

28. Мембранный потенциал. Методы измерения мембранного потенциала. Микроэлектродная техника.
Метод фиксации потенциала. В данном методе используют электронную систему с обратной связью, которая обеспечивает автоматическое поддержание мембранного потенциала. Разность потенциалов по разные стороны мембраны фиксируют на определенном уровне, при этом мембранный потенциал можно ступенчато изменять на строго определенную величину. Такой метод позволяет измерить ионные токи, протекающие сквозь мембрану через каналы, которые активируются при изменении потенциала.
Метод фиксации напряжения позволяет измерять ионный поток при перемещении иона по контролируемому градиенту электрохимического потенциала и получать информацию об электрической проводимости мембраны и её пассивной проницаемости в отношении интересующего нас иона.
Метод пэтч-кламп применяется стеклянная микропипетка со сглаженными краями диаметром около 1 мкм вместо «острых» микроэлектродов. Пипетка наполняется раствором, в зависимости от конкретного исследования или метода пэтч-кламп. Контактирующий с этим раствором металлический электрод передает изменения заряда на усилитель мембранного потенциала Он прижимается к клеточной мембране, после чего мембрана всасывается внутрь пипетки с электродом, образуя электронепроницаемую «гигаомическую» изоляцию. При данном методе один-единственный электрод используется для фиксации потенциала клеточной мембраны, за счет чего напряжение сохраняется на постоянном уровне, в то время как регистрируются изменения тока. Аналогично, может быть использован токопроводящий зажим для записи изменений напряжения.
Регистрация от целой клетки в условиях плотного контакта. Для регистрации на целой клетке пэтч-пипетки заполняют соответствующим раствором с низкой концентрацией ионов кальция. Пипетку прижимают к поверхности клеточной мембраны, в результате чего формируется высокоомный контакт. Затем потенциал пипетки изменяют до отрицательного значения и подают импульсы напряжения с амплитудой в несколько милливольт. На этом этапе производится компенсация быстрой ёмкостной компоненты, обусловленной ёмкостью держателя и стенок.
Гигантский аксон кальмара, диаметром 0,5мм. Можно ввести микроэлектрод не нанося повреждений. Микроэлектрод стеклянный – микропипетка с тонким кончиком. Металлический электрод не может проколоть мембрану. Для исключения поляризации электрода используют неполяризационные электроды – серебряная проволока покрытая AgCl, в растворе KCl или NaCl (заполняют электрод). Второй электрод (сравнение) расположен у наружной поверхности клетки. Регистрирующее устройство (с усилителем постоянного тока) измеряет мембранный потенциал.
29. Возникновение потенциала покоя. Гипотеза Бернштейна. Уравнение Нернста. Уравнение Гольдмана – Ходжами – Катца. Эквивалентная электрическая схема мембраны.
Для описания изменений мембранного потенциала при возбуждении часто используют и другой подход, основанный на представлении ионных проводимостей мембраны в виде эквивалентной электрической цепи
1/gK
1/gCl
1/gNa
CM K
Cl
Na

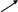




 о
о
i
Мембранный потенциал, измеряемый на схеме между точками i и о, создается электродвижущими силами, равными равновесными потенциалами отдельных видов ионов, последовательно соединенных с соответствующими сопротивлениями. Ток в каждой из ветвей равен:
Ii=gi(i), (3)
Где gi – проводимость мембраны для данного вида ионов, i – равновесный потенциал данного вида ионов, а - общий потенциал на мембране. Используя условие, что в разомкнутой цепи при постоянном напряжении на мембране сумма токов равна нулю, можно получить следующее выражение для мембранного потенциала:
![]() , (4)
, (4)
где gM – суммарная проводимость мембраны: gM = gK + gNa + gCl.
Из (4) видно, что значение мембранного потенциала определяется теми ионами, для которых проводимость максимальна.
Эквивалентная схема ионных проводимостей мембраны и основанное на ней уравнение (4) предполагают, что разные виды ионов проникают через клеточную мембрану по пространственно разделённым путям.