

Федеральное государственное автономное образовательное учреждение высшего образования «СИБИРСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ» ИНСТИТУТ ФУНДАМЕНТАЛЬНОЙ БИОЛОГИИ и БИОТЕХНОЛОГИИ
РЕФЕРАТ по ГЕНЕТИКЕ
Современные методы селекции
Преподаватель ________
Студент ___ _______ ________
Красноярск 2019
Содержание
ВВЕДЕНИЕ 3
Отбор и его творческая роль 4
Методы гибридизации 5
Мутагенез 7
Полипюидия и отдаленная гибридизация 6
Хромосомная инженерия 10
Клеточная инженерия 11
Генная инженерия 12
ЗАКЛЮЧЕНИЕ 14
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 15
Введение
Селекция (лат. selectio «выбирать») — наука о методах создания новых и улучшения существующих пород животных, сортов растений, штаммов микроорганизмов. Селекция разрабатывает способы воздействия на растения и животных с целью изменения их наследственных качеств в нужном для человека направлении. Селекцией называют также отрасль сельского хозяйства, занимающуюся выведением новых сортов и гибридов сельскохозяйственных культур и пород животных.
Основными методами селекции являются отбор, гибридизация (с использованием гетерозиса и цитоплазматической мужской стерильности), полиплоидия и мутагенез.
Бурное развитие новых методов исследований в генетике, расширение и углубление наших представлений о структуре и законах организации наследственного аппарата клетки обусловили создание и разработку принципиально новых методов. Родились новые понятия и направления современной генетики: клеточная, хромосомная инженерия и генная инженерия.
О них и пойдет речь в данном реферате.
Отбор и его творческая роль. В основе селекционного процесса лежит искусственный отбор. В сочетании с генетическими методами он позволяет создавать сорта, породы и штаммы с заранее определенными признаками и свойствами. В селекции различают два основных типа отбора: массовый и индивидуальный.
Массовый отбор — это выделение группы особей по внешним, фенотипическим признакам без проверки их генотипа. Например, при массовом, или стихийном, отборе из всей популяции кур той или иной породы в хозяйствах оставляют для размножения птиц с яйценоскостью 200—250 яиц, живой массой не менее 1,5 кг, определенной окраски, не проявляющих инстинкты высиживания и т. д. Все остальные куры выбраковываются. При этом потомство каждой курицы и петуха оценивается только по фенотипу. Следовательно, массовый отбор может дать хорошие результаты только при высоком коэффициенте наследуемости ценных признаков, избранных селекционером.
Массовый отбор наиболее эффективен в отношении качественных признаков, контролируемых одним или несколькими генами. Вместе с тем он редко бывает успешным по полигенным признакам с низким коэффициентом наследования. В этом случае необходимо применять индивидуальный, или методический, отбор.
При индивидуальном отборе (по генотипу) получают и оценивают потомство каждого отдельного растения или животного в ряду поколений при обязательном контроле наследования интересующих селекционера признаков. На последующих этапах отбора используют только тех особей, которые дали наибольшее число потомков с высокими показателями. В результате появляется возможность оценивать наследственные качества отдельных особей, т. е. способность передавать свойства потомству.
Значение индивидуального отбора особенно велико в тех отраслях сельскохозяйственного производства, где имеется возможность получения от одного организма большого количества потомков. Так, используя искусственное осеменение, от одного быка можно получить до 35000 телят с помощью глубокого замораживания семени, сохраняющегося долгие годы. Поэтому уже теперь во многих странах мира существуют банки спермы животных с ценными генотипами. Отбор в селекции отличается наибольшей эффективностью в том случае, если он сочетается с определенными типами скрещиваний.
Методы гибридизации (типы скрещивания) в селекции. Все разнообразие типов скрещиваний сводится к инбридингу и аутбридингу. Инбридинг — это близкородственное (внутрипородное или внутрисортовое), а аутбридинг — неродственное (межпородное или межсортовое) скрещивание.
