

Биология как одна из теоретических основ медицины, ее задачи, объект и методы исследования. Разделы дисциплины биологии и их значение для деятельности врача.
Биология — наука о жизни. Она изучает жизнь как особую форму движения материи, законы ее существования и развития. Предметом изучения биологии являются живые организмы, их строение, функции, их природные сообщества. Термин «биология», предложенный в 1802 г. впервые Ж. Б. Ламарком. Вместе с астрономией, физикой, химией, геологией и другими науками, изучающими природу, биология относится к числу естественных наук. В общей системе знаний об окружающей мире другую группу наук составляют социальные, или гуманитарные науки, изучающие закономерности развития человеческого общества.
Современная биология представляет собой систему наук о живой природе. Биологические науки служат теоретической основой медицины, агрономии, животноводства, а также всех тех отраслей производства, которые связаны с живыми организмами.
Методы биологических наук. Основными частными методами в биологии являются: описательный, сравнительный, исторический и экспериментальный.
Для того чтобы выяснить сущность явлений, необходимо прежде всего собрать фактический материал и описать его. Собирание и описание фактов были главным приемом исследования в ранний период развития биологии, который, однако, не утратил значения и в настоящее время.
Еще в XVIII в. получил распространение сравнительный метод, позволяющий путем сопоставления изучать сходство и различие организмов и их частей. На принципах этого метода была основана систематика, сделано одно из крупнейших обобщений — создана клеточная теория. Применение сравнительного метода в анатомии, палеонтологии, эмбриологии, зоогеографии способствовало утверждению эволюционных представлений. Сравнительный метод перерос в исторический, но не потерял значения и сейчас.
Исторический метод выясняет закономерности появления и развития организмов, становления их структуры и функции. Утверждением в биологии исторического метода наука обязана Дарвину.
Экспериментальный метод исследования явлений природы связан с активным воздействием на них путем постановки опытов (экспериментов) в точно учитываемых условиях и путем изменения течения процессов в нужном исследователю направлении. Этот метод позволяет изучать явления изолированно и добиваться повторяемости их при воспроизведении идентичных условий. Эксперимент обеспечивает не только более глубокое, чем другие методы, проникновение в сущность явлений, но и непосредственное овладение ими. Высшей формой эксперимента является моделирование изучаемых процессов.
Место и задачи биологии в системе подготовки врача. Важность изучения биологии для медика определяется тем, что биология — это прежде всего основа медицины. «Медицина, взятая в плане теории,— это прежде всего общая биология»,— писал один из крупнейших теоретиков медицины И. В. Давыдовский (1887—1968). Успехи медицины связаны с биологическими исследованиями, поэтому врач постоянно должен быть осведомлен о новейших достижениях биологии. Достаточно привести несколько примеров из истории науки, чтобы убедиться з тесной связи успехов медицины с открытиями, казалось бы, в чисто теоретических областях биологии. Исследования Л. Пастера (1822—1895), опубликованные в 1862 г. и доказавшие невозможность самопроизвольного зарождения жизни в современных условиях, открытие микробного происхождения процессов гниения и брожения произвело переворот в медицине и обеспечило развитие хирургии. В практику были введены сначала антисептика (предохранение заражения раны посредством химических веществ), а затем асептика (предупреждение загрязнения путем стерилизации предметов, соприкасающихся с раной). Это же открытие послужило стимулом к поискам возбудителей заразных болезней, а с обнаружением их связаны разработка профилактики и рационального лечения.
Изучение физиологических и биохимических закономерностей, открытие клетки и изучение микроскопического строения организмов позволило глубже понять причины возникновения болезненного процесса, способствовали внедрению в практику новых методов диагностики и лечения. Новейшие исследования в области закономерностей деления клеток и клеточной дифференцировки имеют прямое отношение как к проблеме регенерации, т. е. восстановлению поврежденных органов, так и к проблеме злокачественного роста, борьбе с онкологическими заболеваниями. Изучение И. И. Мечниковым (1845— 1916) процессов пищеварения у низших из многоклеточных организмов привело к открытию фагоцитоза и способствовало объяснению явлений иммунитета, сопротивляемости организма возбудителям болезни. И современные представления об иммунитете опираются на биологические исследования. Раскрытие механизмов иммунитета необходимо также для преодоления тканевой несовместимости, проблемы очень важной для восстановительной хирургии, с которой связаны вопросы трансплантации органов.
Исследования И. И. Мечникова по межвидовой борьбе у микроорганизмов явились предпосылкой открытия антибиотиков, используемых для лечения многих болезней, а массовое производственное получение антибиотиков стало возможно лишь благодаря применению методов генетики для создания высокопродуктивных штаммов продуцентов антибиотиков.
Советский исследователь Б. П. Токин открыл у растений летучие вещества — фитонциды, нашедшие широкое применение в медицине.
Следует помнить, что структуры и функции человеческого организма, в том числе защитные механизмы,—результат длительных эволюционных преобразований предшествующих форм. Поэтому в основе патологических процессов также лежат общебиологические закономерности. Необходимой предпосылкой для понимания сущности патологического процесса является знание биологии. Филогенетический принцип, учитывающий эволюцию органического мира, может подсказать правильный подход к изучению патологического процесса, а также для испытания новых лекарственных препаратов. Этот же метод помогает понять происхождение аномалий и уродств, найти наиболее рациональные пути реконструкции органов и т.
Большое число болезней имеет наследственную природу. Профилактика и лечение их требуют знаний генетики. Но и ненаследственные болезни протекают неодинаково и требуют различного лечения в зависимости от генетической конституции человека, чего не может не учитывать врач. Многие врожденные аномалии возникают вследствие воздействия неблагоприятных условий среды. Предупредить их — задача врача, вооруженного знаниями биологии развития организмов.
Здоровье людей в большой мере зависит от состояния окружающей среды. Знание биологических закономерностей необходимо для научно обоснованного отношения к природе, охране и использованию ее ресурсов, в том числе и с целью лечения и профилактики заболеваний.
Развитие представлений о сущности жизни. Определение жизни. Гипотезы о происхождении жизни. Главные этапы возникновения и развития жизни. Иерархические уровни организации жизни.
Уровня организации живого. В серединеХХ в. в биологии сложились представления об уровнях организации как конкретном выражении упорядоченности, являющейся одним из основных свойств живого (биологические микросистемы: мол., субклеточ., клеточ.; биолог.мезосист.:тк., ор., орг.; биол.макросис.: поп.-вид., биоценотич.).
Живое на нашей планете представлено в виде дискретных единиц — организмов, особей. Каждый организм, с одной стороны, состоит из единиц подчиненных ему уровней организации (органов, клеток, молекул), с другой — сам является единицей, входящей в состав надорганизменных биологических макросистем (популяций, биоценозов, биосферы в целом).
На всех уровнях жизни проявляются такие ее атрибуты, как дискретность и целостность, структурная организация (упорядоченность), обмен веществ, энергии и информации и т.д. Характер проявления основных свойств жизни на каждом из уровней имеет качественные особенности, упорядоченность. Как известно, в результате обмена веществ, энергии и информации устанавливается единство живого и среды, но понятие среды для разных уровней различно. Для дискретных единиц молекулярного и надмолекулярного (субклеточного) уровней окружающей средой является внутренняя среда клетки; для клеток, тканей и органов — внутренняя среда организма. Внешняя живая и неживая среда на этих уровнях организации воспринимается через изменение внутренней среды, т. е. опосредованно. Для организмов (индивидуумов) и их сообществ среду составляют организмы того же и других видов и условия неживой природы.
Существование жизни на всех уровнях подготавливается и определяется структурой низшего уровня. Характер клеточного уровня организации определяется молекулярным и субклеточным уровнями, организменный— клеточным, тканевым, органным, видовой (популяционный) — организменным и т. д. Следует отметить большое сходство дискретных единиц на низших уровнях и все возрастающее различие на высших уровнях.
Молекулярный уровень. На молекулярном уровне обнаруживается удивительное однообразие дискретных единиц. Жизненный субстрат для всех животных, растений, вирусов составляет всего 20 одних н тех же аминокислот и 4 одинаковых азотистых основания, входящих в состав молекул нуклеиновых кислот. Близкий состав имеют липиды и углеводы. У всех организмов биологическая энергия запасается в виде богатых энергией аденозинфосфорных кислот (АТФ, АДФ, АМФ). Наследственная информация у всех заложена в молекулах ДНК (исключение составляют лишь РНК-содержащие вирусы), способной к саморепродукции. Реализация наследственной информации осуществляется при участии молекул РНК, синтезируемых на матричных молекулах ДНК. В связи с тем, что с молекулярными структурами связано хранение, изменение и реализация наследственной информации, этот уровень иногда называют молекулярно-генетическим.
Клеточный уровень. На клеточном уровне также отмечается однотипность всех живых организмов. Клетка является основной самостоятельно функционирующей элементарной биологической единицей, характерной для всех живых организмов. У всех организмов только на клеточном уров-не возможны биосинтез и реализация наследственной информации. Клеточный уровень у одноклеточных организмов совпадает с организменным. В истории жизни на нашей планете был такой период (первая половина архейской эры), когда все организмы находились на этом уровне организации. Из таких организмов состояли все виды, биоценозы и биосфера в целом.
Тканевый уровень. Совокупность клеток с одинаковым типом организации составляет ткань. Тканевый уровень возник вместе с появлением многоклеточных животных и растений, имеющих дифференцированные ткани. У многоклеточных организмов он развивается в период онтогенеза. Большое сходство между всеми организмами сохраняется на тканевом уровне. Совместно функционирующие клетки, относящиеся к разным тканям, составляют органы. Всего лишь 5 основных тканей входят в состав органов всех многоклеточных животных и 6 основных тканей образуют органы растений.
Организменный (онтогенетический) уровень. На организменном уровне обнаруживается труднообозримое многообразие форм. Разнообразие организмов, относящихся к разным видам, да и в пределах одного вида,— следствие не разнообразия дискретных единиц низшего порядка, а все усложняющихся их пространственных комбинаций, обусловливающих новые качественные особенности. В настоящее время на Земле обитает более миллиона видов животных и около полумиллиона видов высших растений. Каждый вид состоит из отдельных индивидуумов.
