

Ассоциативное звено анс
В зависимости от расположения ассоциативного звена (уровня замыкания), различают следующие рефлексы:24
-
Местные25
-
ганглионарные, 26
-
спинальные,
-
бульбарные
Доказано существование периферических рефлекторных дуг в желудочно-кишечном тракте, сердечной мышце и т.п.27
Эфферентное звено анс
Эффекторных нейронов несравненно больше, чем преганглионарных волокон. Например, в верхнем шейном симпатическом ганглии одно преганглионарное симпатическое волокно контактирует более чем с сотней эффекторных нейронов.
При этом на одном и том же эффекторном нейроне могут оканчиваться разветвления нескольких преганглионарных волокон. Наличие таких широких конвергентных и дивергентных отношений обеспечивает надежность проведения возбуждения.
Эта закономерность касается только симпатической части автономной нервной системы, в двух других ее частях подобная конвергенция практически отсутствует.
В интеграции сигналов в низших центрах вегетативной периферии значительная роль отводится пространственной и временной суммации постсинаптических потенциалов. Роль их состоит в том, что поступающие по пресинаптическим терминалям присущие им относительно слабые сигналы благодаря этим процессам трансформируются, превращаясь в сверхпороговые постсинаптические потенциалы эфферентного нейрона.
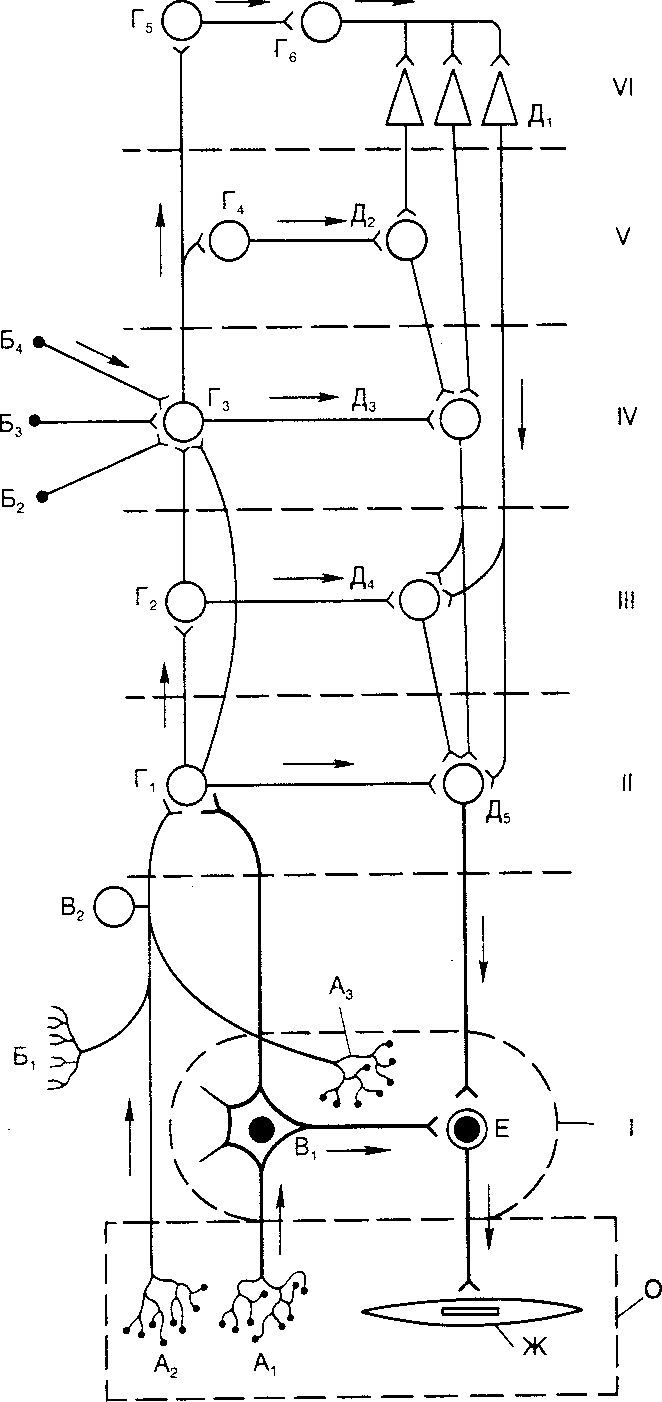
Рис. 4.24. Основные связи автономной нервной системы (по С. П. Семенову).
