

Вопрос 35. Рецептивные поля.
Рецептивным полем называется область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 17.1). Максимальная величина рецептивного поля первичного сенсорного нейрона определяется пространством, которое занимают все ветви его периферического отростка, а число рецепторов, имеющихся в этом пространстве, указывает на плотность иннервации. Высокая плотность иннервации сочетается, как правило, с малыми размерами рецептивных полей и, соответственно, высоким пространственным разрешением, позволяющим различать стимулы, действующие на соседние рецептивные поля. Малые рецептивные поля типичны, например, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов значительно выше, чем на периферии сетчатки или в коже спины, для которых характерны большая величина рецептивных полей и меньшее пространственное разрешение. Рецептивные поля соседних сенсорных нейронов могут частично перекрывать друг друга, поэтому информация о действующих на них стимулах передается не по одному, а по нескольким параллельным аксонам, что повышает надежность ее передачи.
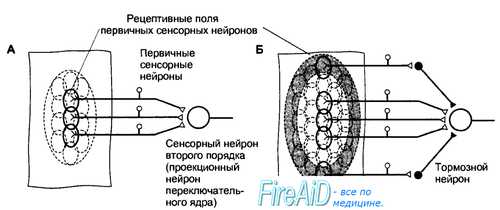
Рис. 17.1. Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Б. Раздражение центральной или периферической области рецептивного поля сенсорного нейрона второго и следующих порядков сопровождается противоположным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение). В результате контраста, создаваемого между центром и периферией рецептивного поля, выделяется информация для передачи на следующий иерархический уровень.
Величина рецептивных полей сенсорных нейронов второго и следующих порядков больше, чем у первичных сенсорных нейронов, поскольку центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предшествующего уровня. От центра рецептивного поля информация передается непосредственно к сенсорным нейронам следующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, поэтому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В результате сигналы от центра рецептивного поля беспрепятственно достигают следующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая функциональная организация рецептивных полей обеспечивает выделение наиболее значимых сигналов, легко различаемых на контрастном с ними фоне.
Рецептивное поле (англ. receptive field) сенсорного нейрона — участок с рецепторами, которые при воздействии на них определённого стимула приводят к изменению возбуждения этого нейрона.
Концепция рецептивных полей может быть применима ко всей нервной системе. Если множество сенсорных рецепторов образуют синапсы c единственным нейроном, они совместно формируют рецептивное поле этого нейрона. Например, рецептивное поле ганглионарной (ганглиозной) клетки сетчатки глаза представлено фоторецепторными клетками (палочками или колбочками), а группа ганглионарных клеток в свою очередь создаёт рецептивное поле для одного из нейронов мозга. В итоге к одному нейрону более высокого синаптического уровня сходятся импульсы от многих фоторецепторов; и этот процесс называется конвергенцией.
Периферия рецептивного поля - это область тела, стимуляция которой сопровождается ответом сенсорных нейронов.
Рецепторная клетка воспринимает энергию внешнего раздражения и перерабатывает ее в нервный импульс. При этом каждая рецепторная клетка воспринимает раздражения с определенной зоны - рецептивного поля, представляющего собой все точки периферического отдела анализатора, возбуждение которых влияет на данный нейрон. Рецептивным полем называется совокупность точек на периферии, с которых периферические стимулы влияют на данную нервную клетку .
Рецептивные поля сенсорных нейронов широко варьируют по размерам. Одни нейроны имеют очень маленькие рецептивные поля - например, в зрительной коре для некоторых нейронов участок сетчатки , в пределах которого световой стимул оказывает на них действие, составляет всего 0,02 кв.мм. В то же время другие клетки в центральной нервной системе активируются кожными стимулами, действующими на область, занимающую всю ногу , причем эфффективны и прикосновение, и вибрация, и холодовые стимулы. Рецептивные поля смежных нейронов в сенсорном центре сильно перекрываются, что позволяет с большей точностью определять положение стимула.
Как правило, центр и периферия рецептивного поля связаны с противоположными ответами - например, от стимула в центре частота разрядов повышается во время стимуляции, а на периферии действие этого стимула будет обратное. Такая организация рецептивных полей основана на латеральном торможении . Разделение рецептивного поля на центр и периферию с противоположными свойствами обостряет способность к пространственному различению в мозговых центрах и усиливает контраст, т.е. ощущаемое различие между двумя интенсивностями стимулов. Размеры и организация рецептивных полей не являются неизменным свойством сенсорного нейрона. Величина рецептивного поля может быть уменьшена управляемыми центральными тормозными процессами, и даже относительные размеры центра и периферии могут изменяться.
Понятие рецептивного поля включает в себя сложную организацию на разных уровнях: синаптические связи сенсорной клетки с популяцией рецепторов , специализацию в отношении определенных качеств и возможность контролируемых из центров функциональных изменений в синаптических связях. Эти высокоспециализированные связи, как было показано в опытах на новорожденных котятах, существуют уже при рождении, однако могут быть утрачены, если не используются. Таким образом, хотя очень специфические синаптические связи являеются врожденными, они могут быть изменены практикой или фактором научения , в особенно восприимчивые периоды развития животного.
Во многих случаях, рецептивные поля имеют свою внутреннюю организацию. Часто стимуляция одной из частей РП ведет к возбуждению сенсорного нейрона, в другой - к торможению. Рецептивные поля существенно разнятся и по площади. В тех участках сенсорной поверхности, где важно, чтобы стимул был точно локализован, РП малы. Напротив, там, где локализация стимула не так важна, РП больше. Это хорошо показано опытом Фехнера по изучению порога в двух точках. Некоторые части тела - кончики пальцев и губы - способны различать два близко расположенных стимула (укола), тогда как другие (тыльная сторона ладони, спина) - только значительно дальше разнесенные стимулы. В некоторых случаях есть и связь между чувствительностью и точностью локализации ощущения.