Особь — организм как целое — элементарная единица жизни. Вне особей в природе жизнь не существует. На организменном уровне протекают процессы онтогенеза, поэтому уровень этот называют еще онтогенетическим. Нервная и гуморальная системы осуществляют саморегуляцию в организме и обусловливают определенный гомеостаз.
Популяционно-видовой уровень. Совокупность организмов (особей) одного вида, населяющих определенную территорию, свободно между собой скрещивающихся, составляет популяцию. Популяция — это элементарная единица эволюционного процесса; в ней начинаются процессы видообразования. Популяция входит в состав биогеоценозов.
Биоценотический и биосферный уровни. Биогеоценозы — исторически сложившиеся устойчивые сообщества популяций разных видов, связанных между собой и с окружающей неживой природой обменом веществ, энергии и информации. Они являются элементарными системами, в которых осуществляется вещественно-энергетический круговорот, обусловленный жизнедеятельностью организмов. Биогеоценозы составляют биосферу и обусловливают все процессы, протекающие в ней.
Только при комплексном изучении явлений жизни на всех уровнях можно получать целостное представление об особой (биологической) форме существования материи.
Представление об уровнях организации жизни имеет непосредственное отношение к основным принципам медицины. Оно заставляет смотреть на здоровый и больной человеческий организм как на целостную, но в то же время сложную иерархически соподчиненную систему организации. Знание структур и функций на каждом из них помогает вскрыть сущность болезненного процесса. Учет той человеческой популяции, к которой относится данный индивидуум, может потребоваться, например, при диагностике наследственной болезни. Для вскрытия особенностей течения заболевания и эпидемического процесса необходимо также учитывать особенности биоценотической и социальной среды. Имеет ли дело врач с отдельным больным или человеческим коллективом, он всегда основывается на комплексе знаний, полученных на всех уровнях биологических микро-, мезо- и макросистем.
Существуют две главные гипотезы, по-разному объясняющие появление жизни на Земле. Согласно гипотезе панспермии, жизнь занесена из космоса либо в виде спор микроорганизмов, либо путем намеренного заселения планеты разумными пришельцами из других миров. Прямых свидетельств в пользу космического происхождения жизни нет. Космос, однако, наряду с вулканами мог быть источником низкомолекулярных органических соединений, раствор которых послужил средой для развития жизни.
Современной наукой возраст Земли оценивается в 4,5—4,6 млрд. лет. Появление на планете первых водоемов, с которыми связывают зарождение жизни, отстоит от настоящего времени на 3,8—4 млрд. лет. Полагают, что около 3,8 млрд. лет назад жизнь могла стать определяющим фактором планетарного круговорота
углерода.
Согласно второй гипотезе, жизнь возникла на Земле, когда сложилась благоприятная совокупность физических и химических условий, сделавших возможным абиогенное образование органических веществ из неорганических.
В середине прошлого столетия Л. Пастер окончательно доказал невозможность самозарождения жизни в теперешних условиях. В 20-х годах текущего столетия биохимики А. И. Опарин и Дж. Холдейн предположили, что в условиях, имевших место на планете несколько миллиардов лет назад, образование
живого вещества было возможно. К таким условиям они относили наличие атмосферы восстановительного типа, воды, источников энергии (в виде ультрафиолетового (УФ) и космического излучения, теплоты остывающей земной коры, вулканической деятельности, атмосферных электрических явлений, радиоактивного распада), приемлемой температуры, а также отсутствие других живых существ.
Главные этапы на пути возникновения и развития жизни, по-видимому, состоят в: 1) образовании атмосферы из газов, которые могли бы служить сырьем для синтеза органических веществ (метана, оксида и диоксида углерода, аммиака, сероводорода, цианистых соединений), и паров воды; 2) абиогенном (т.е. происходящем без участия организмов) образовании простых органических веществ, в том числе мономеров биологических полимеров — аминокислот, Сахаров, азотистых оснований, АТФ и других мононуклеотидов; 3) полимеризации мономеров в биологические полимеры, прежде всего белки (полипептиды) и нуклеиновые кислоты (полинуклеотиды); 4) образовании предбиологических форм сложного химического состава — протобионтов, имеющих некоторые свойства живых существ; 5) возникновении простейших живых форм, имеющих всю совокупность главных свойств жизни,—примитивных клеток; 6) биологической эволюции возникших живых существ.
Существует много гипотез, пытающихся объяснить возникновение и развитие жизни на нашей планете. И хотя они предлагают различные подходы к решению данной проблемы, большинство из них предполагает наличие трех эволюционных этапов: химической, предбиологической и биологической эволюции.
На этапе химической эволюции происходил абиогенный синтез органических полимеров. На втором этапе формировались белково-нуклеиново-липоидные комплексы (ученые называли их по-разному: коацерваты, гиперциклы, пробион-ты, прогеноты и т. д.), способные к упорядоченному обмену веществ и самовоспроизведению. В результате предбиологи-ческого естественного отбора появились первые примитивные живые организмы, которые вступили в биологический естественный отбор и дали начало всему многообразию органической жизни на Земле.
Большинство ученых считают, что первыми примитивными живыми организмами были прокариоты. Они питались органическими веществами «первичного бульона» и получали энергию в процессе брожения, т. е. были анаэробными гетеротрофами.
С увеличением численности гетеротрофных прокари-отических клеток запас органических соединений в первичном океане истощался. В этих условиях значительное преимущество при отборе получали организмы, способные к автотрофности, т. е. к синтезу органических веществ из неорганических за счет реакций окисления и восстановления. Видимо, первыми автотрофными организмами были хемосинтезирующие бактерии. Следующим этапом было развитие фотосинтеза — комплекса реакций с использованием солнечного света. В результате фотосинтеза в земной атмосфере начал накапливаться кислород. Это явилось предпосылкой для возникновения в ходе эволюции аэробного дыхания. Способность синтезировать при дыхании большее количество АТФ позволила организмам расти и размножаться быстрее, а также усложнять свои структуры и обмен веществ.
Большинство ученых считает, что эукариоты произошли от прокариотических клеток. Существуют две наиболее признанные гипотезы происхождения эукариотических клеток и их органоидов.
Первая гипотеза связывает происхождение эукариотической клетки и ее органоидов с процессом впячивания клеточной мембраны.
Больше сторонников имеет гипотеза симбиотического происхождения эукариотической клетки. Согласно этой гипотезе, митохондрии, пластиды и базальные тельца ресничек и жгутиков эукариотической клетки были когда-то свободноживущими прокариотическими клетками. Органоидами они стали в процессе симбиоза. В пользу этой гипотезы свидетельствует наличие собственных РНК и ДНК в митохондриях и хлоропластах. По строению РНК митохондрии сходны с РНК пурпурных бактерий, а РНК хлоропластов ближе к РНК цианобактерий.
Жизнь можно определить как активное, идущее с затратой полученной извне энергии, поддержание и самовоспроизведение специфической структуры.
Согласно взглядам одного из основоположников танатологии М. Биша, жизнь — это совокупность явлений, сопротивляющихся смерти.С точки зрения второго начала термодинамики, жизнь — это процесс, или система, вектор развития которой противоположен по направлению остальным, «неживым» объектам вселенной, и направлен на уменьшение собственной энтропии
Фридрих Энгельс дал следующее определение: «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка.»
Опираясь на современные достижения биологической науки, русский ученый М. В. Волькенштейн дал новое определение понятию жизнь: «Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров — белков и нуклеиновых кислот.»
В. Н. Пармон дал следующее определение: «Жизнь — это фазово-обособленная форма существования функционирующих автокатализаторов, способных к химическим мутациям и претерпевших достаточно длительную эволюцию за счёт естественного отбора».
В настоящее время не вызывает сомнения тот факт, что жизнь, прежде чем она достигла современного многообразия, прошла длительный путь эволюции.
ЦИТОЛОГИЯ
Клеточная теория, основные ее положения. История становления и современное содержание. Значение клеточной теории в развитии биологии и медицины. Вклад отечественных и зарубежных ученых в учение о клетке.
Клеточная теория Шванна. Немецкий зоолог Т. Шванн (1810-1882) в 1839 г. опубликовал труд «Микроскопические исследования о соответствии в структуре и росте животных и растений». В этой классической работе были заложены основы клеточной теории. Шванн нашел верный принцип сопоставления клеток растительных и животных организмов. Он установил, что хотя клетки животных крайне разнообразны и значительно отличаются от клеток растений, ядра во всех клетках обладают большим сходством. Если в каком-либо видимом под микроскопом образовании присутствует ядро, это образование, по мнению Шванна, можно считать клеткой. Основываясь на таком критерии, Шванн выдвинул основные положения клеточной теории: 1) клетка является главной структурной единицей всех организмов (растительных и животных); 2) процрсс образования клеток обусловливает рост, развитие и дифференцировку растительных и животных тканей.
Развитие клеточной теории Р. Вирховом. В 1858 г. вышел в свет основной труд немецкого патолога Р. Вирхова (1821—1902) «Целлюлярная патология». Это произведение, ставшее классическим, оказало, влияние на дальнейшее развитие учения о клетке и для своего времени имело большое прогрессивное значение. До Вирхова основу всех патологических процессов видели в изменении состава жидкостей и борьбе нематериальных сил организма. Вирхов подошел к объяснению патологического процесса материалистически, показав связь его в организме с морфологическими структурами, с определенными изменениями в строении клеток. Это исследование положило начало новой науке — патологии, которая является основой теоретической и клинической медицины. Вирхов ввел в науку ряд новых представлений о роли клеточных структур в организме.
Положение Вирхова «каждая клетка из клетки» — блестяще подтвердилось дальнейшим развитием биологии. В настоящее время неизвестны иные способы появления новых клеток, помимо деления уже существующих. Однако этот тезис не отрицает того факта, что на заре жизни клетки развились из образований, еще не имевших клеточной структуры.
