

Лекція №5. Гени імуноглобулінів. Біосинтез антитіл.
Довгий час великою проблемою імунології була неможливість пояснити походження великої різноманітності антитіл. Ми згадували, що таке різноманіття антитіл є неадекватним числу генів, які локалізовані в ядрі лімфоцитів. Так, в клітинах людини є загалом не більше 105 генів (250000), а вважається, що може утворюватись 1011 імуноглобулінів. Враховуючи догму молекулярної біології, один ген - один поліпептидний ланцюг, кількість генів для кодування антитіл всіх можливих специфічностей перевищувала б розмір геному хребетних. Після відкриття поліфункціональності антитіл (одне антитіло здатне зв’язуватись більше, ніж з 1 антигеном) їх розрахована кількість значно знизилась (до 106 – 108 варіантів), але все ж таки залишалась високою.
Одна з перших теорій, що пояснювала весь арсенал можливого утворення антитіл- інструктивна гіпотеза : антиген, впливаючи на гнучку молекулу Іg, формує в ній комплементарний собі центр зв’язування. Але в 60-ті роки 20 ст. ця теорія зазнала поразки, перемогла клонально –селекційна теорія, яка була висунута одночасно Ерне і Бернетом, де стверджувалось, що кожний лімфоцит утворює імуноглобуліни лише однієї специфічності, і антиген вибирає і стимулює клітини, які несуть антитіла, специфічні саме для цього антигену.
Але лишалось ще одне питання - які джерела різноманіття антитіл? Теоретично припустили, що існують свої особливі гени для антитіл кожної специфічності, але половина амінокислотної послідовності легкого ланцюга і четвертина важкого- є завжди варіабельною, а інші- константні. Як останні зберігають свій склад незмінним в разі начебто такої великої кількості генів антитіл? В 1965 році Дрейєр і Беннет вперше запропонували, що V-і С-домени імуноглобулінів кодуються різними генами, причому варіабельні домени кодуються великою кількістю генів, а константні- лише певною(невеликою) кількістю генів. А передумовою початку експресії цих генів є їх попередня перебудова (реаранжування). Справа в тому, що в геномі незрілих чи зародкових В-клітин, як і в інших соматичних клітинах, гени, що кодують Vi C домени одного ланцюга Іg, знаходяться хоч і на одній хромосомі , але є на значній відстані одне від одного ( до 100000 п.н.). Така нативна локалізація генів отримала назву- зародкова лінія (germline). При дозріванні В-клітини від про-В до зрілих плазмоцитом відбувається реорганізація геному, а саме вирізання= делетування великого об’єму генетичної інформації , закладеної в ДНК, що забезпечує зближення V i C областей і утворення повного гену – тобто єдиної інформаційної ділянки, яка кодує легкі ланцюги та повного гену, що кодує важкі ланцюги імуноглобулінів. Особливу перебудову зазнають гени варіабельних областей, які складаються з певних фрагментів, що розділені також великими генними сегментами, які делетуються в свою чергу, на етапах дозрівання В-клітин. Цей процес отримав назву реаранжування (перебудова) генів.
Рис.1. Гени імуноглобулінів. Біосинтез антитіл. 1976 р. Тонегава і співавт.-доведення можливості реаранжування генів.
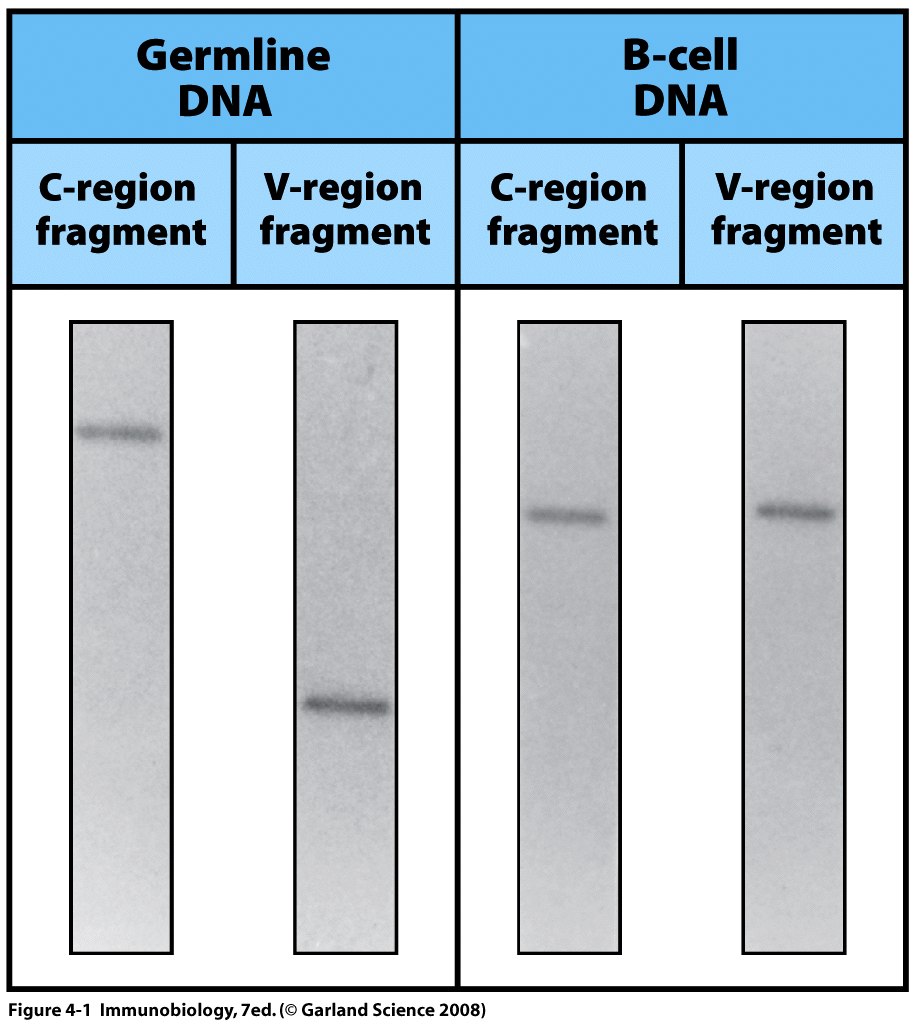
Гіпотеза, що V-і С-домени імуноглобулінів кодуються різними генами, була блискуче доведена в 1976 році .
Було знайдено, що в клітинах ембріону V- і С гени знаходяться у різних ділянках ДНК, а в ДНК зрілої мієломи вони локалізуються в одному місці.
Таким чином доведено, що в процесі онтогенезу гени імуноглобулінів підлягають перебудові (реаранжуванню), реорганізації, в результаті якої V- і С-гени зближуються і об'єднуються.
За вивчення будови генів імуноглобулінів С. Тонегава отримав Нобелівську премію 1987 року.