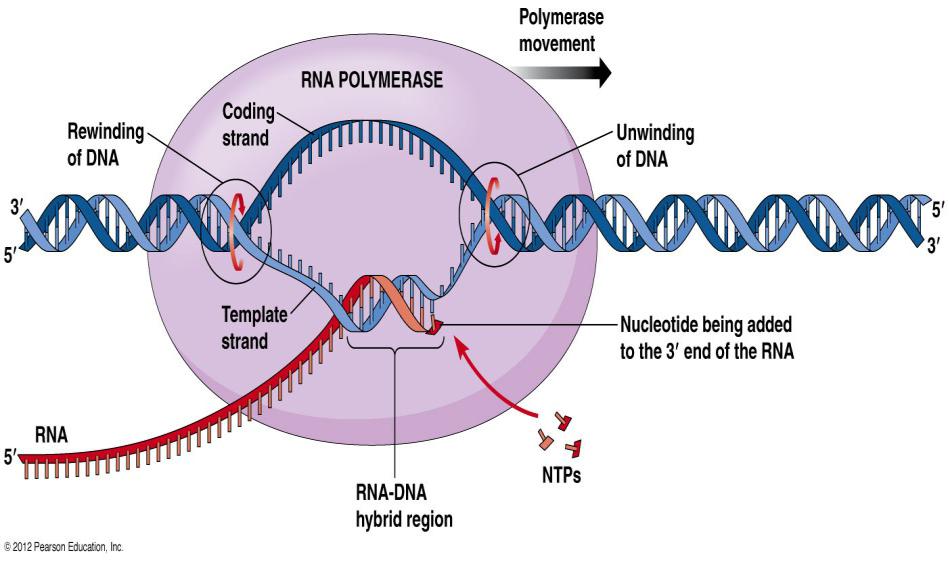
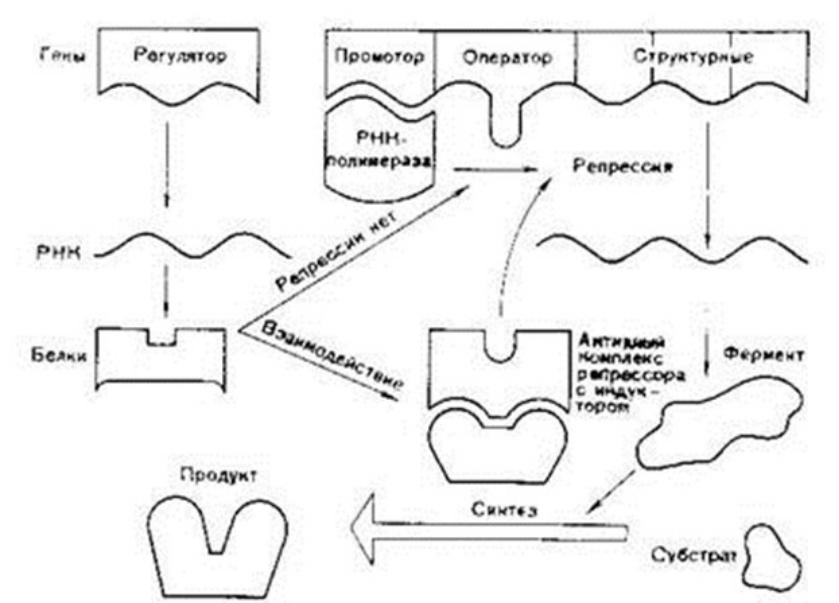
У эукариот имеются три типа РНК-полимераз:
РНК-полимераза1 - катализирует транскрипцию рибосомальных генов.
РНК-полимераза2 - катализирует транскрипцию всех структурных генов.
РНК-полимераза3- катализирует транскрипцию тРНК и 5S-рибосомальной РНК (катализирует образование мяРНК, присутствующие только у эукариот).
Промоторный участок (промотор – участок присоединения РНК-полимеразы) у эукариот более длинный.
Здесь три последовательности:
одна проксимальная и две дистальные.
В проксимальной основной мотив ТАТА. У эукариот это бокс Хогнеса-Гольдберга,
у прокариот – Бокс Прибнова (ТАТААТ в
проксимальном положении, ТТGАСА – в дистальном положении).
Особенность дистальной последовательности заключается в том, что с одной стороны они имеют GC-основания и образуют шпильку посредством инвертированного повтора, и освобождается транскрибируемая нить РНК. Терминирующая последовательность содержит мотив ААУААА. Терминирующая последовательность расположена у конца, представленным поли-А.
Регуляция у эукариот
1)Регуляция генной активности у эукариот намного сложнее, чем у бактерий. У эукариот она происходит не только на уровне клетки.
Существуют системы регуляции организма как
целого. Огромную роль в регуляции играют гормоны, но регулируют они процессы синтеза белков лишь в клетках-мишенях.
Гормоны связываются с белками-рецепторами,
расположенными в мембранах таких клеток и включают системы изменения структуры клеточных белков в печени.
Гормоны могут влиять как на процессы транскрипции, так и процессы трансляции.
Каждый гормон через систему посредников активирует свою группу генов.
Так адреналин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а инсулин влияет на образование гликогена из глюкозы в печени
Различают экспрессию гена:
1) конститутивную – происходящую в
клетке независимо от внешних обстоятельств. Сюда относят экспрессию генов, определяющих синтез макромолекул, необходимых для жизнедеятельности всех клеток, и
спец. генов (тканеспецифичная экспрессия
гена), характерных для конкретного вида клеток.
2) Индуцибельная экспрессия гена
определяется действием каких-либо агентов - индукторов. Ими м. б. гормоны, ростовые вещества и вещества, определяющие дифференцировку клеток (напр., ретиноевая кислота). Индукция может происходить:
на определенной стадии развития организма,
в определенной ткани;
время и место индукции регулируются геномом.
Изменения в экспрессии гена носят необратимый характер, по крайней мере в нормальных клетках. У раковых и трансформированных клеток эта закономерность может нарушаться.
В роли индукторов м. б. также и факторы внешней
среды, например изменение температуры, питательные вещества. После прекращения действия индуктора первоначальная картина экспрессии гена восстанавливается (временная экспрессия гена).
Большое значение экспрессия гена имеет в оптимизации синтеза белков методами генетич. инженерии.
В качестве продуцента используют бактерии, дрожжи, растительные и животные клетки и даже живые организмы, такие организмы называют
трансгенными.
Искусственные гены конструируются таким образом, чтобы получить макс. кол-во желаемого продукта с миним. затратами, другими словами, чтобы достичь максимально высокого уровня экспрессии активного белка.
Для сильной экспрессии в искусств, гене используют "сильные" регуляторные последовательности генов, обеспечивающие наибольшую продукцию белка.
Часто эти последовательности ДНК имеют вирусное происхождение.
Описаны случаи экспрессии целевого продукта в бактериях до уровня 50% от всего клеточного белка,
Как правило, суперэкспрессированные белки нерастворимы и секретируются в периплазматич. пространство бактерии. Особую сложность представляет получение белков, токсичных для клетки. В таких случаях используют строго индуцибельные системы (напр., РНК-по-лимеразу фага Т7 и ген с промотором для нее) или системы, позволяющие быстро выводить продукт наружу (секретирую-щие системы