

12.4. Биосинтез нуклеиновых кислот
Первичную структуру важнейших биополимеров - белков и нуклеиновых кислот можно сравнить с буквенной записью: и в том, и в другом случае имеется не произвольное, а строго определенное, имеющее смысл, чередование элементов - мономеров или «букв». На этом основании нуклеиновые кислоты и белки называют информационными молекулами.
До сих пор не раскрыты в деталях молекулярные механизмы передачи генетической информации, записанной (закодированной) в нуклеотидной последовательности ДНК.
Но однозначно, что при биосинтезе новых молекул нуклеиновых кислот и белков носителями информационной программы являются нуклеиновые кислоты; в этой роли их называют матрицами. Матрица в ходе матричного синтеза не расходуется и может использоваться многократно; в этом отношении она сходна с катализатором.
Различают три основных этапа реализации генетической информации (три основных типа матричных биосинтезов).
1-й этап - биосинтез ДНК (репликация) с использованием в качестве матрицы уже существующих молекул ДНК. Репликация ДНК является ключевой функцией делящейся клетки.
2-й этап - транскрипция - генетическая информация, записанная в первичной структуре ДНК, переписывается в нуклеотидную последовательность РНК (т. е. биосинтез РНК на матрице ДНК).
3-й этап - трансляция - генетическая информация, содержащаяся в нуклеотидной последовательности молекулы мРНК, переводится в аминокислотную, последовательность белка (биосинтез белков на матрице мРНК).
В данном разделе рассматриваются первые два этапа, третий этап будет рассмотрен в главе 13 «Обмен белков».
Биосинтез днк (репликация)
Для биосинтеза ДНК прежде всего необходимо наличие всех четырех видов дезоксирибонуклеозидтрифосфатов (дАТФ, дГТФ, дЦТФ и дТТФ). Кроме того, известно, что для любого синтеза полимерной органической молекулы, осуществляемого в живой или неживой природе, требуется большое количество энергий. Источником энергии в реакциях полимеризации мононуклеотидов служит энергия, освобождаемая всеми четырьмя типами дезоксирибонуклеозидтрифосфатов, участвующих в синтезе ДНК. Образующийся пирофосфат (Н4Р2О7) под действием пирофосфатазы также расщепляется на две молекулы ортофосфата (Н3РО4), давая дополнительную энергию для биосинтеза ДНК, который требует кроме того наличия специфических ферментов.
Ферментная система ДНК до конца не расшифрована, она включает более 20 ферментов и белковых факторов, объединенных в единую ДНК - репликационную систему.
Общая схема биосинтеза ДНК может быть представлена в следующем виде:
m(дАТФ
+ дТГФ) + n(дГТФ
+дЦТФ)
 ДНК +(m+n)
+ Н4Р2О7
ДНК +(m+n)
+ Н4Р2О7
Химический смысл полимеризации состоит в том, что 3'-гидроксильная группа матрицы атакует α - атом фосфора соответствующего нуклеозидтрифосфата (определяемого матрицей по комплементарному механизму); при этом происходит отщепление остатка пирофосфата. Далее 3'-гидроксил вновь образованного мононуклеотида атакует α-атом фосфора следующего нуклеозидтрифосфата, и таким путем продолжается процесс полимеризации, идущий в направлении 5'→3', в противоположность матрице, оканчивающейся 5'-фосфатом. Реакция требует присутствия в качестве матрицы одноцепочной ДНК или в крайнем случае небольших полинуклеотидов.
В настоящее время механизм репликации ДНК представляется следующим образом. Первоначально с помощью ферментов происходит раскручивание двойной спирали ДНК, в результате чего образуется репликативная вилка - цепи ДНК расходятся в стороны. Затем при участии ДНК-полимеразы (репликазы) образуются новые полинуклеотидные цепи, комплементарные исходным матричным; в результате получаются две двухцепочечные молекулы ДНК, полностью идентичные исходной молекуле:
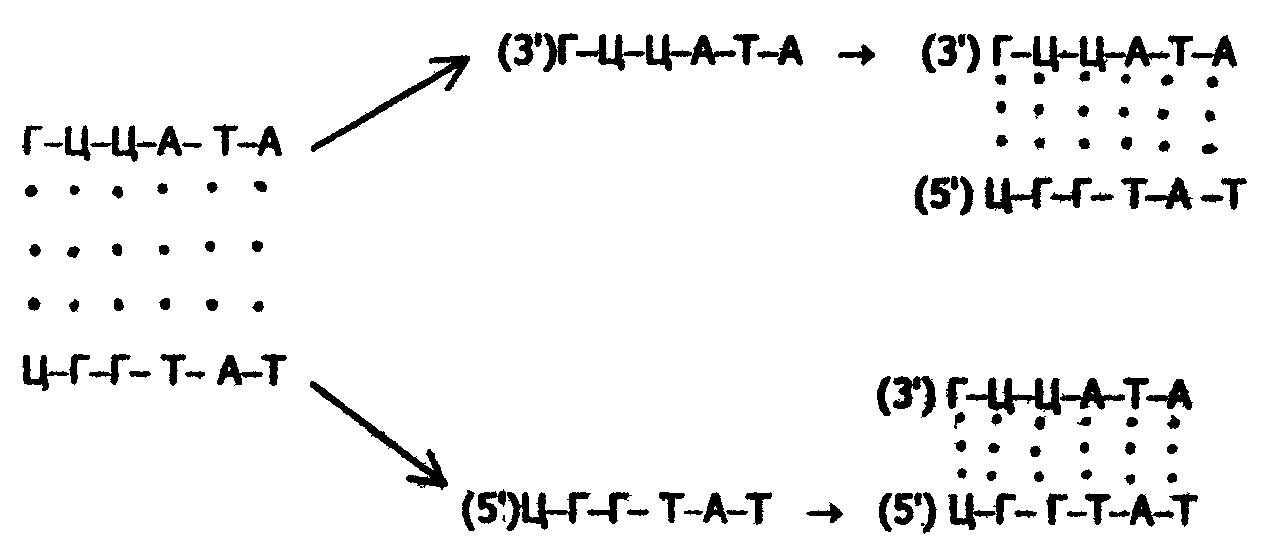
Такой способ репликации: получил название полуконсервативного (в принципе возможен и другой механизм - консервативный, при котором вновь синтезируемая нуклеотидная цель образуется прямо на двойной спирали ДНК, без ее раскручивания).
Сложность процесса репликации ДНК объясняется тем, что обе цепи реплицируются одновременно, хотя имеют разное направление (5'→3' и 3´→5'), кроме того рост (элонгация) дочерних цепей также должен происходить в противоположных направлениях. С другой стороны, как уже выше было сказано, элонгация каждой дочерней цепи может идти только в направлении 5'→ 3'. Поэтому ученым Р. Оказаки было высказано предположение, что синтез одной из дочерних цепей происходит непрерывно в одном направлении, а в это время синтез второй дочерней цепи идет прерывисто путем соединения коротких фрагментов (фрагментов Оказаки), синтезируемых в противоположном направлении (рис. 36). По мере роста новых цепей репликативная вилка перемещается по ДНК.

Рис 36. Синтез ДНК: а - на одной ветви репликативной вилки синтезируется непрерывная нуклеотидная цепь, на другой - фрагменты Оказаки; 6 - фрагменты Оказаки соединяются друг с другом в результате действия ДНК-лигазы