Положение Вирхова о том, что вне клеток нет жизни, тоже не потеряло своего значения. В многоклеточном организме имеются неклеточные структуры, но они — производные клеток. Примитивные организмы — вирусы — приобретают способность к активным процессам жизнедеятельности и размножению лишь после проникновения в клетку.
Важным обобщением явилось также утверждение, что наибольшее значение в жизнедеятельности клеток имеют не оболочки, а их содержимое: протоплазма и ядро.
Однако представления Вирхова не были лишены ошибок. Уже у Шванна проявилась тенденция рассматривать организмы как своеобразную сумму составляющих их клеток. Вирхов и особенно его последователи не только не отказались от этого положения, но и развили его дальше. Так, известный немецкий зоолог-дарвинист Э. Геккель (1834—1919) рассматривал всякий многоклеточный организм как некое «государство» клеток, в котором каждая клетка «живет» своей самостоятельной жизнью. Отсюда вытекало ошибочное мнение, что патологический процесс в организме представляет собой сумму нарушении жизнедеятельности отдельных клеток, что это —локальный (местный) процесс.
Вирхов и его последователи не видели также качественного различия между частью и целым, рассматривая организм вне его исторического развития и условий существования. Вирховскую концепцию критиковали русские естествоиспытатели и клиницисты И. М. Сеченов (1829—1905), С. П. Боткин (1832—1889) и И. П. Павлов (1849— 1936). И. М. Сеченов уже в 1860 г. отметил, что Вирхов изучает организм оторвано от среды, а органы — от организма. Русские клиницисты и физиологи своими исследованиями показали, что организм — единое целое и что интеграция его частей осуществляется, в первую очередь, нервной системой. И. П. Павлов установил ведущую координирующую роль центральной нервной системы в организме. Оказалось, что обмен веществ, питание органов и клеток находятся также под контролем нервной системы.
В настоящее время наука располагает большим фактическим материалом, убеждающим в том, что не только процессы жизнедеятельности, но также форма и величина клеток, как и другие морфологические особенности каждой клетки, связаны с теми процессами, которые протекают в организме. Единство частей целого обусловлено нервной и гуморальной регуляцией.
В целом появление «Целлюлярной патологии» Вирхова следует рассматривать как важную веху в истории биологии и медицины. Освобожденная от механистических ошибок и дополненная позднейшими открытиями, она легла в основу современных представлений о клеточном строении организма.
4.Клетка: определение. Основные типы организации клеток: про- и эукариотические клетки, общие черты и различия. Теория происхождения эукариотических клеток, ее доказательства. Неклеточные формы жизни.
Клетка — элементарная биологическая система, способная к самообновлению, самовоспроизведению и развитию. Клеточные структуры лежат в основе строения растений и животных. Каким бы многообразным ни представлялось строение организмов, в основе его лежат сходные структуры—клетки. Среди современных организмов можно последовательно проследить формирование клетки в процессе эволюции органического мира — от прокариотов, таких, как микоплазма и дробянки (общее название бактерий и синезеленых водорослей), к эукариотам. В отношении прокариот и животных типа простейших понятия «клетка» и «организм> совпадают. Их называют одноклеточными. Одноклеточными являются также некоторые виды- водорослей и грибов. Большинство растений и животных состоят из многих клеток; они получили название многоклеточных. У многоклеточных организмов клетки образуют ткани, входящие в состав органов. Жизнедеятельность клеток у многоклеточных подчинена координирующему влиянию целостного организма. Координация у животных осуществляется нервной системой и гуморальными факторами, т. е. жидкостями, циркулирующими в организме, а у растений — непосредственной цитоплазматической связью между клетками и циркулирующими веществами (фитогормонами).
Прокариоты — доядерные организмы, не имеющие типичного ядра, заключенного в ядерную мембрану. Генетический материал представлен единственной нитью ДНК, образующей кольцо,— генофором. Эта нить не приобрела еще сложного строения, характерного для хромосом, в ней нет белков-гистонов. Деление клетки только амитотическое. В клетке прокариотов отсутствуют митохондрии, центриоли, пластиды, развитая система мембран. Из организмов, имеющих клеточное строение, наиболее примитивны микоплазмы. Это бактериоподобные существа, ведующие паразитический или сапрофитный образ жизни. По размерам микоплазма приближается к вирусам. Самые мелкие клетки микоплаз-мы крупнее вируса гриппа, но мельче вируса коровьей оспы. Так, если вирус гриппа имеет диаметр от 0,08 до 0,1 мкм, а вирус коровьей оспы — от 0,22 до 0,26 мкм, то диаметр «клеток» микоплазмы — возбудителя повального воспаления легких рогатого скота — от 0,1 до 0,2 мкм.
В отличие от вирусов, осуществляющих процессы жизнедеятельности только после проникновения в клетки, микоплазма способна проявлять жизнедеятельность, свойственную организмам, имеющим клеточное строение. Эти бактериоподобные существа могут расти и размножаться на синтетической среде. Их «клетка» построена из сравнительно небольшого числа молекул (около 1200), но имеет полный набор макромолекул, характерных для любых клеток (белки, ДНК и РНК) и содержит около 300 различных ферментов.
По некоторым признакам «клетки» микоплазмы ближе стоят к клеткам животных, чем растений. Они не имеют жесткой оболочки, окружены гибкой мембраной; состав липидов близок к таковому клеток животных. Как уже сказано, к прокариотам относятся бактерии и синезеленые водоросли, объединяемые общим термином «дробянки». Клетка типичных дробянок покрыта оболочкой из целлюлозы. Дробянки играют существенную роль в круговороте веществ в природе: синезеленые водоросли — как синтетики органического вещества, бактерии — как минерализирующие его. Многие бактерии имеют медицинское и ветеринарное значение как возбудители заболеваний. Эукариоты — ядерные организмы, имеющие ядро, окруженное ядерной мембраной. Генетический материал сосредоточен преимущественно в хромосомах, имеющих сложное строение и состоящих из нитей ДНК и белковых молекул. Деление клеток митоти-ческое. Имеются центриоли, митохондрии, пластиды. Среди эукариотов существуют как одноклеточные, так и многоклеточные организмы.
троение вирусов. Наряду с одно- и многоклеточнымиорганизмами в природе существуют и другие формы жизни. Таковыми являютсявирусы, не имеющие клеточного строения. Они представляют собой переходную форму между неживой и живой материей.
Вирусы (лат. virus — яд) были открыты в 1892 г. русским ученым Д. И. Ивановским при исследовании мозаичной болезни листьев табака.
Каждая вирусная частица состоит из РНКили ДНК, заключенной в белковую оболочку, которую называюткапсидом. Полностью сформированная инфекционная частица называется вирионом. У некоторых вирусов (например, герпеса или гриппа) есть еще и дополнительная липопротеидная оболочка, возникающая из плазматической мембраны клетки хозяина.
Поскольку в составе вирусов присутствует всегда один типнуклеиновой кислоты— ДНК или РНК, вирусы делят также на ДНК-содержащие и РНК-содержащие. При этом наряду с двухцепочечными ДНК и одноцепочечными РНК встречаются одноцепочечные ДНК и двухцепочечные РНК. ДНК могут иметь линейную и кольцевую структуры, а РНК, как правило, линейную. Подавляющее большинство вирусов относится к РНК-типу.
Вирусы способны размножаться только в клетках других организмов. Вне клеток организмов они не проявляют никаких признаков жизни. Многие из них во внешней среде имеют форму кристаллов. Размеры вирусов колеблются в пределах от 20 до 300 нм в диаметре.
Хорошо изучен вирус табачной мозаики, имеющий палочковидную форму и представляющий собой полый цилиндр. Стенка цилиндра образована молекулами белка, а в его полости расположена спираль РНК (рис. 5.2). Белковая оболочка защищает нуклеиновую кислоту от неблагоприятных условий внешней среды, а также препятствует проникновению ферментов клеток к РНК и ее расщеплению.

Рис. 5.2. Схема строения вируса (а) и бактериофага (б); 1— нуклеиновая кислота; 2 — белковая оболочка; 3 — полый стержень; 4 — базальная пластинка; 5 — отростки (нити).
Молекулы вирусной РНК могут самовоспроизводиться. Это означает, что вирусная РНК является источником генетической информации и одновременно иРНК. Поэтому в пораженной клетке в соответствии с программой нуклеиновой кислоты вируса на рибосомах клетки хозяина синтезируются специфические вирусные белкии осуществляется процесс самосборки этих белков с нуклеиновой кислотой в новые вирусные частицы.Клеткапри этом истощается и погибает. При поражении некоторыми вирусами клетки не разрушаются, а начинают усиленно делиться, часто образуя у животных, в том числе и человека, злокачественные опухоли.
Бактериофаги. Особую группу представляют вирусы бактерий — бактериофаги, или фаги, которые способны проникать в бактериальную клетку и разрушать ее.
Тело фага кишечной палочкисостоит из головки, от которой отходит полый стержень, окруженный чехлом из сократительного белка. Стержень заканчивается базальной пластинкой, на которой закреплены шесть нитей (см. рис. 5.2). Внутри головки находится ДНК. Бактериофаг при помощи отростков прикрепляется к поверхности кишечной палочки и в месте соприкосновения с ней растворяетс помощью фермента клеточную стенку. После этого за счет сокращения головки молекула ДНК фага впрыскивается через канал стержня в клетку. Примерно через 10—15 мин под действием этой ДНК перестраивается весьметаболизмбактериальной клетки, и она начинает синтезировать ДНК бактериофага, а не собственную. При этом синтезируется и фаговый белок. Завершается процесс появлением 200— 1 000 новых фаговых частиц, в результате чегоклеткабактериипогибает.
Бактериофаги, образующие в зараженных клетках новое поколение фаговых частиц, что приводит к лизису (распаду) бактериальной клетки, называются вирулентными фагами.
Некоторые бактериофаги внутри клетки хозяина не реплицируются. Вместо этого их нуклеиновая кислота включается в ДНК хозяина, образуя с ней единую молекулу, способную к репликации. Такие фаги получили название умеренных фагов или профагов.
5.Основные структурные компоненты клетки. Структура и функция цитоплазмы. Органеллы животной и растительной клетки: определение, классификация. Включения: определение, виды.
