

Стр. 62 № 12 (б)
МУТАЦИИ – это наследуемые изменения генетического материала
Классификация мутаций:
Существует несколько принципов такой классификации.
-
По характеру изменения генома:
-
Геномные мутации – изменение числа хромосом.
-
Хромосомные мутации, или хромосомные перестройки, - изменение структуры хромосом.
-
генные мутации – изменение генов
-
По проявлению в гетерозиготе:
-
Доминантные мутации
-
Рецессивные мутации.
-
По уклонению от нормы
-
Прямые мутации.
-
Реверсии. Иногда говорят об обратных мутациях, однако очевидно, что они представляют собой только часть реверсий, поскольку в действительности широко распространены т.н. супрессорные мутации.
-
В зависимости от причин, вызывающих мутации:
-
Спонтанные, возникающие без видимой причины, т.е. без каких-либо индуцирующих воздействий со стороны экспериментатора.
-
Индуцированные мутации.
Только эти четыре способа классификации изменений генетического материала носят достаточно строгий характер и имеют универсальное значение. Каждый из подходов в этих способах классификации отражает некоторую существенную сторону возникновения либо проявления мутаций у любых организмов: эукариот, прокариот и их вирусов.
Существуют и более частные подходы к классификации мутаций:
-
По локализации в клетке:
-
Ядерные.
-
Цитоплазматические. В этом случае обычно подразумевают мутации неядерных генов.
-
По отношению к возможности наследования:
-
Генеративные, происходящие в половых клетках
-
Соматические, происходящие в соматических клетках.
Очевидно два последних способа классификации мутаций применимы только к эукариотам, а рассмотрение мутаций с точки зрения их возникновения в соматических или половых клетках имеет отношение только к многоклеточных эукариотам.
-
По фенотипическому проявлению:
1. Летальные
2. Морфологические
3. Биохимические
4. Поведенческие
Межхромосомные мутации , илимутации перестройки - обмен фрагментами между негомологичными хромосомами. Такие мутации получили название транслокации (от лат. trans - за, через + locus - место)

Схематическое изображение транслокации хромосом.
А – нормальные хромосомы, Б – транслоцированные хромосомы.
Это:
- реципрокная транслокация, когда две хромосомы обмениваются своими фрагментами;
- нереципрокная транслокация, когда фрагмент одной хромосомы транспортируется на другую;
- "центрическое" слияние (робертсоновская транслокация) – соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч.
При поперечном разрыве хроматид через центромеры "сестринские" хроматиды становятся "зеркальными" плечами двух разных хромосом, содержащих одинаковые наборы генов. Такие хромосомы называют изохромосомами.
Как внутрихромосомные (делеции, инверсии и дупликации), так и межхромосомные (транслокации) аберрации и изохромосомы связаны с физическими изменениями структуры хромосом, в том числе с механическими разломами.
Стр. 55 № 5
Репликация ДНК (DNA) (воспроизведение генотипа) происходит по полуконсервативному механизму. Каждая нить двойной спирали выступает в роли матрицы для синтеза новой цепи. Следовательно, вновь образованные двухспиральные молекулы состоят из одной «новой» и одной «старой» цепи.