При инбридинге в качестве исходных форм используются братья и сестры или родители и потомство (отец — дочь, мать — сын, двоюродные братья — сестры и т. д.). Этот тип скрещивания применяют в тех случаях, когда желают перевести большинство генов породы в гомозиготное состояние и, как следствие, закрепить хозяйственно ценные признаки, сохраняющиеся у потомков. Такое скрещивание в определенной степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности.
Вместе с тем при инбридинге часто наблюдается ослабление животных, их постепенное вырождение, обусловленное гомозиготизацией рецессивных аллелей. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственные скрещивания используют при инбридинге. Для избежания этого явления необходимо проводить строгий отбор особей, обладающих ценными хозяйственными признаками.
У растений чистые линии также обладают пониженной жизнеспособностью, что, вероятно, связано с переходом в гомозиготное состояние всех рецессивных мутаций, которые в основном являются вредными.
Чистые линии, полученные в результате инбридинга, отличаются не только различными признаками, но и степенью снижения жизнеспособности. Если эти чистые линии скрещивать между собой, то обычно наблюдается эффект гетерозиса.
Неродственное скрещивание между особями одной породы или между особями разных пород (кроссбридинг) животных позволяет поддерживать свойства или улучшать их в ряду следующих поколений гибридов.
При аутбридинге скрещивают организмы, относящиеся к разным линиям внутри сорта (или породы), к разным сортам (или породам), к разным видам или даже родам. В двух последних случаях речь идёт об отдаленной гибридизации. Скрещивание организмов, относящихся к разным видам и тем более родам, крайне сложно, т.к. родительские организмы отличаются по генетическому материалу, физиологическим или морфологическим признакам. Очень часто такое скрещивание приводит к образованию бесплодных (т.е. не дающих потомство) гибридов. Связано это с тем, что в гибридном организме встречаются родительские хромосомы, столь несхожие между собой морфологически, а часто и по числу, что они не способны к конъюгации, это ведет к нарушениям процесса мейоза, а, следовательно, и не формируются нормальные половые клетки (гаметы). Однако способность растений к вегетативному размножению, а также разработка специальных методов преодоления бесплодия у отдаленных гибридов в ряде случаев делает возможным формирование новых форм, обладающих определенными ценными качествами, характерными для разных видов.
Мутагенез. Бывает двух типов: cпонтанный и индуцированный мутагенез. Спонтанные мутанты используются преимущественно в селекции растений. Так, на основе мутанта желтого безалкалоидного люпина получено несколько сортов сладкого люпина, которые выращивают на корм скоту. Люпин, содержащий алкалоиды, для этой цели непригоден, поскольку животные его не едят.
Большое число мутантов известно у плодовых культур, которые используются как новые сорта или в гибридизации с другими формами. Один из наиболее известных спонтанных мутантов кукурузы opaque, отличающийся высоким содержанием аминокислоты лизина в зерне, используется для создания так называемых высоколизиновых гибридов кукурузы.
В последние десятилетия во многих странах мира развернуты работы по получению индуцированных мутантов. Индуцированные рентгеновыми лучами мутанты были выделены у многих злаков (ячменя, пшеницы, ржи и др.). Они отличаются не только повышенной урожайностью, но и укороченным побегом. Такие растения устойчивы к полеганию и имеют заметные преимущества при машинной уборке. Кроме того, короткая и прочная соломина позволяет вести дальнейшую селекцию на увеличение размера колоса и массы семян без опасения, что повышение урожая зерна приведет к полеганию растений.
Особенно успешно индуцированный мутагенез применяют в селекции микроорганизмов.
Полипюидия и отдаленная гибридизация. При создании новых сортов растений селекционерами широко используется метод автополиплодии, который приводит к увеличению размеров клеток и всего растения вследствие умножения числа наборов хромосом. Кроме того, избыток хромосом повышает их устойчивость к патогенным организмам (вирусам, грибам, бактериям) и ряду других неблагоприятных факторов, например к радиации: при повреждении одной или даже двух гомологичных хромосом остаются неповрежденными другие такие же. Полиплоидные особи жизнеспособнее диплоидных.