Структура и функция компонентов клетки. Как правило, клетки обладают микроскопическими размерами. Части клетки, выполняющие различные функции,— органоиды— имеют микроскопические и субмикроскопические размеры. Диаметр большинства клеток колеблется от 0,01 до 0,1 мм (или от 10 до 100 мкм). Диаметр самых мелких клеток животных равен 4 мкм. Объем большинства клеток человека находится в пределах 200—15 000 мкм3. Однако известны и очень крупные клетки, видимые невооруженным глазом. Величина клеток зависит от выполняемых ими функций. Так, яйцеклетки благодаря накоплению в них питательных веществ достигают больших размеров. У многих растений (арбуз, помидор, лимон и др.) крупные размеры имеют клетки плодов, включающие вакуоли с клеточным соком.
Размеры клеток прямо не связаны с величиной организма. Так, клетки печени и почек у лошади, крупного скота и мыши имеют примерно одинаковую величину. Величина органов, как и размеры целого организма животных и растений, зависит от числа клеток.
Форма клеток также обусловлена выполняемыми ими функциями. Мышечные клетки вытянуты. Клетки покровной ткани многоугольны. Нервные клетки благодаря большому числу отростков приобрели звездчатую форму. Свободно подвижные лейкоциты имеют округлую и могут принимать амебоидную форму и т. д.
Число клеток, строящих организм, разнообразно: от одной (у протестов) или небольшого числа (у коловраток и круглых червей) до многих миллиардов, как у большинства многоклеточных.
Структурные компоненты цитоплазмы. Строение клеток животных и растений в основных чертах сходно. В теле клетки — протоплазме — различают цитоплазму и кариоплазму. Цитоплазма и кариоплазма (ядро) — обязательные составные части клетки. При удалении ядра клетка длительно существовать не может; точно так же ядро, выделенное из клетки, погибает.
Цитоплазма составляет основную массу клетки. При рассматривании живой клетки в световом микроскопе цитоплазма представляется гомогенной, бесцветной, прозрачной вязкой жидкостью. Однако электронный микроскоп позволил увидеть тонкую структуру цитоплазмы (рис. 2.2). В цитоплазме различают гиалоплазу — цитоплазматический матрикс, органоиды и включения.
Цатоплазматаческий матрикс. Основное вещество клетки составляет цитоплазматический матрикс, или гиалоплазма. С ним связаны коллоидные свойства цитоплазмы, ее вязкость, эластичность, сократимость, внутреннее движение. По химическому составу цитоплазматический матрикс построен преимущественно из белков; в состав его входят ферменты. Под электронным микроскопом цитоплазматическиймат-рикс представляется однородным тонкозернистым веществом. Иногда обнаруживаются тонкие нити (толщиной менее 10 нм) или пучки их. Даже в одной клетке разные участки цитоплазматического матрикса могут иметь неодинаковую макромолекулярную структуру.
Функционально цитоплазматический матрикс является внутренней средой клетки, местом осуществления внутриклеточного обмена. В нем осуществляется гликолиз, с которым связан поток энергии. В цитоплазматическом матриксе расположены структуры клетки — органоиды, ядра и включения
Органоиды— это постоянные дифференцированные участки цитоплазмы, имеющие определенные функции и строение. Различают органоиды общего значения и специальные. Специальные органоиды характерны для клеток, выполняющих определенные функции: миофибрилы, с которыми связано сокращение мышечных клеток, реснички эпителия в трахеях и бронхах, микроворсинки всасывающей поверхности эпителия клеток тонких кишок и т.д. К органоидам общего значения относятся: эндоплазматическнй ретикулум, рибосомы, лизосомы, митохондрии, пластинчатый комплекс, клеточный центр (центросома), микротрубочки, пластиды.
Эндоплазматическая сеть, или вакуолярная система, обнаружена в клетках всех растений и животных, подвергнутых исследованию под электронным микроскопом. Она представляет собой систему мембран, формирующих сеть канальцев и цистерн. Эндоплазматическая сеть имеет большое значение в процессах внутриклеточного обмена, так как увеличивает площадь «внутренних поверхностей» клетки, делит ее на отсеки, отличающиеся физическим состоянием и химическим составом, обеспечивает изоляцию ферментных систем, что, в свою очередь, необходимо для их последовательного вступления в согласованные реакции. Непосредственным продолжением эндоплазматической сети являются ядерная мембрана, отграничивающая ядро от цитоплазмы, и наружная мембрана (плазмалемма), расположенная на периферии клетки.
В совокупности внутриклеточные канальцы и цистерны образуют целостную систему, называемую некоторыми исследователями вакуолярной. Наиболее развита вакуолярная система в клетках с интенсивным обменом веществ.
Предполагают ее участие в активном перемещении внутри клетки жидкостей, как тех, которые синтезируются в клетке, так и поступающих извне.
Часть мембран несет на себе рибосомы, на другой части рибосом нет. В связи с этим различают два типа эндоплазматической сети — гранулярную и гладкую. С гранулярной эндоплазматической сетью связан синтез белков. В одних специальных лишенных гранул вакуолярных образованиях происходит синтез жиров, в других — гликогена. Ряд частей эндоплазматической сети связан с пластинчатым комплексом Гольджи и, по-видимому, имеет отношение к выполняемым им функциям.
Образования вакуолярной системы очень лабильны и могут меняться в зависимости от физиологического состояния клетки, характера обмена и при дифференцировке.
Рибосомы— небольшие сферические тельца, имеющие размеры от 15 до 35 нм. Они расположены в цитоплазматическом матриксе, а также связаны с мембранами эндоплазматической сети.
Наибольшее количество рибосом обнаружено в клетках, интенсивно синтезирующих белок. Рибосомы любых органов — от бактерий до млекопитающих — характеризуются сходством структуры и состава. В состав их входят белок и так называемая рРНК. Каждая из рибосом состоит из двух неравных частей — субъединиц. В каждой из субъединиц находится по молекуле РНК в виде свернутого в спираль тяжа, между витками которого находится белок. Кроме того, рибосомы содержат магний.
Молекулы информационной РНК (иРНК), синтезированные в ядре, поступают к рибосомам. Из цитоплазмы молекулами транспортных РНК (тРНК) к меньшей субъединице рибосом доставляются аминокислоты. Из них с участием ферментов полимераз и АТФ здесь синтезируются белки. Образующаяся белковая цепочка выстраивается в большей субъединице.
Рибосомы обычно объединены в группы — полисомы (или полирибосомы) — от 5 до 70 рибосом. Считается, что рибосомы формируются ядрышками и затем из ядра поступают в цитоплазму.
Лизосом ы (гр. lisis— растворение, soma — тело) —шаровидные образования, имеющие диаметр от 0,2 до 1 мкм. В лизосомах содержатся ферменты, разрушающие большие молекулы сложных органических соединений, поступающих в клетку (белки, нуклеиновые кислоты, полисахариды). Таким образом, проникающие в клетку вещества подготавливаются ферментами лизосом к синтезу белков и других веществ.
В лизосомах подвергаются разрушению микроорганизмы и вирусы. Ферменты лизосом переваривают также отмершие структуры клетки и целые погибшие в организме клетки, т. е. выполняют процессы аутофагии клетки (гр. autos— сам, fagos— пожирание).
Лизосомы играют существенную роль в индивидуальном развитии организмов, разрушая временные органы эмбрионов и личинок например, жабры и хвост у головастиков лягушки. Они встречаются в любых растительных и животных клетках. Выделяют три группы этих органоидов: прелизосомы, собственно лизосомы и постлизосомы. В прелизосомах находятся вещества, подлежащие перевариванию, но отсутствуют ферменты. Собственно лизосомы подразделяются на пеовичные и вторичные. Первичные лизосомы содержат вновь синтезированные ферменты. Вторичные образуются в результате слияния первичных лизосом с прелизосо-мами: таким образом в них содержится как субстрат, подлежащий перевариванию, так и необходимые ферменты. В зависимости от перевариваемого материала различают вторичные лизосомы двух типов: аутосомы (переваривающие утратившие свою функцию внутриклеточные структуры) и гетерофагосомы (переваривающие вещества, поступившие в клетку). Пищеварительные вакуоли простейших и фагоцитов образуются из слившихся гетерофагосом.
Постлизосомы содержат только остатки непереваренного субстрата. Каждая лизосома ограничена плотной мембраной, изолирующей содержащиеся в ней ферменты от остальной цитоплазмы. Повреждение лизосом и выход ферментов из них в цитоплазму приводит к быстрому растворению (лизису) всей клетки.
Утрата лизосомами какой-либо из ферментативных систем приводит к тяжелым патологическим состояниям целого организма — обычно наследственным болезням. Они получили название болезней накопления, так как связаны с накоплением в лизосомах полноценных, но непереваренных веществ. Эти болезни могут проявляться в недостаточности развития скелета, ряда внутренних органов, центральной нервной системы и т. д. С дефицитом лизосомных ферментов связывают развитие атеросклероза, ожирения и других нарушений. Патологическая активность лизосом может повлечь за собой разрушение жизненно важных структур.
Митохондрии (гр. mitos — нить, chondros — зернышко) — органоиды в виде гранул, палочек, нитей, видимых в световом микроскопе. Величина митохондрий сильно колеблется от 0,5 мкм до максимальной длины — 7 мкм у палочковидных. Митохондрии встречаются обязательно во всех клетках растений и животных. Число их в клетках, выполняющих различную функцию, неодинаково и колеблется от 50 до 5000. Электронная микроскопия дала возможность изучить детали строения митохондрий. Стенка митохондрии состоит из двух мембран: наружной и внутренней: последняя имеет выросты внутрь—гребни, или кристы, делящие митохондрию на отсеки, заполненные гомогенным веществом — лттриксом. Основная функция митохондрий — окисление с последующим превращением энергии разлагаемых соединений в энергию фосфатных связей (АТФ — аденозинтрифосфат и АДФ — аденозиндифосфат). В таком состоянии энергия становится наиболее доступной для использования в жизнедеятельности клетки, в частности для синтеза веществ.
