-
Класс Гинковые: анатомо-морфологическая характеристика, особенности экологии, биология, география.
Класс Ginkgopsida. Платиспермическая линия эволюции. Включает несколько порядков.
Порядок Calamopityales.
-
Семена эллиптические в сечении.
-
В каждой половине интегумента проходит по одному пучку.
-
Нуцеллус срастается с интегументом почти на всю длину.
-
Вегетативные вайи сложноперистые.
-
Главный рахис вильчатый. Жилкование перышек веерное (как у археоптерисовых).
-
Стволики тонкие, эустелические.
-
По периферии сердцевины расположены несколько первичных пучков. Они, как и у археоптерисовых, делятся радиально, но уже наблюдается пазушное ветвление осей. К осевым пучкам примыкает вторичная древесина, сложенная трахеидами.
-
Кора спорганового типа, т.е. по ее периферии расположены короткие радиальные склеренхимные пластины, идущие вдоль ствола и изредка анастомозирующие. После первого дихотомического деления рахиса в каждом из ответвлений проходит V-образный пучок с несколькими протоксилемными тяжами. Далее рахисы ветвятся перисто.
-
В осях и черешках часто наблюдаются секреторные каналы.
Предполагалось, что каламопитиевые – предки лагеностомовых, но, возможно, обе группы возникли независимо от древних голосеменных с варьирующейся симметрией семени.
Порядок Callistophytales.
-
Каллистофитовые по стелярной организации сходны с каламопитиевыми.
-
Семена имеют свободный интегумент с полностью сформированным микропиле.
-
По строению семян сходны с пельтаспермовыми.
-
Строение и расположение синангиев напоминает мараттиевых.
-
Ствол плагиотропный, есть вторичная древесина.
-
Ветвление пазушное.
-
В стеблях, как у каламопитиевых, отмечены кора спорганового типа, секреторные каналы и полости.
-
Семена мелкие. Интегумент 3-слойный, уплощенный, с секреторными каналами.
-
Семяпочки выделяли пыльцеулавливающую секреторную каплю. Формы из среднего карбона несли по одному семени на нижней лопасти перышек. Формы из позднего карбона имели филлоспермы с модифицированными перышками, несшими большое число семян.
-
Синангии (на перышках) состояли из 5–9 спорангиев, сросшихся нижними частями.
-
Микрогаметофит такой же, как у современных хвойных. Сходство в строении и онтогенезе пыльцы и микроспорангия каллистофитовых и хвойных – один из примеров параллелизма в эволюции растений.
Порядок Peltaspermales.
-
Пельтасперовые схожи с каллистофитовыми, но отличаются от них тем, что семяносные листья преобразуются в кладоспермы, утрачивающие сходство с вегетативной листвой и приобретающие облик специализированных фертильных побегов.
-
Кладоспермы более примитивных родов сходны с вегетативными листьями типом рассечения. У более продвинутых родов кладоспермы чаще всего пельтатные (пельтоиды) или купуловидные. Семяносный диск венчает ножку и несет снизу семена, окружающие ее.
-
Семеносная пластинка может быть перистой с семенами, сидящими двумя рядами. Она может редуцироваться до купулы с единственным семенем (семейство ункомасиация). У семейства кардиолепидация края кладосперма в разной степени подвернуты вниз. При сильном подворачивании образуется почти закрытая капсула.
-
У мужских микроспорокладов редукция листовой пластинки шла гораздо быстрее.
-
Синангии древнейших пельтаспермовых расположены перисто.
-
У семейства ункомасиация сохраняется уплощенность пластинки, несущей спорангии. У остальных пельтаспермовых синангии сидят пучками у концов веточек, слегка сливаясь у основания.
-
Для пельтаспермовых характерны секреторные каналы, или полости, что объединяет их с каламопитиевыми, каллистофитовыми и гинкговыми.
-
Чаще всего листья перистые, с открытым жилкованием.
-
Характерна вильчатость рахиса.
-
Есть роды с цельными листьями и параллельным или веерным жилкованием.
-
В порядке выделяют четыре семейства: трихопитиация (кладоспермы сохраняют листоподобность, делятся перисто или перисто-дихотомически, семена прикрепляются субапикально к конечным долям); пельтаспермация (характерны пель- тоиды и кладоспермы); кардиолепидация (пельтатные капсулы почти замкнутые); ункомасиация (одиночные семена – в купуловидных вместилищах).
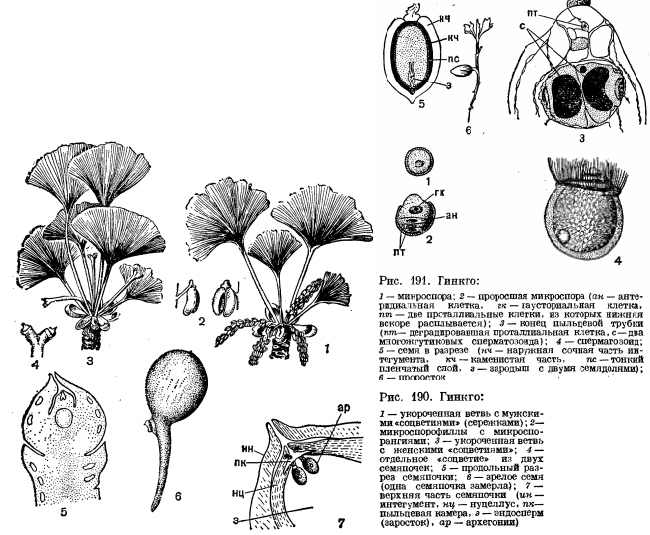
Порядок Ginkgoales. До настоящего времени сохранился один вид – гинкго двулопастной. Появился он около 200 млн лет назад. Это священное дерево высотой 30 м растет во многих парках, окружающих старинные храмы в Китае, Японии и Корее.
-
Пикноксильная линия эволюции; эустела.
-
Двудомное, листопадное растение.
-
Листья с дихотомическим жилкованием расположены на двух типах побегов: брахи– и ауксибластах.
-
Микростробилы сережковидные.
-
На микроспорофиллах образуется по два (до четырех) микроспорангия.
-
Зрелая пыльца 4-клеточная, с двумя проталлиальными клетками. Далее развитие мужского гаметофита гинкго продолжается после попадания пыльцы в пыльцевую камеру семяпочки, где при помощи гаустории гаметофит прикрепляется к стенке пыльцевой камеры. Развитие гаметофита завершается образованием двух сперматозоидов.
-
Макростробилы закладываются в пазухах листьев и несут по две семяпочки, из которых обычно одна образует семя. Оплодотворение у гинкго наступает через несколько месяцев после опыления, а окончательное развитие эмбрио заканчивается уже после опадения семян.