглобулінів),
5)соматичний мутагенез,
6)генна конверсія (активація роботи псевдо-генів).
На рис. наведено за Ройтом можливі джерела різноманіття V областей важких і легких ланцюгів імуноглобулінів.
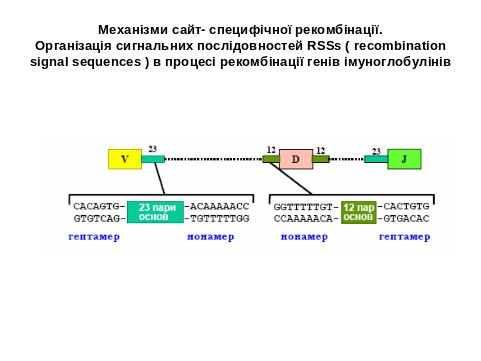
Механізм сайт- специфічної рекомбінації
Рекомбінація генних сегментів є насьогодні ключовим моментом, що формує функційні гени варіабельних ділянок важких і легких ланцюгів. Були ідентифіковані сигнальні послідовності RSSS (recombination signal sequences), які контролюють процес роботи VDJ рекомбіназ. До складу RSS входять спейсори (12 і 23 пн ), які є неконсервативними, а також гептамери (7 пн) і нонамери (9 пн)- високо консервативні ділянки ДНК. Об'єднання сегментів при рекомбінації можливо лише за умови виконання "правила 12/23", а саме- об’єднуються кодуючі сегменти, франковані RSS різних типів.
Наприклад, для важких ланцюгів згідно з рис. сигнальна послідовність нижче V і D сегментів з 3′-кінця складається з 1-гептаметра (7) та 2- спейсора з 12 н.з. та 3- сусіднього нонамера (9 )- RSS 1. Сигнальна послідовність з 5′-кінця D і J сегментів- 1-наномер; 2-спейсор 23 н.з. та 3-гептамер- RSS 2.


Рекомбінаційний процес відбувається під контролем білків, які є продуктами генів RAG1 і RAG2= рекомбіназ. Саме початок експресії цих генів є сигналом для початку перебудови генів імуноглобулінів. Крім того у цьому складному процесі рекомбінації на етапі з'єднання кінців ДНК беруть участь білки, які працюють в системі репарації двониткових розривів ДНК-
[ консервативний процес NHEJ=nonhomologous DNA end-joining = негомологічне з'єднання кінців ДНК], модифікації кінців розривів і т.д.
Це -ДНК-залежна протеїнкіназа (DNA-PK) , білок Ки (гетеродимер Ки 70:80), який асоціює з DNA-PK, ендонуклеаза Artemis, процес лігування відбувається за участі ДНК-лігази IVта ДНК-репараційного білка XRCC4 , ендо- та екзонуклеази, термінальні дезоксинуклеотидил-трансферази (TdT), полімерази та інші.