Около 80 % современных культурных растений являются полиплоидами. Среди них хлебные злаки, овощные и плодово-ягодные культуры, цитрусовые, технические, лекарственные и декоративные растения, которые гораздо более урожайны, чем исходные диплоидные сорта. Так, триплоидная сахарная свекла отличается от обычной не только большей урожайностью вегетативной массы и более крупными размерами корнеплодов, но и повышенной их сахаристостью, а также устойчивостью к болезням. Однако триплоиды стерильны, поэтому необходимо каждый раз получать гибридные семена от скрещивания диплоидной и тетраплоидной форм. Успешному решению этой проблемы способствовало открытие мужской стерильности свеклы. Стерильность триплоидных гибридов может иметь положительное значение при получении бессемянных плодов, например винограда и арбуза.
Ценные результаты дает использование в селекции явления аллополиплоидии, основой которого служит метод отдаленной гибридизации, т. е. скрещивания организмов, относящихся к разным видам и даже родам. Например, получены межвидовые полиплоидные гибриды капусты и редьки, ржи и пшеницы. Гибридизация пшеницы (Triticum) и ржи (Secale) позволила получить ряд форм, объединенных общим названием тритикале. Они обладают высокой урожайностью пшеницы и зимостойкостью, и неприхотливостью ржи, устойчивостью ко многим болезням, в том числе к линейной ржавчине, являющейся одним из главных факторов, ограничивающих урожайность пшеницы.
На основе гибридизации пшеницы и пырея российским академиком Н. В. Цициным получены пшенично-пырейные гибриды, обладающие высокой урожайностью и устойчивостью к полеганию. Однако отдаленные гибриды, как правило, бесплодны. Это связано с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. Для восстановления плодовитости у межвидовых гибридов в 1924 г. советский генетик Г. Д. Карпеченко предложил использовать удвоение числа хромосом (полиплоидию) у отдаленных гибридов.
Г. Д. Карпеченко проводил скрещивание редьки и капусты. Число хромосом у этих растений одинаково. Соответственно их гаметы несут по 9 хромосом. Гибрид капусты и редьки имеет 18 хромосом, но он бесплоден, так как хромосомы капусты и редьки в мейозе не конъюгируют, поэтому процесс образования гамет не может протекать нормально. В результате удвоения числа хромосом в бесплодном гибриде оказалось 36 хромосом, слагающихся из двух полных диплоидных наборов редьки и капусты. Это создало нормальные возможности для мейоза; хромосомы капусты и хромосомы редьки конъюгировали между собой. Каждая гамета несла по одному гаплоидному набору редьки и капусты (9 + 9 = 18). В зиготе вновь оказалось 36 хромосом; межвидовой гибрид стал плодовитым. По фенотипу этот новый растительный организм совмещал признаки редьки и капусты, например в строении стручка.
Получение экспериментальным путем полиплоидных животных представляет большую трудность, поэтому такие формы животных — редкость. Так, советскому ученому генетику Б. Л. Астаурову путем межвидовой гибридизации удалось получить полиплоидную форму тутового шелкопряда. На сегодняшний день есть уже полиплоидные рыбы, птицы (например, куры), однако внедрение полиплоидных пород животных в практику сельского хозяйства — дело будущего.