Установлено также, что в матриксе митохондрий находятся рибосомы, осуществляющие синтез белка. Таким образом, митохондрии — не только энергетические центры, но и органоид, в котором наряду с ядром и рибосомами происходят биосинтетичёские процессы.
Существует структурная связь митохондрий с ядром, особенно заметная в некоторых, переходящих к делению, клетках. В таких клетках обнаружены мельчайшие структуры в виде трубочек, соединяющих митохондрии с ядерной оболочкой. Считается, что по этим трубочкам происходит обмен веществ.
Митохондрии размножаются путем перешнуровки; при делении клетки они более или менее равномерно распределяются между дочерними клетками. Таким образом между митохондриями клеток последовательных генераций осуществляется преемственность.
Как видно из сказанного, митохондриям, в отличие от других органоидов, присуща определенная автономия внутри клетки. Они никогда не возникают наново, а всегда образуются лишь в результате деления, обладают собственной ДНК, отличающейся от ядерной по своему составу и иногда имеющей форму кольца, как у прокариот. Рибосомы митохондрий мельче цитоплазматических. На этих рибосомах синтезируются митохондриальные белки, но этот синтез можно подавить действием антибиотика хлорамфеникола. Этот антибиотик способен прекращать синтез белков в бактериях, но не оказывает такого действия на цитоплазматические рибосомы.
Перечисленные особенности митохондрий, указывающие на их сходство с прокариотами, привели к представлению о симбиотическом происхождении этого органоида. Согласно данной гипотезе, какие-то из аэробных прокариот проникли в более крупную анаэробную клетку и вели первоначально возможно даже паразитический образ жизни. В дальнейшем партнеры этого сожительства в процессе эволюции приспособились друг к другу и бывший «паразит» превратился в органоид, необходимый для существования клетки. Но, став органоидом, предки митохондрий потеряли часть своего генетического материала. В эукариотных клетках митохондриальная ДНК кодирует лишь часть митохондриальных белков, большая же часть их синтезируется вне митохондрии и связана с ядерной ДНК.
Пластинчатый комплекс Гольджи виден в световом микроскопе как специфический дифференцированный участок цитоплазмы, расположенной обычно возле ядра. В клетках высших животных он представляется как сетчатая структура, иногда в виде скопления чешуек, палочек и зернышек. Электронно-микроскопические исследования позволили убедиться, что пластинчатый комплекс построен также из мембран и напоминает стопку полых рулонов, положенных друг на друга. В его состав входит система трубочек с пузырьками на концах. В клетках растений и беспозвоночных животных пластинчатый комплекс удалось обнаружить лишь с помощью электронного микроскопа. Он образован небольшими тельцами —диктиосомами, рассеянными по всей цитоплазме.
Полагают, что основная функция пластинчатого комплекса — концентрация, обезвоживание и уплотнение продуктов внутриклеточной секреции и веществ, поступивших извне, предназначенных для выделения из клетки. С ним связаны синтез полисахаридов, липидов, образование зерен желтка в развивающихся овоцитах и формирование лизосом.
При делении клеток образование борозды деления связано с комплексом Гольджи. Часть пластинчатого комплекса из материнской клетки переходит к дочерней. Следовательно, этот органоид имеет преемственное происхождение. Образование пластинчатого комплекса заново не наблюдалось.
Клеточный центр (центросома) — органоид, отчетливо видимый в световой микроскоп, состоящий из одной или двух мелких гранул — центриолей и лучистой сферы вокруг них. С помощью электронного микроскопа установлено, что каждая центриоль — это цилиндрическое тельце длиной 0,3—0,5 мкм и диаметром около 0,15 мкм. Стенки цилиндра состоят из 9 параллельно расположенных трубочек. Ох. центриолей под углом отходят отростки, которые, по-видимому, являются дочерними центриолями.
Клеточный центр иногда занимает геометрический центр клетки (откуда происходит название органоида). Чаще же он оттеснен ядром или включениями к периферии, но обязательно располагается вблизи ядра по одной оси с центром ядра и центром клетки. Активная роль клеточного центра обнаруживается при делении клетки. Расходясь в противоположные стороны, центриоли формируют полюсы делящейся клетки. По-видимому, с его структурами связаны участки цитоплазмы, способные к активному движению. Образование новых центриолей происходит путем отпочковывания от родительской. Сначала образуется небольшой зачаток, который постепенно увеличивается и, наконец, полностью сформировавшись, отделяется от материнского органоида.
Микротрубочки — длинные тонкие цилиндры, имеющие диаметр около 24 нм. Оболочка микротрубочек трехслойная, толщиной около 5 нм. Микротрубочки формируются в результате полимеризации белка тубули-на. В делящихся клетках они образуют нити веретена, входят в состав ресничек и жгутиков подвижных клеток, т. е. структур, связанных с движением, и содержат фермент АТФ-азу. Кроме того, они играют опорную роль, являясь как бы цитоскелетом, поддерживающим определенную форму всей клетки и ее органоидов, а также принимают участие в транспорте воды, ионов и некоторых молекул.
Пластиды — органоиды, характерные для клеток растений и отсутствующие в клетках животных. Не имеют пластид также клетки грибов, бактерий и синезеленых водорослей.
Репродукция пластид происходит под контролем содержащейся в них ДНК. Пластиды ранних стадий развития — пропластиды — сходны с митохондриями, имеющими малое число крист. Предполагается, что пластиды имеют симбиотическое происхождение, произошли от синезеленых водорослей, вступивших в симбиоз с первичной эукариотической клеткой.
Цитоплазматические мембраны. При изучении различных клеток животных, растений -и бактерий всегда обнаруживается, что клеточные органоиды имеют в основе своей мембранные структуры. Они характерны для эндоплазматической сети, пластинчатого комплекс", оболочек » крист митохондрий, лизосом, вакуолей, пластид, ядерной оболочки и наружной клеточной мембраны.
Современная цитология рассматривает цитомембраны как один из основных компонентов клеточной организации. Цитоплазматическая мембрана — сложная система, ответственная за основные процессы жизнедеятельности: разделение содержимого клетки на отсеки, или клеточные каналы (вакуоли, канальцы, цистерны), благодаря чему в клетке одномоментно могут
протекать различные, даже антагонистические, процессы; осуществление регуляции метаболических потоков; поддержание разности концентраций веществ (ионы, метаболиты) путем перемещения против градиента концентрации (активный перенос); создание разности электрических потенциалов; участие в процессах синтеза и катализа. Кроме того, мембраны являются стромой для точного размещения ферментов и, следовательно, обусловливают упорядоченность обменных реакций. Так, в эндоплазматической сети происходит синтез белков, жирных кислот и фосфолипидов. В митохондриях осуществляются цикл Кребса, окислительное фосфорилирование, окисление жирных кислот. В плазматической (наружной) мембране в связи с иммунологическими процессами могут протекать гликолитические реакции. Большинство заболеваний человека и животных связаны с нарушением в строении и функциях мембран.
Как показали комплексные цитофи-зические исследования, элементарная мембрана состоит из трех слоев, включающих в себя молекулы белков и липидов. Толщина каждого слоя около 2,5 нм. Часть белковых молекул обладает ферментативными свойствами. Каждая молекула липида имеет водорастворимую и водонерастворимую группы. В клеточных мембранах ли-пидные молекулы располагаются водо-нерастворимыми концами друг к другу, а водорастворимыми направлены к белковым молекулам.
Единого мнения о молекулярной организации мембран нет. По одним представлениям белковые молекулы плотно прилегают друг к другу и представляют наружные слои, по другим — белки не образуют слоя, а в виде мозаики из глобул расположены неравномерно; при этом одни из них находятся только на поверхности, другие погружены в липидную фазу частично или полностью, иногда пронизывая ее насквозь. Большая часть белковых молекул не связана с липидными молекулами и только плавает между ними. Согласно третьей гипотезе, в состав мембран
кроме липидов и белков входят также молекулы гликолипидов и гликопротеидов с разветвленными углеводными цепями. Эти разветвленные цепи на поверхности мембраны переплетаются друг с другом, образуя как бы каркас с вплетенными в него молекулами белков. Более того, углеводные цепи гликолипидов и гликопротеидов связаны с микротрубочками, составляющими цитоскелет. Часто плазматическая мембрана образует множество пальцевидных выступов— микроворсинок. Это значительно увеличивает всасывающую поверхность клеток, облегчая перенос веществ через наружную мембрану и их прикрепление к поверхности субстрата.
Существует, по-видимому, несколько типов мембран, отличающихся по строению в ферментативными свойствами белков, образующих с липидами липопротеидные комплексы. С этим связаны неодинаковые функциональные свойства мембран различных органоидов и различных участков клетки. Так, мембраны митохондрий тонки (около 5 нм) и имеют глобулярную структуру; мембраны сетчатого аппарата толще (6—8 нм), содержат липид-ные и фосфорные молекулы. В мембранах находятся молекулы-рецепторы, благодаря чему они восприимчивы к биологически активным соединениям, например, гормонам.
Наружная или плазматическая мембрана (цитолемма или плазмолемма) ограничивает клетку от окружающей микросреды и благодаря наличию молекул-рецепторов обеспечивает целесообразные реакции клетки на изменения в окружающей ее среде. Она принимает непосредственное участие в процессах обмена, клетки со средой — поступлении веществ в клетку и выведении их из нее. Она никогда не находится в состоянии покоя, совершая обычно волнообразные колебательные движения.
В тканях растений между соседними клетками образуются цитоплазматические мостики — плозмодесмы, через которые обеспечивается взаимосвязь лежащих рядом клеток. В растительных клетках цитоплазматическая мембрана снаружи покрыта клеточной оболочкой.
6. Наследственный аппарат клетки; роль ядра и цитоплазмы в передаче наследственной информации. Ядро, его значение для жизнедеятельности клеток, основные компоненты ядра. Цитоплазматическая наследственность: плазмиды и эписомы.
Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую – с ее реализацией. Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК), осуществляющий основные функции: хранение, передача и реализация наследственной информации с обеспечением синтеза белка.
