

5.6. Решение:
1). р атм = 740 мм рт ст = 98658,28 Па (табличные данные)
р изб = 3 кгс/см2 = 294300 Па
р абс (рабочее) = р атм + р изб = 98658,28 + 294300 = 0,4 мПа = 4 кгс/см2
2). Q – теплота, необходимая для осуществления процесса
Q = cб/м ∙ ρб/м ∙ V ∙ (t2-t1) = 4000∙ 950∙ 5∙ 45 = 855 ∙ 106 Дж
3). При рабочем давлении 4 кгс/см2 удельная теплота конденсации r = 2141 кДж/кг
Q = G ∙ r, где G – количество водяного пара
G = Q/ r = 855 ∙ 106/2141000 = 400 кг.
6.1. Хемостатные кривые при лимитировании процесса различными субстратами и кислородом.
Графические зависимости между установившимися значениями X и S в хемостатном процессе и скоростью разбавления D называют хемостатными кривыми (рис. 9.5).

Хемостатная кривая X(D) имеет особенность: концентрация биомассы снижается до нуля при повышении скорости разбавления до некоторого критического значения и при более высоких скоростях разбавления остается равной нулю. Физически это означает, что культура вымывается. Выходит больше биомассы, чем может вырасти.
Определим, при каких значениях Dкр происходит вымывание культуры:


Отсюда следует, что вид хемостатной кривой зависит от S0. При D = 0
X = YXSS0
Таким образом, и начальное положение хемостатной кривой, и точка вымывания зависят от концентрации субстрата в свежей среде (подпитке) S0. Эта величина наряду со скоростью разбавления является параметром, с помощью которого оператор может управлять процессом. Чем больше S0, тем выше концентрация биомассы в выходном потоке, причем в довольно широком диапазоне скоростей разбавления концентрация биомассы сохраняется почти на постоянном высоком уровне, лишь немного снижаясь с возрастанием скорости разбавления.
Интересную особенность имеет зависимость стационарной (остаточной) концентрации субстрата от начальной (входной) его концентрации. В уравнении (9.19) нет ничего, кроме D и кинетических констант mum и KS Из этого следует парадоксальный вывод: при любом изменении концентрации субстрата во входящем потоке S0 в стационарном состоянии при заданной скорости разбавления устанавливается одна и та же остаточная концентрация субстрата S. Именно это свойство хемостата дало ему название: концентрация субстрата (химического соединения) стабилизируется сама по себе независимо от колебаний на входе.
Лимитирование субстратами.
Хемостатные кривые X(D) чувствительны к изменению кинетических зависимостей, происходящему при лимитировании процесса различными субстратами.
Углерод. Отклонение от классической хемостатной кривой (снижение концентрации биомассы при малых скоростях разбавления) связано с повышенным расходом субстрата на поддержание жизнедеятельности микроорганизмов при малых удельных скоростях роста биомассы. Если решить систему уравнении (9.13), (9.14) с учетом затрат на поддержание жизнедеятельности, получится хемостатная кривая, приведенная на рис. 9.8, а.

Азот или сера. Наблюдаемые отклонения отражают превышение концентрации биомассы при малых скоростях разбавления по сравнению с идеальным хемостатом (рис. 9.8, б). Клетки при малой скорости роста накапливают резервные соединения, и по массе их как бы становится больше. Такими соединениями могут быть полисахариды, полиоксибутират, липиды. Такой режим лимитирования в обиходе называют «жировым». Если выражать концентрацию биомассы не в г/л, а в числе клеток на литр, получается обычная форма графика.
Магний, фосфор, калий. Подвиду хемостатная кривая (рис. 9.8, в) похожа на кривую для азота, но причина подобного отклонения здесь другая: клетки, растущие медленно, требуют меньше РНК, чем быстрорастущие.
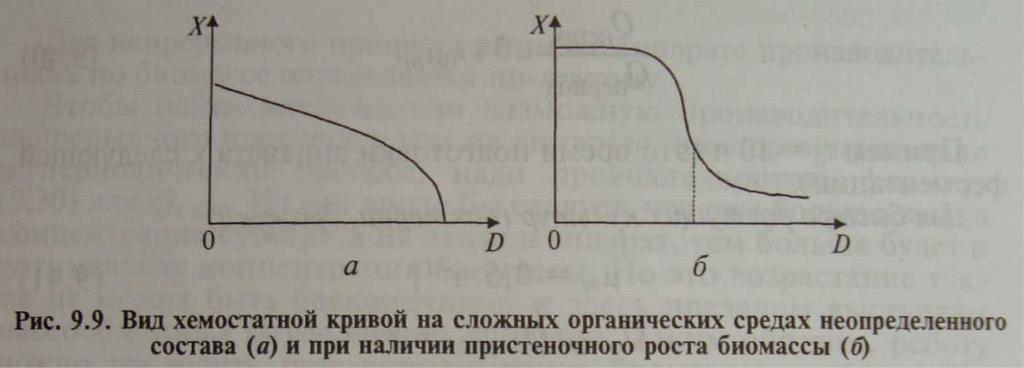
Сложная среда. На неопределенных по составу питательных средах происходит при различных скоростях разбавления изменение лимитирования (переход с лимитирования одним субстратом на лимитирование другим). Хемостатная кривая (рис. 9.9, а) не имеет участка плато, а все время снижается с повышением скорости разбавления. При этом изменяется и биохимический состав биомассы.
Пристеночный рост биомассы. Хемостатная кривая в этом случае (рис. 9.9, б) имеет аномально длинный «хвост», рост биомассы возможен даже при D > mum (так как растущая на стенках биомасса не вымывается).
При использовании сложного субстрата, когда лимитация осуществляется различными количествами источников хемостатная кривая не имеет ярко выраженного плато.
В аэробных условиях при больших концентрациях биомассы возможна смена лимитирующих факторов (с углерода (С) на кислород и т.п.).