O — висцеральный орган; I — автономный (около-, предпозвоночный или интрамуральный) узел; II — сегмент спинного мозга или ствола мозга; III — надсегментарные центры продолговатого мозга; IV — промежуточный мозг; V — ядра основания мозга; VI — кора больших полушарий головного мозга; A1 и А2 — рецепторы внутренних органов; А3 — рецептор автономного нервного узла; Б1 — экстероцептор; Б2 — Б4 — проводящие пути из органов чувств. В1 — чувствительный нейрон автономного нервного узла; В2 — чувствительные нейроны спинномозговых и черепномозговых узлов; Г1 — Г4 — ассоциативные ядра разных отделов мозга; Г5 — клетки афферентных зон коры большого мозга; Г6 — ассоциативные клетки; Д1 — пирамидные клетки коры большого мозга; Д2 — ядра основания; Д3 — ядра подбугорья; Д4 — клетки ретикулярной формации продолговатого мозга; Д5 — ядра автономной нервной системы в стволе мозга и сегментах спинного мозга; Е — эффекторный нейрон дуги автономного рефлекса; Ж — эффекторные клетки стенки висцерального органа. Стрелки обозначают распространение возбуждения.
Тело эффекторной клетки дуги автономного рефлекса представляет собой мигрировавшую из спинного мозга клетку, располагающуюся в одном из периферических автономных ганглиев (рис. 4.24). Нейроны этих ганглиев охватывают своим влиянием, как правило, большие территории висцеральных органов. Ганглии могут располагаться либо около позвоночника (превертебральные), либо в сплетениях вблизи внутренних органов (паравертебралъные), наконец, в тканях внутренних органов (интрамуральные, интервисцеральные).
Эффекторный нейрон дуги автономного рефлекса по электрическим показателям в покоящемся состоянии мало чем отличается от мотонейрона соматической дуги. Однако кратковременная или одиночная стимуляция преганглионарных волокон вызывает появление в нем сложной последовательности медленных деполяризующих и гиперполяризующих постсинаптических процессов. В этом случае вначале возникает локальный отрицательный потенциал (О‑волна), переходящий в положительную П‑волну. Последняя сменяется поздней отрицательной П‑волной. Каждая из этих фаз отражает межнейронную передачу, при этом О‑волна — возникающий в холинергических синапсах возбуждающий постсинаптический потенциал (ВПСП). Появление П‑волны обусловлено возбуждением особой группы преганглионарных волокон, которые оканчиваются на хромаффинных клетках ганглия. Появление деполяризующихся и гиперполяризующихся постсинаптических потенциалов опосредуется мускариноподобным действием ацетилхолина, в то время как гиперполяризация — специальными вставочными адренергическими клетками, регулируя тем самым возбудимость эффекторных нейронов.
Как правило, эффекторный нейрон может иметь, помимо основного возбуждающего холинергического преганглионарного входа, еще и прямой вход сугубо периферического происхождения, представляющий одно из звеньев местной рефлекторной дуги ганглионарного уровня. Электрофизиологические характеристики эффекторного нейрона позволяют интегрировать эти сигналы и формировать новый выходной сигнал. Благодаря этим местным дугам в эфферентном нейроне поддерживается необходимый уровень спонтанной активности и при децентрализации ганглия сохраняется его рефлекторная функция. У спонтанно активных эфферентных нейронов фоновые разряды характеризуются низкой частотой. Они могут возникать синхронно пульсовым толчкам, дыхательным и перистальтическим движениям. Паттерн и ритм разрядов совпадают с показателями преганглионарных волокон или активностью волокон местных рефлекторных дуг.