В первую группу входят процессы, связанные с поддержанием информации в виде неизменной структуры ДНК. Важно, чтобы молекулы ДНК передавались из поколения в поколение стабильными, а поскольку это невозможно в силу мутагенеза, существуют репарационные механизмы на молекулярном уровне, т.е. самовосстановление первичной структуры. Существует световая репарация: при облучении видимым светом (УФ) активируется фермент, восстанавливающий первичную структуру ДНК за счет расщепления образовавшихся в мутированной молекуле димеров пиримидиновых оснований.
При темновой репарации происходит вырезание димеров пиримидинов с помощью эндонуклеазы, далее к интактной цепи ДНК присоединяются комплементарные нуклеотиды и цепь сливается лигазами с получением исходной структуры.
Чтобы дочерние клетки при делении (митозе) получили совершенно одинаковые в количественном и качественном отношении объемы генетической информации, в ядре должна пройти редупликация молекул ДНК, что и наблюдается в S-периоде интерфазы.
Во время образования половых клеток происходят рекомбинации генетической информации, что обеспечивает их генетическую разнородность при одинаковом количественном объеме (кроссинговер при редукционном делении).
Далее, в функции ядра входит распределение генетической информации между дочерними клетками, для чего в ядре происходит предварительная компактизация хромосом (выше описана).
Для реализации генетической информации требуется создание собственно аппарата белкового синтеза. Это включает включает синтез на молекулах ДНК разных информационных РНК, транспортных и рибосомных РНК. Кроме того в ядре эукариотических клеток происходит образование субъединиц рибосом путем образования комплексов рибосомных белков и рибосомных РНК, которые затем переходят в цитоплазму и на мембраны ЭПС, где и функционируют. Коллинеарногенетическому коду, через транскрипцию и трансляцию, конечным результатом реализации генетической информации является синтез полипептидных цепей в рибосоме. Такая однонаправленность и универсальность может быть представлена в виде схемы, известной как «центральная догма молекулярной биологии» ДНК:
ДНК → репликация → ДНК → транскрипция → РНК → трансляция → полипептид → эпигенез → белок → признак.
Таким образом, ядро представляет собой не только вместилище генетической информации (хорошо защищенной ядерной мембраной), но и место где этот материал воспроизводится и функционирует. Поэтому выпадение или нарушение любой из перечисленных функций гибельно для клетки в целом. Так, нарушение репарационных процессов будет приводить к изменению первичной структуры белков до несвойственных данной клетке, что проявится в виде патологии или гибели.
Нарушение процессов распределения генетического материала приведет к грубым нарушениям в кариотипе, летальным исходам или наследственным заболеваниям типа синдромов Тернера, Патау, Эдвардса и других с неблагоприятным прогнозом.
На организменном уровне, ведущая роль ядра проявляется и в поддержании гомеостаза. Живой организм, будучи открытой системой, на любом этапе индивидуального развития существует в единстве со средой обитания, при этом, адекватно реагируя на изменяющиеся условия, сохраняет себя как отдельную биологическую систему, Свойство живых форм поддерживать генетическую конструкцию, структурные показатели, постоянство внутренней среды закреплено генетически и сложилось в процессе эволюции. Эффективность механизмов гомеостаза определяется генотипами особей, т.е. опять же, характером генов, молекул ДНК, нормой реакции на изменение окружающей среды. Появление в клетках чужеродной информации, как результат мутаций под влиянием биологических (вирусы, бактерии), химических (пестициды, гербициды и т.д.), физических (радиация УФ и т.д.) воздействий, оказывает отрицательное действие и изменение показателей гомеостаза. Регуляция гомеостаза на клеточном уровне идет при участии ядра, цитоплазматической мембраны, рибосом, АТФ. Клетка содержит цитоплазму, состав которой модулируется избирательной проницаемостью клеточной мембраны и активностью ферментов, они в свою очередь образуются в результате считывания информации с ДНК (с участков ДНК-генов). «Включение» и «выключение» генов контролируется системами индукции и репрессии. В основе регуляции работы генов лежит репрессионно-депрессивный механизм (Жакоб, Моно, 1961г.). У многоклеточных эукариотических организмов роль регуляторов могут выполнять гормоны, которые диффундируют через клеточные мембраны (из межклеточной жидкости) и связываются с белками рецепторами в цитоплазме. Образующиеся комплексы транспортируются в ядро к начальному звену оперона – оператору, после чего со структурных генов транскрибируется про-и-РНК и запускается механизм синтеза белка, включающегося в обмен веществ и , в конечном итоге происходит коррекция в метаболизме и развитие адаптации в изменившихся условиях.
Важнейшая роль цитоплазмы — объединение всех клеточных структур (компонентов) и обеспечение их химического взаимодействия. Она выполняет и другие функции, в частности, поддерживает тургор клетки.
В процессе реализации заключенной в ядерных генах генетической информации важную роль играет цитоплазма клетки. Именно в цитоплазме осуществляется синтез белковых молекул на основе информации, закодированной в молекулах ядерной ДНК. Одновременно некоторые структурные элементы цитоплазмы могут хранить и передавать по наследству определенную долю генетической информации, не связанной с ядром. Такой способ передачи генетической информации называется цитоплазматической, или нехромосомной, наследственностью.
Цитоплазматическая наследственность связана с действием генов, локализованных в таких элементах цитоплазмы, которые содержат ДНК, способны к автономной репликации и равномерному распределению между дочерными клетками. Важнейшими из них являются пластиды, митохондрии и плазмиды.
Плазми́ды — небольшие молекулы ДНК, физически отдельные от геномных хромосом и способные реплицироваться автономно. Как правило, плазмиды встречаются у бактерий и представляют собой двухцепочечные кольцевые молекулы, но изредка плазмиды встречаются также у архей и эукариот.
Эписомы — это генетические элементы бактерий, способные функционировать в клетке независимо от бактериальной хромосомы. Эписомы представляют собой молекулы ДНК. Они определяют в бактериях ряд признаков, важнейшим из которых является устойчивость к антибиотикам и сульфаниламидным препаратам.
7.Роль хромосом в передаче наследственной информации (косвенные и прямые доказательства). Правила хромосом. Современные представления о строении хромосом. Уровни организации ДНК в хромосомах. Хроматин – как форма существования хромосом: строение и химический состав.
В ядре хромосомы являются материальными носителями информации на клеточном кровне.
Прямыми доказательствами этого являются наследственные болезни, связанные с нарушением числа и структуры хромосом.
Косвенными доказательствами этой функции хромосом являются правила хромосом:
- Правило постоянства числа хромосом. Число хромосом и особенности их строения – видовой признак
- Правило парности хромосом. Число хромосом в соматических клетках всегда четное, это связано с тем, что хромосомы составляют пары, т.к. одна хромосома при половом размножении идет от отцовского организма, а вторая от материнского. Хромосомы, относящиеся к одной паре, одинаковые по величине, форме и расположению центромер называются гамологичными.
- Правило индивидуальности хромосом. Каждая пара хромосом характеризуется своими особенностями. Негомологичные хромосомы всегда имеют ряд отличий.
- Правило непрерывности хромосом. Хромосомы способны к авторепродукции (в результате репликации ДНК). «Дочерние» хромосомы образуются в результате расхождения хроматид материнской хромосомы в анафазу митоза или мейоза 2, что обеспечивает непрерывную передачу наследственной информации при делении клеток.
Компактизация, спирализация или укладка ДНК в хромосому происходит следующим образом: выделяют несколько уровней укладки ДНК в хромосому:
нуклеосомный
нуклеомерный
хромомерный
хромонемный
хромосомный
Структурно-функциональной единицей хромосом на молекулярном уровне является – нуклеосома.
Сердцевиной нуклеосомы является октамер из 8 молекул гистоновых белков. Это так называемый «нуклеосомный кор». Молекула ДНК накручивается на октамер. Через линкерный участок связанный с гистоновым белком ДНК переходит на другую нуклеосому, образуя так называемые «бусинки на нитке».Примерно 90 % ДНК входит в состав нуклеосом, а 10 % на линкерные участки между нуклеосомами. Количество нуклеосом в ядре огромно. Этот уровень обеспечивает сверхскручивание ДНК на поверхности гистоновой сердцевины и укорочение ДНК в 7 раз.
Нуклеомерный уровень укладки хроматина обеспечивает сорокократное укорочение ДНК. Как нуклеосомный, так и нуклеомерный уровень уровень компактизации ДНК хроматина осущетвляется за счет гистоновых белков. Нуклеосомная фибрилла скручивается в спираль. Нуклеомерный тип укладки заключается в том, что 8-10 нуклеосом объединяются в нуклеомер («сверхбусина»). В результате такой упаковки образуется хроматиновое волокно, которое подвергается дальнейшей компактизации с уменьшением длины в 100 раз. Все остальные уровни компактизации связаны с укладкой хроматиновых фибрилл в новые структуры, где ведущую роль играют негистоновые белки.
Негистоновые белки связываются с особыми участками ДНК, которая в местах связывания образует большие петли или домены. Петли доменов заякорены на внутреннем поддерживающем матриксе – ламине, которая прилегает к внутренней ядерной мембране. Следующие более высокие уровни компактизации ДНК связаны не с ее дополнительной спирализацией, а с образованием поперечной петельной структуры.
Белки ядерного матрикса формируют не сплошной остов по длине хромосомы, а множество отдельных центров, к которым крепятся петли ДНК, образуя розетки (хромомеры)
Белки образуют в центре хромосомы непрерывный тяж, к которому крепятся петли нуклеомеров. Затем сближенные хромомеры образуют толстые нити. Эти образования называют хромонемы.
И последний уровень структурной организации хроматина – хроматидный. Хромонемы укладываются спирально или петлеобразно, образуя хроматиду.
Метафазная хромосома состоит из двух хроматид, соединенных первичной перетяжкой – центромерой. Таким образом, в результате в результате суперспирализации происходит компактизация ДНК и образование хромосом. Это необходимый этам организации хроматина в подготовке к клеточному делению.