Бурное развитие новых методов исследований в генетике, расширение и углубление наших представлений о структуре и законах организации наследственного аппарата клетки обусловили создание и разработку принципиально новых методов. Родились новые понятия и направления современной генетики: клеточная, хромосомная инженерия и генная инженерия. При этом принципиальное отличие данных методов от традиционно используемых в селекции, состоит в целенаправленном, а не случайном расширении границ изменчивости генотипа, в планируемом разнообразии исходного материала для селекции. Эти современные методы большее применение пока получили в селекции растений. Клеточная инженерия связана с культивированием отдельных клеток или тканей на специальных искусственных средах. Показано, что если взять кусочки ткани или отдельные клетки из разных органов, допустим, растений, хотя это возможно и у животных, и пересадить их на специальные среды, содержащие минеральные соли, аминокислоты, гормоны и другие питательные компоненты, то они способны расти. Это значит, что в таких изолированных от организма тканях и клетках продолжаются клеточные деления. Но самым важным и интересным оказалось то, что отдельные растительные клетки (в отличие от клеток животных) в таких искусственных условиях обладают тотипотентностью, т.е. способны к регенерации (формированию) полноценных растений. Эта их способность и была использована для селекции в разных направлениях.
Хромосомная инженерия. В настоящий момент связывается прежде всего с возможностями замещения (замены) отдельных хромосом у растений или добавления новых. Известно, что в клетках каждого диплоидного организма имеются пары гомологичных хромосом. Такой организм называют дисомиком. Если в какой-либо паре хромосом остается одна гомологичная хромосома, то получается моносомик. При добавлении третьей гомологичной хромосомы возникает трисомик, а при отсутствии в геноме одной пары гомологичных хромосом возникает нуллисомик. Такие манипуляции с хромосомами дают возможность заменять одну или обе гомологичные хромосомы, допустим, одного сорта пшеницы на ту же пару хромосом, но из другого сорта. Тем самым он может один признак, который ему кажется слабым у данного сорта, заменить на этот же, но более сильный признак из другого сорта. Таким образом, он приближается к созданию «идеального» сорта, у которого все полезные признаки будут выражены в максимальной степени. Эту же цель преследует и методика замены отдельных хромосом одного вида на хромосомы другого вида, близкого по своему происхождению.
Клеточная инженерия. Основана на культивировании отдельных клеток или тканей на искусственных питательных средах. Такие клеточные культуры используются для синтеза ценных веществ, производство незараженного посадочного материала, получения клеточных гибридов. Метод гибридизации клеток приобретает все большее значение в селекции. Оказалось, что если взять клетки разных органов и тканей или клетки разных организмов, объединить их с помощью специальных приемов, разработанных учеными в одну, то образуется новая, гибридная клетка. Свойства этой клетки существенно отличаются от свойств родительских клеток. Таким путем можно получать клетки, выделяющие необходимые человеку лекарства.
Успехи клеточной инженерии могут открыть новые возможности в селекции животных. В 1997 г. научная общественность была взбудоражена сообщением, что в Англии были проведены успешные эксперименты по генетическому клонированию овцы. Для этого использовали ядра соматических клеток, полученных из ткани молочной железы взрослой овцы. Из яйцеклетки удалялось ядро и замещалось ядром соматической клетки. Образовавшуюся диплоидную зиготу стимулировали к дроблению электрошоком и трансплантировали в овцу-реципиента. Через 148 дней приемная мама родила живую овечку, её назвали Долли.
