

Глава IV. Корень и корневые системы
КОРЕНЬ
Функции корня. Корень — основной вегетативный орган растения, выполняющий функцию почвенного питания. Корень — осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не возникают листья, а апикальная меристема всегда прикрыта чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции:
а) корни укрепляют растения в почве, делают возможным вертикальный рост и вынесение побегов вверх;
б) в корнях синтезируются различные вещества (многие аминокислоты, гормоны, алкалоиды и пр.), которые затем передвигаются в другие органы растения;
в) в корнях могут откладываться запасные вещества;
г) корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве.
У многих растений корни выполняют особые функции.
Эволюционное возникновение корня. Тело риниофитов (см. введение) еще не было расчленено на побеги и корни. Оно состояло из осевых органов, которые нарастали в длину с помощью апикальных меристем и ветвились дихотомически. Одни из ответвлений поднимались вертикально, а другие прижимались к почве и поглощали воду и минеральные соли. Несмотря на примитивное строение, эти растения были обеспечены водой, так как имели небольшие размеры, жили у воды и в воде, а климат был влажным.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и дали начало корням, приспособленным к более совершенному почвенному питанию. Возникновение корней как специализированных органов почвенного питания сопровождалось глубокой перестройкой их структуры и появлением специализированных тканей.
Функцию поглощения веществ из почвы приняли на себя молодые окончания, сохраняющие на поверхности жизнедеятельные клетки. Эти клетки образовали важнейшую в функциональном отношении ткань — ризодерму. Общая поглощающая поверхность корней увеличилась благодаря:
а) обильному ветвлению и образованию большого числа всасывающих окончаний;
б) постоянному нарастанию корней и передвижению всасывающих окончаний в новые участки почвы; в) образованию корневых волосков;
г) образованию новых придаточных корней.
Защиту апикальной меристемы от повреждений при нарастании корней в плотной почве обеспечило возникновение чехлика.
Образование корней было крупным эволюционным достижением, благодаря которому растения смогли осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх, к свету.
Зоны молодого корневого окончания.
Нежные клетки апикальной меристемы, словно живым наперстком, всегда прикрыты чехликом (рис.1). Лишь в редчайших случаях (у некоторых водных растений и паразитов) чехлик отсутствует. Чехлик состоит из живых клеток и постоянно обновляется: по мере того как с его поверхности слущиваются старые клетки, на смену им, изнутри, апикальная меристема образует новые молодые клетки.
Слущивание наружных клеток чехлика нельзя представлять как результат простого механического стирания их по мере продвижения среди твердых частиц почвы. Наружные клетки отслаиваются от поверхности чехлика еще будучи живыми. Они продуцируют обильную слизь, которая облегчает продвижение. Центральная (осевая) часть чехлика составляет, так называемую, колумеллу (к о л о н к у), в клетках которой содержится много крахмальных зерен. Предполагают, что эти зерна служат статолитами (греч. статос — стоящий; литос — камень), т. е. способны перемещаться в клетке при изменении положения кончика корня в пространстве, благодаря чему корень изгибается и растет в прежнем направлении.
Под чехликом находится зона деления, клетки которой имеют характер меристемы (рис. 1)Почти все делящиеся клетки сосредоточены в этой зоне, имеющей длину менее 1 мм. Зону деления часто можно отличить на живом корешке по желтоватой окраске, зависящей от того, что клетки меристемы заполнены цитоплазмой и не имеют заметных вакуолей.