Хроматин, нуклеопротеид клеточного ядра, составляющий основу хромосом. В состав хроматина входят: ДНК (30-40% по массе), гистоны (30-50%), негистоновые белки (4-33%) и РНК. В зависимости от степени конденсации (плотности упаковки) и коррелирующей с ней активности хроматина в интерфазе различают гетерохроматин и эухроматин. Гетерохроматин бывает конститутивный (структурный) и факультативный. Если для факультативного гетерохроматина конденсированное (плотно упакованное) состояние - явление временное, наступающее как следствие инактивации хроматина, например, в ходе развития или дифференцировки, то конститутивный гетерохроматин конденсирован всегда. Функции его неясны.
Эухроматин отличается от гетерохроматина менее плотной упаковкой хромосом.ого материала, большим кол-вом негистоновых белков и др. Может инактивироваться и приобретать свойства факультативного гетерохроматина.
8.Биологические мембраны: их строение и свойства. Плазмалемма: строение, функция. Клетка как открытая система. Способы проникновения веществ в клетку: сущность, значение в медицине. Пассивный путь проникновения веществ в клетку (осмос, диффузия, фильтрация). Медицинское значение изучения данных процессов в клетке. Активный путь проникновения веществ в клетку («ионный насос», пиноцитоз, фагоцитоз). Их роль для одноклеточных и многоклеточных организмов.
Биологическая мембрана - это структура, состоящая из органических молекул, которая имеет толщину около 7-10нм и видима только посредством электронного микроскопа. В каждой клетке есть плазматическая мембрана, которая ограничивает содержимое клетки от наружней среды, и внутренние мембраны, которые формируют различные органоиды клетки (митохондрии, органоиды, лизосомы и т.п.)
Плазматическая мембрана выполняет несколько важных функций.
1) Образует избирательный барьер, который отделяет содержимое клетки от окружающей среды, что позволяет поддерживать постоянными химический состав цитоплазмы и её физические свойства.
2) Регулирует транспорт веществ между содержимым клетки и окружающим клетку раствором.
3) Принимает участие в информационных процессах в живой клетке.
Химическая состав и структура плазматической мембраны
В состав плазматической мембраны входят липиды, белки и углеводы. Соотношение между липидами и белками может значительно варьировать в различных клетках.
Липиды мембраны бывают трех видов: глицерофосфолипиды, сфингофосфолипиды и стероиды (холестерол).
Белки мембраны подразделены на два вида. Молекулы первого типа являются гидрофильными. Эти белки, называемые периферическими, соединены с поверхностью мембраны сравнительно слабыми электростатическими силами. Белки второго вида имеют как гидрофильные, так и гидрофобные группы. Их молекулы более или менее погружены в мембрану, и удерживаются в ней более прочными гидрофобными силами. Некоторые белки пронизывают мембрану от ёё внутренней до внешней поверхностей - интегральные белки.
Многочисленные белки мембраны выполняют различные функции (метаболическую, транспортную, рецепторную и т.п.). Функции белков мембраны существенно зависят от строения их молекул. Плазмолемма — оболочка животной клетки, ограничивающая ее внутреннюю среду и обеспечивающая взаимодействие клетки с внеклеточной средой.
Плазмолемма имеет толщину около 10 нм, и состоит на 40 % из липидов, на 5-10 % из углеводов (в составе гликокаликса), и на 50-55 % из белков.
Функции плазмолеммы:
разграничивающая (барьерная);
рецепторная или антигенная;
транспортная;
образование межклеточных контактов.
Основу строения плазмолеммы составляет:
двойной слой липидных молекул (билипидная мембрана), в которую местами включены молекулы белков;
надмембранный слой — гликокаликс, структурно связанный с белками и липидами билипидной мембраны;
в некоторых клетках имеется подмембранный слой.
Строение билипидной мембраны
Каждый монослой ее образован в основном молекулами фосфолипидов и, частично, холестерина. При этом в каждой липидной молекуле различают две части:
гидрофильную головку;
гидрофобные хвосты.
Гидрофобные хвосты липидных молекул связываются друг с другом и образуют билипидный слой. Гидрофильные головки билипидного слоя соприкасаются с внешней или внутренней средой. Билипидная мембрана, а точнее ее глубокий гидрофобный слой, выполняет барьерную функцию, препятствуя проникновению воды и растворенных в ней веществ, а также крупных молекул и частиц.
На электроннограмме в плазмолемме четко определяются три слоя:
наружный (электронноплотный);
внутренний (электронноплотный);
промежуточный (с низкой электронной плотностью).
Белковые молекулы встроены в билипидный слой мембраны локально и не образуют сплошного слоя.
По локализации в мембране белки подразделяются на:
интегральные (пронизывают всю толщу билипидного слоя);
полуинтегральные, включающиеся только в монослой липидов (наружный или внутренний);
прилежащие к мембране, но не встроенные в нее.
По выполняемой функции белки плазмолеммы подразделяются на:
структурные белки;
транспортные белки;
рецепторные белки;
ферментные.
Находящиеся на внешней поверхности плазмолеммы белки, в также гидрофильные головки липидов обычно связаны цепочками углеводов и образуют сложные полимерные молекулы гликопротеиды и гликолипиды. Именно эти макромолекулы и составляют надмембранный слой — гликокаликс. В неделящейся клетке имеется подмембранный слой, образованный микротрубочками и микрофиламентами.
Значительная часть поверхностных гликопротеидов и гликолипидов выполняют в норме рецепторные функции, воспринимают гормоны и другие биологически активные вещества. Такие клеточные рецепторы передают воспринимаемые сигналы на внутриклеточные ферментные системы, усиливая или угнетая обмен веществ, и тем самым оказывают влияние на функции клеток. Клеточные рецепторы, а возможно и другие мембранные белки, благодаря своей химической и пространственной специфичности, придают специфичность данному типу клеток данного организма и составляют трансплантационные антигены или антигены гистосовместимости.
Помимо барьерной функции, предохраняющей внутреннюю среду клетки, плазмолемма выполняет транспортные функции, обеспечивающие обмен клетки с окружающей средой.
Различают следующие способы транспорта веществ:
пассивный транспорт — способ диффузии веществ через плазмолемму (ионов, некоторых низкомолекулярных веществ) без затраты энергии;
активный транспорт веществ с помощью белков-переносчиков с затратой энергии (аминокислот, нуклеотидов и других);
везикулярный транспорт через посредство везикул (пузырьков), который подразделяется на эндоцитоз — транспорт веществ в клетку, и экзоцитоз — транспорт веществ из клетки.
В свою очередь эндоцитоз подразделяется на:
фагоцитоз — захват и перемещение в клетку крупных частиц (клеток или фрагментов, бактерий, макромолекул и так далее);
пиноцитоз — перенос воды и небольших молекул.
Процесс фагоцитоза подразделяется на несколько фаз:
адгезия (прилипание) объекта к цитолемме фагоцитирующей клетки;
поглощение объекта путем образования вначале углубления (инвагинации), а затем и образования пузырьков — фагосомы и передвижения ее в гиалоплазму.
Жизненный цикл клетки, его периоды, их характеристика, особенности у различных видов клеток. Морфофункциональная характеристика и динамика структуры хромосом в клеточном цикле. Механизм регуляции митотической активности. Понятия о митогенах и митостатиках. Митотический индекс. Категории клеточных комплексов (растущие, обновляющиеся, статические). Главные механизмы митотического цикла, обеспечивающие поддержание генетического гомеостаза. Понятие об апоптозе.
Совокупность процессов, происходящих от образования клетки до ее гибели называется жизненным циклом.
Интерфаза состоит из нескольких периодов:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки, РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов.
2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков.
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
Регуляторные факторы, контролирующие размножение клеток можно условно разделить на две группы: экзогенные и эндогенные. Экзогенные факторы взаимодействуют с поверхностью клетки. Факторы, которые синтезируются самой клеткой и действуют внутри нее, относятся к эндогенным. Такое подразделение весьма условно, поскольку некоторые факторы, будучи эндогенными по отношению к продуцирующей их клетке, могут выходить из нее и действовать как экзогенные регуляторы на другие клетки. Если регуляторные факторы взаимодействуют с теми же клетками, которые их продуцируют, то такой тип контроля называется аутокринным.
Для изучения митотической активности и обновления клеточных комплексов применены новейшие методы: определение числа ядер, изучение изменения количественного содержания ДНК в ткани; изучение клеточного деления посредством радиоавтографии, т. е. путем включения радиоактивных изотопов в ДНК и др.
Применение названных методов позволило разделить все ткани на три категории клеточных комплексов: стабильные, растущие и обновляющиеся.
В стабильных клеточных комплексах не обнаруживаются митозы и количественное содержание ДНК остается постоянным. К таким клеткам, которые никогда не делятся, относятся клетки центральной и периферической нервной системы. Эти клетки сохраняются на протяжении всей жизни, но в них происходят возрастные изменения.
К числу растущих клеточных комплексов относятся такие группы однородных клеток, в которых всегда встречаются отдельные клетки, находящиеся в стадии митоза. Предполагается, что клетки в этих комплексах живут на протяжении всей жизни организма, а за счет вновь образующихся клеток происходит увеличение органа. Из таких клеточных комплексов состоят почки, надпочечники, щитовидная и поджелудочная железы, скелетные и сердечная мышцы.
Обновляющиеся клеточные комплексы - это группы однородных клеток с большим числом митозов. В этих комплексах число вновь образующихся клеток восполняет такое же число погибающих. Примерами обновляющихся комплексов могут служить клетки желудочно-кишечного тракта, клетки кожного эпидермиса, ткань семенников и кроветворных органов и др.
Естественная гибель клетки (апоптоз). Морфологически апоптоз характеризуется разрушением ядра и цитоплазмы. “Осколки” погибшей клетки поглощаются и перерабатываются фагоцитами. Но ведь клетки могут погибнуть и под воздействием случайных факторов (механических, химических и любых других). Случайная гибель клеток (а также ткани, органа) в биологии называется некрозом. Важно то, что естественная клеточная гибель (апоптоз) в отличие от некроза не вызывает воспаления в окружающих тканях. Апоптоз не вызывает воспаления в окружающих тканях. В организме запрограммированная клеточная гибель выполняет функцию, противоположную митозу, и, тем самым, регулирует общее число клеток в организме. Апоптоз играет важную роль в защите организма при вирусных инфекциях.