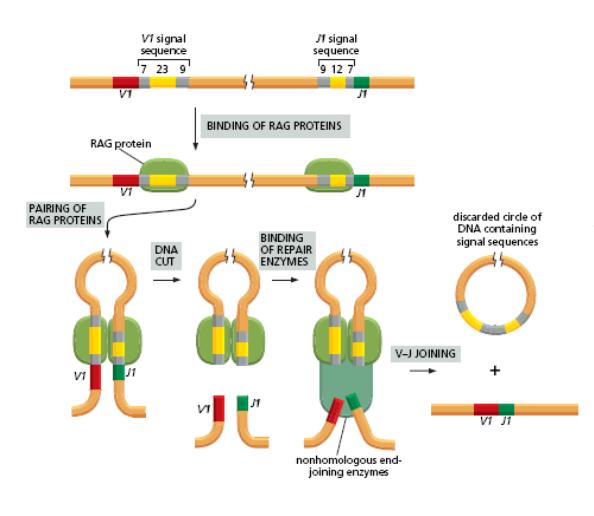
Спочатку білок RAG1 зв'язується з нонамером, а RAG2, який не здатний безпосередньо комплексуватись з ДНК, приєднується до комплексу RAG 1-ДНК поряд зі спейсером 12 н.з. Після цього між спейсорами 12-23 утворюється синапс . Таким чином, приєднання RAG1 і RAG2 до RSS стимулює їх (RAG1 і RAG2) ендонуклеазну активність, і направлене гептамерами розщеплення ДНК на кордоні між 7-гептамерами та кодуючою послідовністю (рис). Стимуляція двониткових розривів за участі RAG1 і RAG2 проходить етапи утворення nick, а потім приводить до утворення шпильковидної структури на кінці кодуючої послідовності. На першому етапі цього процесу –гідролізі- відбувається нуклеофільна атака молекули води на складно-етерний зв'язок в молекулі ДНК. Утворений на цьому етапі вільний 3"-ОН-кінець атакує фосфоетерний зв'язок –трансетерифікація- у другому ланцюгу, що сприяє утворенню шпильки- HAIRPIN- кодуючої послідовності та 5"-фосфорильованої RSS, яка після відщеплення утворює найчастіше кільцеву структуру. Рис. Для видалення шпильок ендонуклеаза Artemis надрізає один з ланцюгів ДНК у будь-якій точці шпильковидного кінця (г), розриви заповнюються шляхом синтезу ДНК і два кінця зшиваються таким чином разом (д). Розщеплення одного з ланцюгів викликає утворення паліндромних послідовностей (П=) –темний квадрат = додавання Р (palindromic sequences) нуклеотидів та збільшує довжину області з'єднання. Отже паліндромні послідовності (перевертыши)- добудовані (приєднані) нуклеотиди, які є чітко комплементарними до термінальних нуклеотидів кодуючої послідовності в районі шпильки, тому мають однакову нуклеотидну послідовність при читанні в обох ланцюгах ДНК.