Вслед за этой зоной корень остается гладким, но становится светлым и как бы прозрачным. Это — зона растяжения (рис. 89, 2). В этой зоне клетки сильно увеличиваются в продольном направлении (вдоль оси корня), однако клеточные деления уже почти отсутствуют и объем корня увеличивается
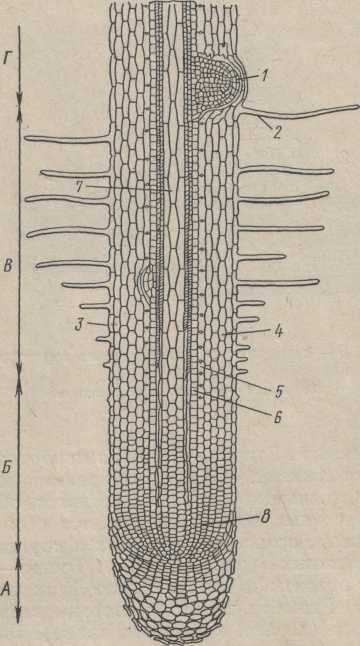
Рис.1. Кончик корня
(схема продольного среза). А — корневой
чехлик; Б — зона роста и растяжения; В
— зона корневых волосков, или всасывающая;
Г — начало зоны проведения
/ — начало роста
бокового корня, 2 — корневые 'волоски,
3 — эпиблема, 4 — кора корня, 5 —
эндодерма, 6 ~— перицикл, 7 — центральный
цилиндр, 8 — меристема
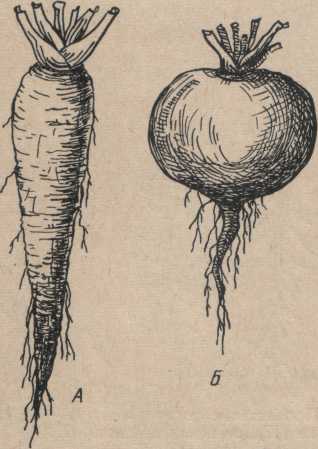
Классификация корней. Корни очень разнообразны как по способу ветвления, так и по морфологии (форме), происхождению и экологическим типам.
Рис.
2. Формы корней: А — веретеновидный
корень; Б — реповидный корень
Формы корней исключительно многообразны: шнуровидные, бичевидные, конусовидные, веретеновид- ные, реповидные, клубневидные и др. (рис. 2).
По отношению к субстрату, в котором корни размещаются, различают четыре экологических типа корней: подземные, водяные (или плавающие), воздушные и гаустории (присоски растений-паразитов) (рис. 2, 3).

Рис.3. Водяные корни лягушатника и воздушные корни у эпифитных орхидей.
рен для большинства двудольных растений. Вторая — система придаточных корней, или мочковатая корневая система, — может развиваться из любой части стебля или листа. Придаточные корни также ветвятся, образуя боковые корни. Система придаточных корней развивается обычно у однодольных, например, у злаковых, осоковых, лилейных и других, редко у двудольных, например лютиковых (рис. 4, А, Б). Третья — смешанная корневая система, когда развиты и одновременно функционируют обе предыдущие (рис. 4, В). Размеры корневой системы зависят как от жизненной формы растений (дерево, кустарник, трава), так и от внешних условий. У водных и болотных растений корневая система слабо развита. Растения засушливых местообитаний, например степных или пустынных, развивают очень мощную корневую систему.
Приведенная классификация не охватывает всего разнообразия корней. Так, например, выделяют особую группу сокращающихся, или контрактильных корней, а также корневые мочки. Контрактильные корни втягивают в почву корневую шейку и основания побегов с почками возобновления. Они хорошо выражены у клевера, гречихи и др. Корневые мочки— это кратковременные нитевидные корни, развивающиеся близко к поверхности почвы в связи с сезонным увлажнением или поливом.
б
рен для большинства двудольных растений. Вторая — система придаточных корней, или мочковатая корневая система, — может развиваться из любой части стебля или листа. Придаточные корни также ветвятся, образуя боковые корни. Система придаточных корней развивается обычно у однодольных, например, у злаковых, осоковых, лилейных и других, редко у двудольных, например лютиковых (рис. 4, А, Б). Третья — смешанная корневая система, когда развиты и одновременно функционируют обе предыдущие (рис. 4, В). Размеры корневой системы зависят как от жизненной формы растений (дерево, кустарник, трава), так и от внешних условий. У водных и болотных растений корневая система слабо развита. Растения засушливых местообитаний, например степных или пустынных, развивают очень мощную корневую систему.
Приведенная классификация не охватывает всего разнообразия корней. Так, например, выделяют особую группу сокращающихся, или контрактильных корней, а также корневые мочки. Контрактильные корни втягивают в почву корневую шейку и основания побегов с почками возобновления. Они хорошо выражены у клевера, гречихи и др. Корневые мочки— это кратковременные нитевидные корни, развивающиеся близко к поверхности почвы в связи с сезонным увлажнением или поливом.