Основные способы деления клетки: митоз, мейоз и амитоз. Определение, характеристика процессов и их биологическое значение. Биологическое отличие мейоза от митоза. Митоз и проблемы роста, регенерации, опухолевого роста.
Митоз — процесс непрямого деления соматических клеток эукариот, в результате которого из одной диплоидной материнской клетки образуются две дочерние с таким же набором хромосом.
Подготовка клетки к митозу происходит в интерфазу: удваивается ДНК, накапливается АТФ, синтезируются белки веретена деления.
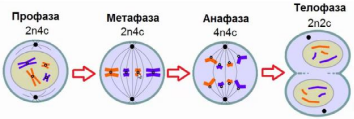
Профаза.
В ядре молекулы ДНК укорачиваются и скручиваются (спирилизуются), образуя компактные хромосомы.
Каждая хромосома состоит из двух молекул ДНК (двух хроматид), соединённых центромерой.
Ядерная оболочка распадается.
Хромосомы неупорядоченно располагаются в цитоплазме.
Растворяются ядрышки.
Начинает формироваться веретено деления, часть нитей которого прикрепляется к центромерам хромосом.
В животной клетке центриоли удваиваются и начинают расходиться.
Метафаза.
Хромосомы располагаются на экваторе клетки, образуя метафазную пластинку.
Хроматиды соединены в области первичной перетяжки с нитями веретена деления.
Центриоли располагаются у полюсов клетки.
Анафаза.
Каждая хромосома, состоящая из двух хроматид, разделяется на две идентичные дочерние хромосомы.
Дочерние хромосомы растягиваются нитями веретена деления к полюсам клетки.
У каждого полюса оказывается одинаковый генетический материал.
Телофаза.
Хромосомы раскручиваются.
Вокруг хромосом начинают формироваться ядерные оболочки.
В ядрах появляются ядрышки.
Нити веретена деления разрушаются.
На этом кариокинез завершается. Происходит цитокинез — разделение цитоплазмы
Биологическое значение митоза.
В результате митоза образуются генетически одинаковые дочерние клетки с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений.
Мейоз — это способ деления клеток, в результате которого из одной диплоидной материнской клетки образуется четыре гаплоидные дочерние клетки.
Подготовка клетки к мейозу происходит в интерфазу: удваивается ДНК, накапливается АТФ, синтезируются белки веретена деления.
Профаза I
Происходит скручивание молекул ДНК и образование хромосом. Каждая хромосома состоит из двух гомологичных хроматид — 2n4c.
Гомологичные (парные) хромосомы сближаются и скручиваются, т.е. происходит конъюгация хромосом.
Затем гомологичные хромосомы начинают расходиться.
При этом образуются перекрёсты и происходит кроссинговер — обмен участками между гомологичными хромосомами.
Растворяется ядерная оболочка.
Разрушаются ядрышки.
Формируется веретено деления.
Метафаза I
Спирилизация хромосом достигает максимума.
Пары гомологичных хромосом (четыре хроматиды) выстраиваются по экватору клетки.
Образуется метафазная пластинка.
Каждая хромосома соединена с нитями веретена деления.
Хромосомный набор клетки 2n4c.
Анафаза 1
Гомологичные хромосомы, состоящие из двух хроматид, отходят друг от друга.
Нити веретена деления растягивают хромосомы к полюсам клетки.
Из каждой пары гомологичных хромосом к полюсам попадает только одна.
Происходит редукция — уменьшение числа хромосом вдвое.
У полюсов клетки оказываются гаплоидные наборы хромосом, состоящих из двух хроматид.
Хромосомный набор у полюсов 1n2c.
Телофаза I
Происходит формирование ядер.
Делится цитоплазма.
Образуются две клетки с гаплоидным набором хромосом.
Каждая хромосома состоит из двух хроматид.
Хромосомный набор каждой из образовавшихся клеток 1n2c.
Через короткий промежуток веремени начинается второе деление мейоза. В это время не происходит удвоения ДНК. Делятся две гаплоидные клетки, которые образовались в результате первого деления.
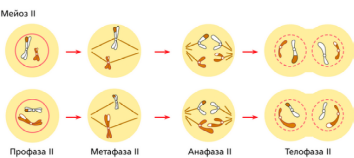
Профаза II
Ядерные оболочки разрушаются.
Хромосомы располагаются беспорядочно в цитоплазме.
Формируется веретено деления.
Хромосомный набор клетки 1n2c.
Метафаза II
Хромосомы располагаются в экваториальной плоскости.
Каждая хромосома состоит из двух хроматид.
К каждой хроматиде прикреплены нити веретена деления.
Хромосомный набор клетки 1n2c.
Анафаза II
Нити веретена деления оттягивают сестринские хроматиды к полюсам.
Хроматиды становятся самостоятельными хромосомами.
Дочерние хромосомы направляются к полюсам клетки.
Хромосомный набор у кадого полюса 1n1c.
Телофаза II
Формируются ядра.
Делится цитоплазма.
Образуется четыре гаплоидные клетки 1n1c.
Хромосомные наборы образовавшихся клеток не идентичны.
Значение мейоза.
Образовавшиеся в результате мейоза клетки отличаются своими хромосомными наборами, что обеспечивает разнообразие живых организмов.
Число хромосом при мейозе уменьшается в два раза, что необходимо при половом размножении. Процесс оплодотворения опять восстанавливает в зиготе диплоидный набор хромосом.
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
Отличия мейоза от митоза по итогам
1. После митоза получается две клетки, а после мейоза – четыре.
2. После митоза получаются соматические клетки (клетки тела), а после мейоза – половые клетки (гаметы – сперматозоиды и яйцеклетки; у растений после мейоза получаются споры).
3. После митоза получаются одинаковые клетки (копии), а после мейоза – разные (происходит рекомбинация наследственной информации).
4. После митоза количество хромосом в дочерних клетках остается таким же, как было в материнской, а после мейоза уменьшается в 2 раза (происходит редукция числа хромосом; если бы её не было, то после каждого оплодотворения число хромосом возрастало бы в два раза; чередование редукции и оплодотворения обеспечивает постоянство числа хромосом).
Отличия мейоза от митоза по ходу
1. В митозе одно деление, а в мейозе – два (из-за этого получается 4 клетки).
2. В профазе первого деления мейоза происходит конъюгация (тесное сближение гомологичных хромосом) и кроссинговер (обмен участками гомологичных хромосом), это приводит к перекомбинации (рекомбинации) наследственной информации.
3. В анафазе первого деления мейоза происходит независимое расхождение гомологичных хромосом (к полюсам клетки расходятся двуххроматидные хромосомы). Это приводит к рекомбинации и редукции.
4. В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, поскольку они и так двойные.
...
Второе деление мейоза ничем не отличается от митоза. Как и в митозе, в анафазе II мейоза к полюсам клетки расходятся одинарные сестринские хромосомы (бывшие хроматиды).
Размножение - основное свойство живого. Способы бесполого размножения. Эволюция форм полового размножения (изогамия, анизогамия, оогамия). Определение, сущность, биологическое значение.
Среди многообразных проявлений жизнедеятельности (питание, обустройство местообитания, защита от врагов) размножению принадлежит особая роль. Способность к размножению является неотъемлемым свойством живых существ. С его помощью сохраняются во времени биологические виды и жизнь как таковая. Биологическая роль размножения состоит в том, что оно обеспечивает смену поколений. Различия, закономерно проявляющиеся в фенотипах особей разных поколений, делают возможным естественный отбор и, следовательно, эволюцию жизни
Различают два способа размножения: бесполое и половое.
Бесполое размножение — форма размножения, не связанная с обменом генетической информацией между особями — половым процессом.
Деление надвое приводит к возникновению из одного родительского организма двух дочерних. Оно является преобладающей формой у прокариот и простейших, у медуз, у кольчатых червей. Множественное деление (шизогония) встречается среди простейших, в том числе паразитов человека (малярийный плазмодий). При размножении почкованием потомок формируется первоначально как вырост на теле родителя с последующей его отшнуровкой (гидра). Фрагментация заключается в распаде тела многоклеточного организма на части, которые далее превращаются в самостоятельных особей (плоские черви, иглокожие). У видов, размножающихся спорами, дочерний организм развивается из специализированной клетки-споры. В зависимости от формы бесполого размножения потомок развивается либо из одной клетки (спорообразование, шизогония, деление), либо из группы клеток родителя. В последнем случае размножение называют вегетативным. Оно распространено среди растений.
Бесполое размножение наблюдается у животных с относительно низким уровнем структурно-физиологической организации, к которым принадлежат многие паразиты человека.
Половое размножение отличается наличием полового процесса, который обеспечивает обмен наследственной информацией и создает условия для возникновения наследственной изменчивости. В нем, как правило, участвуют две особи — женская и мужская, которые образуют гаплоидные женские и мужские половые клетки — гаметы.
Формами полового процесса являются конъюгация и копуляция.
Конъюгация — своеобразная форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, образуемому двумя особями. Копуляция (гаметогамия) — форма полового процесса, при которой две различающиеся по полу клетки — гаметы — сливаются и образуют зиготу. При этом ядра гамет образуют одно ядро зиготы. Различают следующие основные формы гаметогамии: изогамия, анизогамия и оогамия. При изогамии образуются подвижные, морфологически одинаковые гаметы, однако физиологически они различаются на «мужскую» и «женскую». Изогамия встречается у многих водорослей.
При анизогамии (гетерогамии) формируются подвижные, различающиеся морфологически и физиологически гаметы. Такой тип полового процесса характерен для многих водорослей.
В случае оогамии гаметы сильно отличаются друг от друга. Женская гамета — крупная неподвижная яйцеклетка, содержащая большой запас питательных веществ. Мужские гаметы — сперматозоиды - мелкие, чаще всего подвижные клетки, которые перемещаются с помощью одного или нескольких жгутиков. Оогамия характерна для животных, высших растений и многих грибов.