Додаткове різноманіття вносить робота термінальної дезокси-нуклеотидил-трансферази (TdT), яка виконує вставки додаткових нуклеотидів (за азотистими основами- найчастіше зустрічаються цитозин і гуанін)- додавання N-нуклеотидів (Non-template-encoded-не закодованих у матриці). З цим ферментом координовано взаємодіють АДА (аденозиндезаміназа) і фосфорилаза пуринових нуклеотидів.


RAG-залежна рекомбінація генів V-області імуноглобулінів (звернути увагу на специфічні ферменти і білки комплексу).
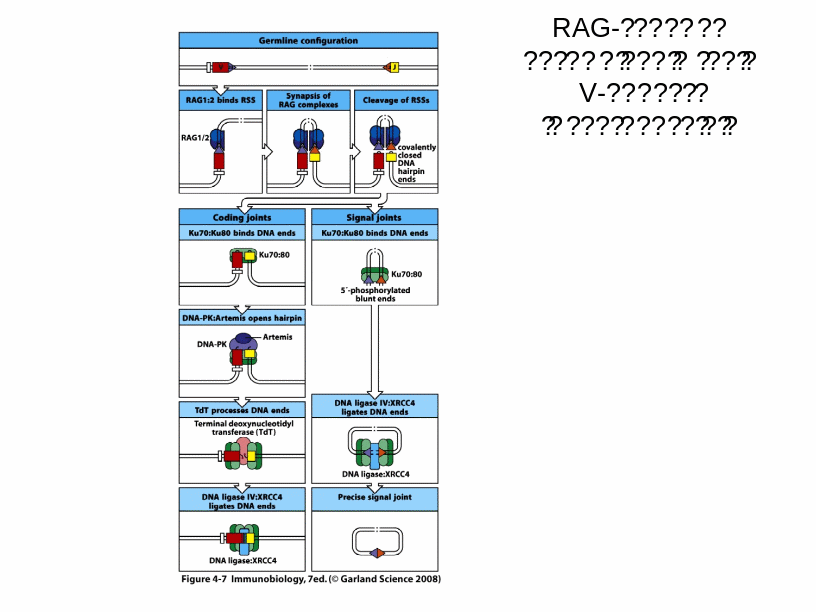
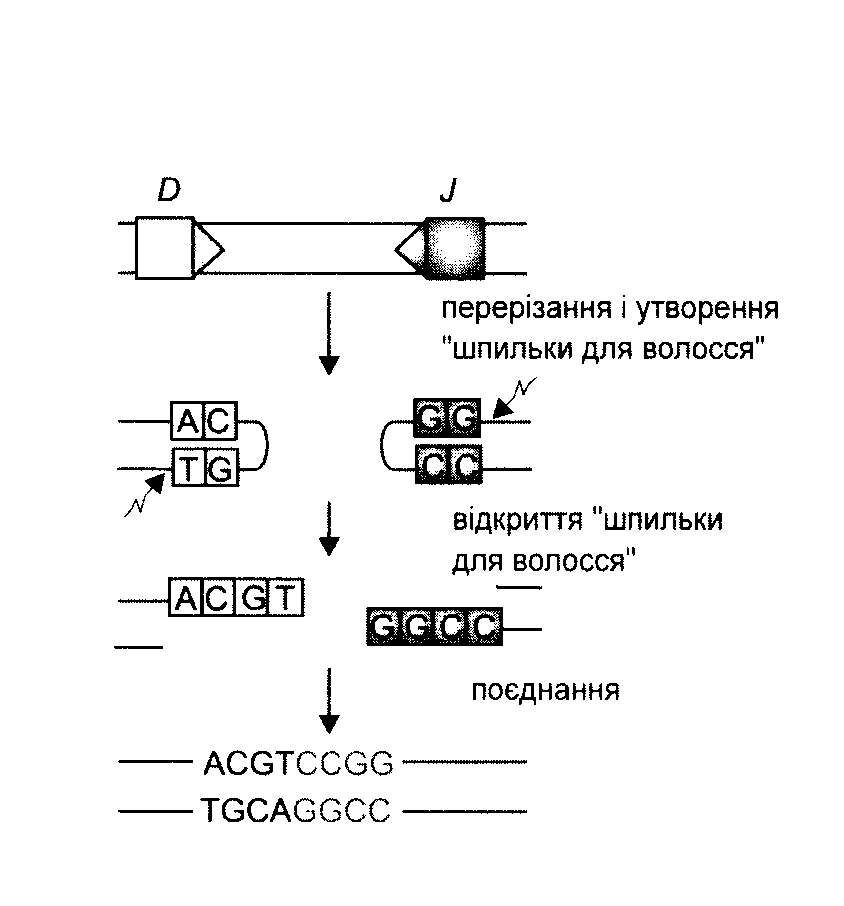
Рис. (Простіше). Схема приєднання нуклеотидів Р до рекомбінуючих генів D і J. Спочатку відбувається перерізання подвійної ДНК на межі сигнальних і кодуючих послідовностей з утворенням характерних "шпильок для волосся". Потім ці "шпильки" відкриваються в місцях, позначених ламаними стрілками. Виникають одно ланцюгові відрізки ДНК, в яких два останні нуклеотиди комплементарні до нуклеотидів початкової послідовності (нуклеотиди Р-паліндромні). Тепер може відбутись і видалення, а також приєднання поодиноких нуклеотидів (не позначене на рисунку). Після приєднання комплементарних нуклеотидів відбувається відновлення кодуючої послідовності ДНК.
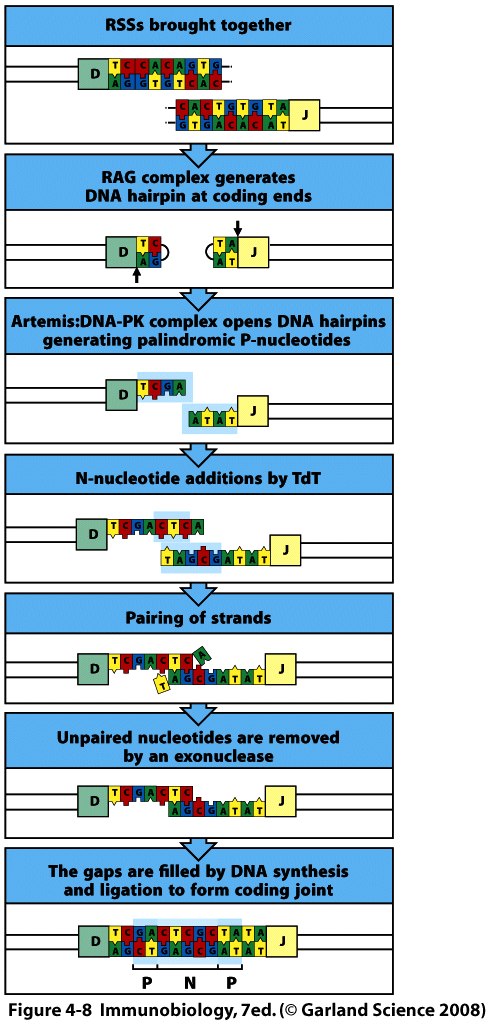
P- i N-нуклеотидні вставки при з’єднанні кодуючих сегментів генів імуноглобулінів.
Таким чином, рекомбінція запускає транскрипцію перебудованого ланцюга, що припиняє рекомбінацію інших варіантів відповідного гену і, таким чином, визначає клональний характер лімфоцитів: кожна клітина продукує антитіла тільки однієї специфічності.
Важливо відмітити, що гіперваріабільні ділянки V-доменів (CDRs) кодуються:
- на легкому ланцюгу: CDR 1, CDR 2 — на V-гені, CDR3 - на з'єднанні VJ;
- на важкому ланцюгу: CDR 1, CDR 2 - на V-гені, CDR 3 - на D-сегменті. Таким чином, у будові активного центру антитіл беруть участь продукти V-, D- і J-генів.