Метаморфозы корней. Часто корни выполняют особые функции, и в связи с этим у них меняется строение. Если корни изменены сравнительно мало, то их морфологическая природа легко устанавливается. В крайних же случаях строение изменено так сильно, что для выяснения морфологической природы требуется специальное исследование, и тогда говорят, что корни метаморфизированы. Под метаморфозом понимают резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций. Видоизменения корней очень разнообразны.
Микориза и сожительство с бактериями. Корни многих растений сожительствуют с почвенными грибами. Корневые окончания и сросшиеся с ними гифы грибов образуют микоризу (дословно «грибокорень»). Высшее растение и гриб извлекают из такого сожительства взаимную пользу, т. е. находятся в состоянии симбиоза. Грибной компонент облегчает корням восприятие воды и минеральных веществ из почвы и, по-видимому, передает им некоторые органические вещества. В свою очередь, гриб получает от высшего растения углеводы и другие питательные вещества. Симбиотические отношения не исключают того, что гриб на некоторых этапах развития угнетает высшее растение и паразитирует на нем, а высшее растение в известные моменты «переваривает» грибные гифы, находящиеся в нем. Словом, этот симбиоз можно рассматривать как хорошо отрегулированный паразитизм.
Грибные гифы проникают внутрь сосущих корешков, растут по межклетникам и проникают в клетки коры в зоне поглощения. Апикальная меристема остается свободной от гриба, так же как и ростовые корни. Если гифы грибов образуют чехол, окутывающий корень снаружи, то говорят об эктомикоризе. Этот тип более характерен для деревьев и кустарников. Если все гифы находятся внутри корня, как у большинства травянистых растений, то микоризу называют эндомико ризой.
Микотрофное питание (т. е. питание с помощью грибов), видимо, возникло сотни миллионов лет назад у примитивных сухопутных растений, у которых еще не было корней. В настоящее время микотрофия распространена очень широко. Большая часть дикорастущих и культурных трав и деревьев образует микоризу. Некоторые растения, например орхидные, вообще не могут жить без симбиоза с грибами.
Бактериальные клубеньки на корнях бобовых, видимо, представляют собой измененные боковые корни, приспособленные к симбиозу с бактериями из рода Rhizobium. Эти бактерии проникают через корневые волоски внутрь молодых корней и вызывают образование на них клубеньков.
 Между клетками
корня и бактериями существует тесное
биохимическое взаимодействие,
Благодаря этому происходит синтез
органических веществ с использованием
молекулярного азота, недоступного
в такой форме для высших зеленых рутений.
Часть веществ, синтезированных в
клубеньках, усваивают бобовые растения,
бактерии же используют различные
вещества, находящиеся в корнях. Таким
образом, бобовые растения и клубеньковые
бактерии находятся в состоянии симбиоза.
Между клетками
корня и бактериями существует тесное
биохимическое взаимодействие,
Благодаря этому происходит синтез
органических веществ с использованием
молекулярного азота, недоступного
в такой форме для высших зеленых рутений.
Часть веществ, синтезированных в
клубеньках, усваивают бобовые растения,
бактерии же используют различные
вещества, находящиеся в корнях. Таким
образом, бобовые растения и клубеньковые
бактерии находятся в состоянии симбиоза.
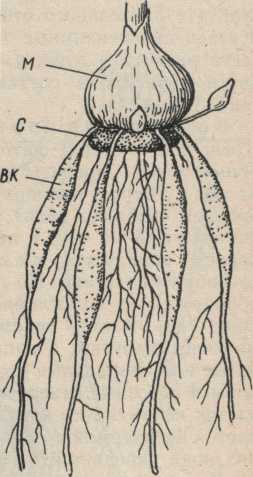
Рис.6.
Клубнелуковица
гладиолуса с
утолщенными
у основания
втягивающими
корнями:
ВК — втягивающие
корни; М - молодая клубнелуковица;
С — старая клубнелуковица.
Кроме бобовых, ольха, лох, подокарпус и
некоторые другие растения способны образовывать на корнях клубеньки или сходные образования, содержащие азотфиксирующие микроорганизмы, однако их экономическое значение ничтожно по сравнению с бобовыми растениями.
Втягивающие корни могут укорачиваться у своего основания. Так как они прочно срастаются с почвой, то укорочение приводит к втягиванию побега (луковицы, корневища) в почву. Следовательно, втягивающие корни помогают побегам находить наилучшую глубину залегания в почве.