Полуконсервативный механизм репликации ДНК
Субстратами синтеза являются дезоксинуклеозидтрифосфаты, выполняющие роль строительного материала и источников энергии. Для репликации ДНК необходим большой набор разнообразных ферментов и белков - репликативный комплекс. Далее будет описана функция основных компонентов этого комплекса. Белки, раскручивающие спираль ДНК, и белки, стабилизирующие разделенные нити ДНК. В результате действия этих белков образуется репликативная вилка - участок ДНК, в пределах которого спираль раскручена и разделена на отдельные цепи. ДНК -полимераза обеспечивает включение в растущую «новую» цепь нуклеотидов комплементарных «старой», то есть матричной цепи. ДНК -полимераза не способна начинать синтез новой цепи с ее первого нуклеотида. Она может удлинять уже имеющуюся цепь, поэтому для начала реакции требуется затравка (праймер), которая представляет собой короткий полинуклеотид, комплементарный матричной цепи ДНК. Фермент присоединяется к матричной цепи ДНК и к праймеру в области3’-концевого нуклеотида праймера. Перемещаясь по матрице в направлении ее 5’-конца, ДНК -полимераза удлиняет затравку, присоединяя к ней один за другим нуклеотиды. Одноцепочечная затравка - праймер синтезируется при участии ДНК - зависимой РНК -полимеразы (праймазы). Синтез новых цепей ДНК может протекать только в направлении 5’ à 3’. Таким образом, на одной цепи ДНК синтезируется непрерывно «лидирующая» цепь, а на другой образуются короткие фрагменты- «запаздывающая» цепь. Затем последовательность праймера удаляется и образовавшийся промежуток заполняется с помощью ДНК -полимеразы.
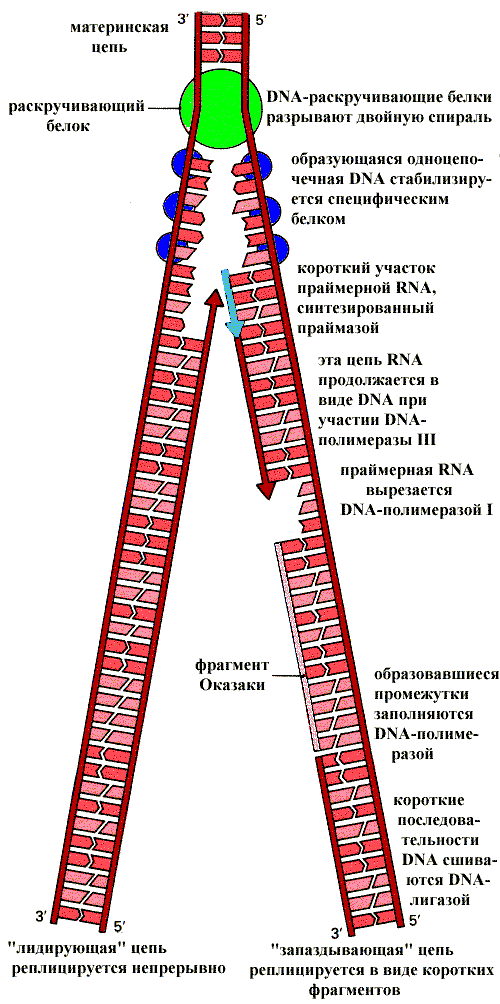
Репликация ДНК
ДНК -лигаза способна сшивать полученные короткие фрагменты, после чего формируется новая двухспиральная молекула DNA.
Стр. 38 № 11
Кроссинговер (от англ. crossing–over – перекрёст) – это обмен гомологичными участками гомологичных хромосом (хроматид).
Само явление перекрёста хромосом открыл Ф. Янссенс (1909, Бельгия), обнаруживший хиазмы в профазе I мейоза у саламандры. Однако теоретическая возможность кроссинговера была предсказана раньше. Во-первых, американский цитолог У. Сэттон (1903) предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться сцепленное наследование признаков, т.е. несколько разных признаков могут наследоваться так, как будто они контролируются одним геном. Тогда совокупность генов в одной хромосоме образует группу сцепления. Во-вторых, в 1906 г. У. Бэтсон и Р. Пеннет обнаружили сцепленное наследование у душистого горошка. Они изучали совместное наследование: окраски цветков (пурпурная или красная) и формы пыльцевых зерен (удлиненная или округлая). При скрещивании дигетерозигот в их потомстве наблюдалось расщепление 11,1:0,9:0,9:3,1 вместо ожидаемого 9:3:3:1. Таким образом, два признака (окраска цветков и морфология пыльцы) оказываются сцепленными. Соответственно, сцеплены и гены, контролирующие эти признаки («эффект взаимного притяжения»). Однако это сцепление не абсолютно: гены, контролирующие сцепленные признаки, могут приобретать независимость, и тогда появляются новые комбинации признаков. Вскоре (1911) были получены данные по сцепленному наследованию и нарушению сцепления некоторых признаков у дрозофилы.
Т. Морган (1910) и его ученик К. Бриджес (1916) доказали, что число групп сцепления соответствует гаплоидному числу хромосом. Опираясь на работы Янссенса и результаты гибридологического анализа, они связали явление перекрёста хромосом с феноменом нарушения сцепления генов. При перекрёсте хромосом происходит обмен между ними генетическим материалом (аллелями), и тогда происходит рекомбинация – появление новых сочетаний аллелей, например, AB + ab → Ab + aB. Таким образом, термин «кроссинговер» употребляется и в цитологическом, и в генетическом значении. В дальнейшем связь между перекрестом хромосом и появлением новых сочетаний признаков была подтверждена работами К. Дарлингтона, К. Штерна, Б. Мак-Клинток и др.
Цитогенетика кроссинговера