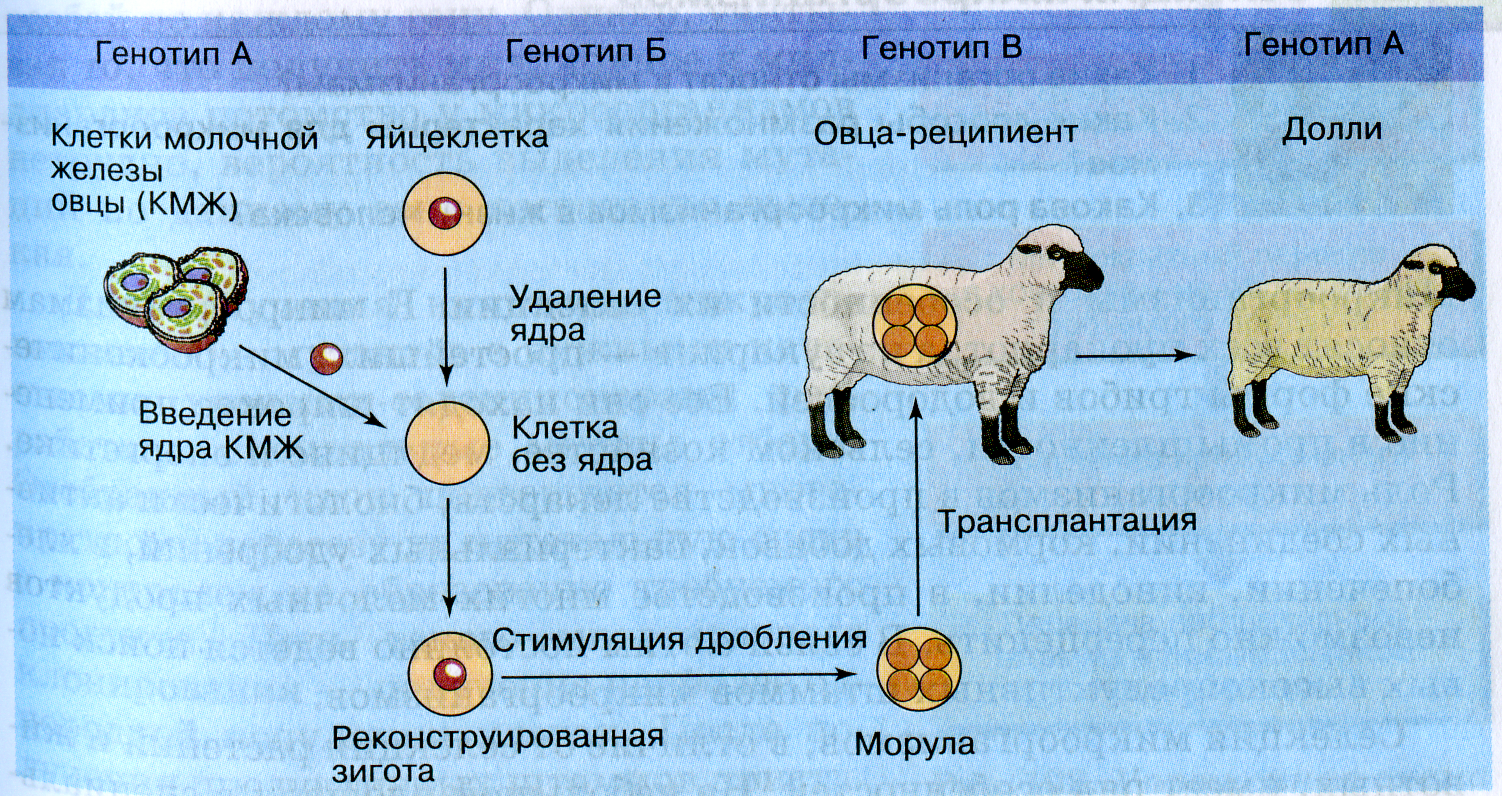
Рис.1. Схема генетического клонирования овцы
Генная инженерия. Под генной инженерией обычно понимают искусственный перенос нужных генов от одного вида живых организмов (бактерий, животных, растений) в другой вид, часто очень далекий во своему происхождению. Чтобы осуществить перенос генов (или трансгенез), необходимо выполнить следующие сложные операции: выделение из клеток бактерий, животных или растений тех генов, которые намечены для переноса. Иногда эту операцию заменяют искусственным синтезом нужных генов, если таковой оказывается возможным; создание специальных генетических конструкций (векторов), в составе которых намеченные гены будут внедряться в геном другого вида. Такие конструкции кроме самого гена должны содержать все необходимое для управления его работой (промоторы,терминаторы) и гены-«репортеры», которые будут сообщать, что перенос успешно осуществлен;внедрение генетических векторов сначала в клетку, а затем в геном другого вида и выращивание измененных клеток в целые организмы (регенерация). Растения и животные, геном которых изменен в результате таких генно-инженерных операций, получили название трансгенных растений или животных.