Втягивающие корни узнаются по утолщенным основаниям с поперечной морщинистостью (рис. 6).
Запасающие корни обычно утолщены и сильно паренхиматизированы. Запасающая паренхима находится в первичной коре, в древесине или в сердцевине.
Сильно утолщенные придаточные корни георгина, чистяка, любки называют корневыми шишками.
У многих, чаще всего двулетних, растений возникает образование, носящее название корнеплода. Морфологическая природа его сложна, так как в его образовании принимают участие и корень, и стебель). У моркови почти весь корнеплод, за исключением самой верхней части, составлен корнем. У репы корень образует лишь самую нижнюю часть корнеплода. Можно найти много случаев промежуточного характера. Даже разные сорта (например, у редиса) различаются по степени участия корня и стебля в образовании корнеплода. Границы этих органов устанавливаются при прослеживании развития растения от проростка до взрослого состояния.
Воздушные корни образуются у многих тропических эпифитов (греч. эпи — сверх; фитон — растение) из семейств орхидных, ароидных, бромелиевых. Эпифиты не паразитируют на деревьях, а только используют их как субстрат, подпорку, для поднятия вверх, к свету. Воздушные корни орхидей свободно висят в воздухе и приспособлены к поглощению влаги, попадающей на них в виде дождя или росы.
Дыхательные корни хорошо развиты у некоторых тропических деревьев, обитающих по болотистым побережьям океанов. У авиценнии образуется очень сложная корневая система с дыхательными корнями, вырастающими вертикально вверх. На концах этих корней имеется система отверстий, связанных с аэренхимой. Воздух через эти ткани поступает в подводные корни

. Рис. 7. Столбовидные корни баньяна.
Ходульные корни образуются у деревьев, живущих в тех же мангровых зарослях по берегам океанов, что и авиценния. Благодаря своим сильно разветвленным корням деревья стоят словно на ходулях и сохраняют устойчивость на зыбком илистом грунте даже во время отлива, распределяя свою массу на большую площадь опоры («лыжный эффект»).
Столбовидные корни (корни-подпорки) особенно замечательны у индийских баньянов (рис. 7). Эти корни закладываются как придаточные на горизонтальных ветвях дерева и свешиваются вниз. Достигнув почвы, они сильно разрастаются, превращаясь в столбовидные образования, поддерживающие крону дерева. Такие корни-подпорки позволяют дереву разрастись в стороны, покрывая площадь до 2500 м2.
ОПОРНЫЕ ТОЧКИ
Корень- основной вегетативный орган растения, выполняющий функцию почвенного питания. Корень — осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы.
Ризодерма - ткань, взявшая на себя функцию поглощения веществ из почвы, сохраняющая на поверхности важнейшие в функциональном отношении жизнедеятельные клетки.
Ринофиты - это растения водоросли имели небольшие размеры, жили у воды и в воде.
Чехлик - покрывает нежные клетки апикальной меристемы, состоит из живых клеток и постоянно обновляется: по мере того как с его поверхности слущиваются старые.
Колонка ( колумелла)- центральная (осевая) часть чехлика.
Статолиты - (греч. статос — стоящий; литос — камень), т. е. способны перемещаться в клетке при изменении положения кончика корня в пространстве, благодаря чему корень изгибается и растет в прежнем направлении.
Меристема- ткань способная к постоянному делению.
Корневой волосок - поглощает из почвы растворы минеральных солей, а также своими выделениями растворяет некоторые химические соединения.
Микориза - корневые окончания и сросшиеся с ними гифы грибов образуют микоризу (дословно «грибокорень»).
Экзодерма - защищает ткани корня в зоне проведения.
ВОПРОСЫ ДЛЯ ПОВТОРЕНИЯ
1.Дать понятие корня.
2.Главная функция и иные функции корня.
3.Эволюция корня.
4.Появление специализированных тканей корня. Каких?
5.Благодаря чему увеличивается поглощающая поверхность корня?
6.Зоны молодого корневого окончания
А. зона деления и корневого чехлика
б. зона всасывания
в. зона проведения
7. Какие по морфологическим признакам корни бывают
8. Что лежит в основе типов корневых систем? Характеристика этих типов.
9. Метаморфозы корней. Какими они бывают?
10.Микориза и сожительство с бактериями. Какие плюсы и минусы из такого взаимного сожительства?